
 商标分类
商标分类  商标转让
商标转让 
上调翻译的反义RNA分子的结构域的制作方法
 2021-02-02 17:02:38|
2021-02-02 17:02:38| 414|
414| 起点商标网
起点商标网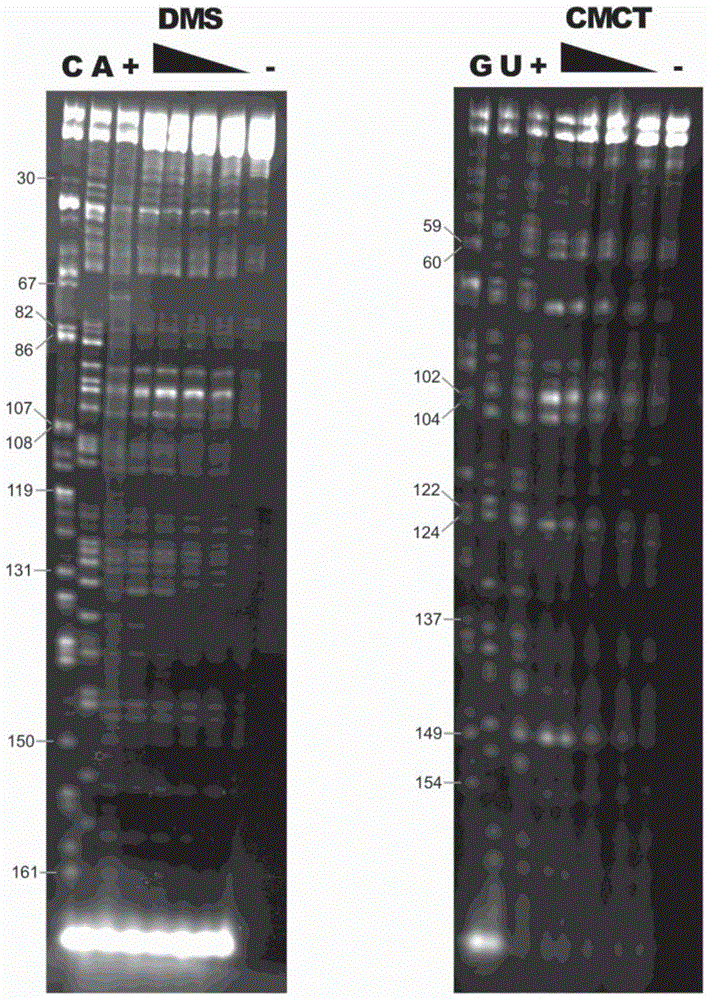
相关申请的交叉引用
本专利申请要求来自2018年2月5日提交的意大利专利申请号102018000002411的优先权,所述申请的公开内容通过引用方式完整并入本文。
本发明涉及功能性核酸分子,其包含靶结合序列和调节序列,所述靶结合序列包含与待增强其蛋白质翻译的靶mrna序列反向互补的序列;并且所述调节序列具有二维结构,所述二维结构包含特定茎-环和内部环结构域和/或特定三维结构。
现有技术
庞大的基因组项目如encode和fantom已经显示,大部分基因组转录,但是小于5%转录的rna编码蛋白质。实际上,非编码性长rna(lncrnas)和转座元件(tes)似乎代表主要的细胞转录输出。在过去十年,数目日益增加的研究正在揭示lncrna功能。总之,日益明显的是,借助众多机制,lncrna在胞核中和细胞质中主要充当调节性分子。尽管取得这种进展,仅小部分注释的lncrnas已经在功能上验证。
最近,鉴定一类新功能性反义(as)lncrna,其增加部分重叠的编码蛋白质的有义mrna翻译。这些rna也称作sineup,因为它们是含有引起蛋白质翻译上调的sine元件的反义rna。
反义(as)uchl1,一种对人uchl1/park5基因的小鼠直向同源物反义的lncrna,可以视为sineup的代表性成员,因为发现它增加在转录后水平发挥作用的uchl1蛋白质合成。asuchl1活性取决于两个功能性结构域的组合:在5'端,作为“结合结构域”显示的重叠区域决定asuchl1针对的特异性uchl1mrna;在3'端,非重叠区域含有嵌合的倒转sineb2元件,所述元件充当“效应子结构域”并触发结合的靶mrna的翻译上调。
超过30种反义lncrnas促进部分重叠性mrna的翻译上调。通过交换结合结构域,可以将asuchl1活性重新指向选择的靶mrna。
将这些天然分析和合成分子命名为sineups,因为它们需要sineb2元件来上调翻译并且在wo2012/133947中公开。可以将asuchl1视为这个新类别lncrnas的代表性成员,其中利用嵌合的转座元件作为实现蛋白质上调的功能性rna结构域。
倒转sineb2元件是充分确定的lncrna结构域的实例。进化研究已经表明,sine元件(如lncrnas)的一级序列通常不充分保守。仅在非嵌合sine中代表rna聚合酶iii内部启动子的a框和b框观察到sine元件内部的部分保守。有趣地,尽管序列同一性差,至少鼠sineb2元件显示功能性保守,因为已经显示众多的sineb2序列在天然和嵌合sineups中充当效应子结构域。另外,鉴定了人sine元件并证明其作为sineup发挥作用(scheina.等人,scirep.(2016),6:33605)。
因此,已经假设嵌合sine的结构域功能保留在基于结构而非基于序列的基序上。
已经假设rna折叠提供为lncrnas和嵌合tes功能性线索。rna序列能够折叠以形成仅已知一级核苷酸序列时难以预测的复杂二级结构和三级结构。系统鉴定lncrna二级结构模块的计算方法仍不令人满意。现有算法能够正确预测约50-70%的已知碱基对。最重要地,二级结构预测模型中的50-70%准确度值因其中该模型正确的区域与明显不一致的区域之间的平均数而产生。因此,测量rna分子在溶液中的实际结构是洞悉rna功能的唯一可用方法。基于化学修饰水可及残基(化学足迹法)和基于核酸磁特征的方法(nmr)可用。最近,已经开发一种新的生物化学方案,称作icshape(体内点击选择性29-羟基酰化和剖析实验),其能够实现活细胞中针对全部四种碱基首次全面查看rna二级结构。
鉴于以上,需要确定特定二维或三维分子结构是否促成lncrna功能,以便在其他研究中和临床环境下能够有效和高效地利用这些分子。尤其,lncrnas代表用于一系列众多研究活动的理想工具,从研究基因功能至工业蛋白质产生和体内应用,如治疗单倍体不足(haploinsufficiencies)。
发明简述
本发明的目的是鉴定来自小鼠asuchl1的倒转sineb2元件的、其他小鼠sines的和来自进化远缘物种的sines的二级和三级结构,所述进化远缘物种的sine在功能性反义lncrna中嵌合,同时与蛋白质编码基因和翻译上调活性有部分重叠。对这些分子的二级和三级结构中的基序及其适宜组合的鉴定将允许设计翻译上调功能增加的sine元件。
此外,对这些基序及其适宜组合的鉴定将允许设计较短的功能性序列。实际上,对于治疗应用,递送sineuprna是一个决定性挑战。rna分子在活生物中高度不稳定。已经广泛使用化学修饰和/或包被于脂质纳米粒子中来克服rna的天然不稳定性。但是,这些递送方法已经被证明对长rna分子低效。设计倒转sineb2效应子结构域的仅保留散在和活性结构性元件的较短变体从“非必需”序列中“扫出”效应子结构域,以获得最短活性sineuprna分子。
通过如权利要求1或2中限定的功能性核酸分子实现这个目的。
本发明的其他目的是提供如权利要求12、15、17、19、22中限定的功能性核酸分子和如权利要求26中限定的组合物。
定义
术语“minisineup”本文中意指由结合结构域(靶mrna的互补序列)、间隔序列(在例举的情况下,21nt间隔序列)和作为效应子结构域的任何sine或sine衍生序列组成的核酸分子(zucchelli等人,frontcellneurosci.,9:174,2015)。
尤其,术语“minisineup-gfp”本文中意指minisineup的结合结构域设计成egfpmrna的互补序列(zucchelli等人,frontcellneurosci.,9:174,2015)。
“功能性核酸分子”本文中一般意指核酸分子能够增强感兴趣的靶mrna翻译。
附图简述
与实施例1有关的图1显示来自天然asuchl1的倒转sineb2序列的化学足迹。
与实施例2有关的图2显示来自天然asuchl1的倒转sineb2序列的二级结构。
与实施例3有关的图3显示倒转sineb2rna的末端缺失片段在溶液中的结构。
与实施例4有关的图4显示依据蛋白质印迹和qrt-pcr,第一茎-环结构域(sl-1)促成asuchl1活性。
与实施例5有关的图5显示依据蛋白质印迹和qrt-pcr,第一茎-环结构域(sl-1)为人肝细胞性细胞系中asuchl1核酸分子翻译上调所需要并伴随uchl1-flag过量表达。
与实施例6有关的图6显示依据蛋白质印迹和qrt-pcr、第一茎-环结构域(sl-1)促成hek293t/17细胞中与gfpmrna(sineup-gfp)重叠的反义lncrna的翻译上调活性。
与实施例7有关的图7显示通过nmr测定倒转sineb2片段(38nt)在溶液中的结构。
与实施例8有关的图8显示invsineb2/38的2dnoesynmr谱(τm=250ms)的芳族异头区域。
与实施例9有关的图9显示用来正确重构倒转sineb238nt片段的3d结构的nmr信号的关联性图。
与实施例10有关的图10显示倒转sineb2效应子结构域的sl-1区域的高分辨率3d结构。
与实施例11有关的图11显示倒转sineb2上的缺失序列和这些缺失对倒转sineb2活性的影响(蛋白质印迹法和qrt-pcr)。
与实施例12有关的图12显示倒转sineb2上突变的序列和这些突变对倒转sineb2活性的影响(蛋白质印迹法和qrt-pcr)。
与实施例13有关的图13显示较短形式的倒转sineb2上突变的序列和这些突变对倒转sineb2活性的影响。
与实施例14有关的图14显示,来自不同小鼠aslncrnas的sineb2元件可以充当在minisineups中的效应子结构域并且在hek293t中上调gfp蛋白表达(蛋白质印迹法)。
与实施例15有关的图15显示来自不同小鼠aslncrnas的sineb2元件的clustalw序列比对。
与实施例16有关的图16显示在具有翻译上调活性的反义功能性核酸分子中充当效应子结构域的来自进化远缘物种的sine序列(蛋白质印迹法和qrt-pcr)。
与实施例17有关的图17显示来自进化远缘物种的sine序列的更强力翻译上调活性与更复杂结构之间的相关性。
与实施例18有关的图18显示来自进化远缘物种的sine序列的2d结构基序分析。
与实施例19有关的图19显示,第一茎-环结构域(sl-1)对于hek293t/17细胞中与gfpmrna重叠的反义lncrna(59-96sl1sineup-gfp)的翻译上调活性足够用。
与实施例20有关的图20显示依据蛋白质印迹,基于结构的较短形式的倒转sineb2足以翻译上调hek293t/17细胞中与gfpmrna(44-120sineup-gfp)重叠的反义lncrna。
与实施例21有关的图21显示,基于结构的较短形式的倒转sineb2足以翻译上调hek293t/17细胞中与内源dj-1mrna(44-120sineup-dj-1)重叠的反义lncrna。还已经测试两种带点突变的变体的功能。
图22显示sineup-icshape实验的示意概览。
图23显示minisineup-gfp效应子结构域在hek293t/17细胞中及其中基序的二级(2d)结构。
图24显示使用exparna,比较(a)长sineup-gfp中和(b)minisineup-gfp中asuchl1sineb2的2d结构。
图25显示(a)长sineup-gfp中和(b)minisineup-gfp中asuxtsineb2-b的2d结构的exparna精确模式匹配结果。
图26显示(a)minisineup-gfp中asuchl1sineb2和(b)长sineup-gfp中astxnipsineb2的2d结构的exparna精确模式匹配结果。
图27显示(a)minisineup-gfp中asuchl1sineb2和(b)长sineup-gfp中asuxtsineb2-b的2d结构的exparna精确模式匹配结果。
图28显示(a)minisineup-gfp中asuchl1sineb2和(b)minisineup-gfp中asuxtsineb2-b的2d结构的exparna精确模式匹配结果。
图29显示(a)minisineup-gfp中asuchl1sineb2和(b)minisineup-gfp中sineb2/b3共有(反向互补物)的2d结构的exparna精确模式匹配结果。
图30显示(a)minisineup-gfp中asuchl1sineb2和(b)minisineup-gfp中asnars2sineb2的2d结构的exparna精确模式匹配结果。
图31显示(a)minisineup-gfp中asuchl1sineb2和(b)minisineup-gfp中asabhd11sineb2的2d结构的exparna精确模式匹配结果。
图32显示(a)minisineup-gfp中asuchl1sineb2和(b)minisineup-gfp中asepb4.9sineb2的2d结构的exparna精确模式匹配结果。
图33显示(a)minisineup-gfp中asuchl1sineb2和(b)minisineup-gfp中aswfdc5sineb2的2d结构的exparna精确模式匹配结果。
图34显示(a)minisineup-gfp中asuchl1sineb2和(b)minisineup-gfp中aspgbd1sineb2的2d结构的exparna精确模式匹配结果。
图35显示(a)minisineup-gfp中asuchl1sineb2和(b)minisineup-gfp中asgsk3βsineb2的2d结构的exparna精确模式匹配结果。
图36显示(a)minisineup-gfp中asuchl1sineb2和(b)minisineup-gfp中asrhodsineb2-mm2的2d结构的exparna精确模式匹配结果。
图37显示(a)minisineup-gfp中asuchl1sineb2和(b)minisineup-gfp中asrhodsineb2-b3a的2d结构的exparna精确模式匹配结果。
图38显示(a)minisineup-gfp中asuchl1sineb2和(b)minisineup-gfp中ase4f1sineb2-mm2的2d结构的exparna精确模式匹配结果。
图39显示(a)minisineup-gfp中asuchl1sineb2和(b)minisineup-gfp中ase4f1sineb2-mm1t的2d结构的exparna精确模式匹配结果。
图40和图41显示as-uchl1sineb2的结构动力学。
图42显示ed长度和结构基序组合各异的合成sineup怎样上调gfp蛋白表达。
图43显示(a)minisineup-gfp中asuchl1sineb2(icshape-导向)和(b)minisineup071ed(预测的rna折叠)的2d结构的exparna精确模式匹配结果。
图44显示(a)minisineup-gfp中asuchl1sineb2(icshape-导向)和(b)minisineup072ed(预测的rna折叠)的2d结构的exparna精确模式匹配结果。
图45显示(a)minisineup-gfp中asuchl1sineb2(icshape-导向)和(b)minisineup073ed(预测的rna折叠)的2d结构的exparna精确模式匹配结果。
图46显示(a)minisineup-gfp中asuchl1sineb2(icshape-导向)和(b)minisineup074ed(预测的rna折叠)的2d结构的exparna精确模式匹配结果。
图47显示(a)minisineup-gfp中asuchl1sineb2(icshape-导向)和(b)minisineup075ed(预测的rna折叠)的2d结构的exparna精确模式匹配结果。
图48显示来自asuchl1的sineb2的结构片段怎样上调gfp蛋白表达。
与实施例48有关的图49显示作为环状rna表达时,sineup-dj-1有活性。
发明详述
在本发明中,公开了来自asuchl1的倒转sineb2转座元件的二级和三级结构作为sineup的代表性效应子结构域,这为其中之一。
缺失突变体用来显示sineup活性所需的结构性元件。
另外,在活细胞中测定来自具有功能翻译上调的反义lncrnas(天然sineups)的结构sineb2序列。
还基于sineup活性的生物化学表征和活性和无活性sine的icshape特征(后者未显示)限定结构-功能关系。
公开了sineb2序列的基于结构的活性基序。
还公开了来自进化远缘物种的sine序列的功能性效应子结构域的翻译上调活性,所述sine序列的一级序列不保守并且其二级结构元件已知。
特别地,功能性实验显示,短末端发夹环结构,也称作第一茎-环结构域(sl-1),是sineups上调部分重叠性mrna翻译的能力的关键结构性决定因素,并且额外的结构性元件还对增强活性重要并且区分那些更有活性的sine与活性更小的那些。
证明rna分子的一级序列不必然地导致预定的二级结构以及取决于条件,分子的备选折叠可能发生。换句话说,从rna分子的一级序列获得的二级结构绝对不是预定的且不是仅从一级序列可预测。
此外,提供了sine序列的效应子结构域的几种变体,其中突变所述变体以强化二级结构和/或限制分子的大小。已经通过以下方式设计这些变体:鉴定二级结构中特别相关和保守的基序并且组合它们,从而以实现功能所要求的最小结构并且按最佳序列和条件以获得这种结构。
实际上,本发明的功能性核酸分子包含靶结合序列,所述靶结合序列包含与待增强其蛋白质翻译的靶mrna序列反向互补的序列;并且具有二维结构的调节序列,所述二维结构包含第一茎-环结构域(sl-1),所述第一茎-环结构域包含8至12个配对核苷酸的茎和包含5至9个核苷酸的环。该调节序列具有蛋白质翻译增强效率。
根据本发明,它已经显示,只要二级结构保守,则一级序列的碱基可以在很大程度上变动。
靶结合序列需要与反向互补于靶mrna的序列具有仅约60%相似性。作为事实,靶结合序列可以甚至显示众多错配并保留活性。实际上,mirnas可以影响其甚至具有众多错配的有义mrna配偶物。这个构思已经显示于wo2012/133947中。
靶结合序列和调节序列优选地由间隔序列/接头序列连接。
优选地,第一茎-环结构域(sl-1)的茎富含g/c,因而“富含g/c序列”本文中意指具有超过50%g和c和优选地超过75%g和c的序列。
优选地,第一茎-环结构域(sl1)的环富含a/u,因而“富含a/u序列”本文中意指具有超过50%a和u和优选地超过75%a和u的序列。
第一茎-环结构域(sl-1)的茎优选地具有10个配对核苷酸并且第一茎-环结构域(sl-1)的环优选地具有7个核苷酸。
本发明的功能性核酸分子包含靶结合序列和调节序列,所述靶结合序列包含与待增强其蛋白质翻译的靶mrna序列反向互补的序列;并且所述调节序列具有二维结构列,所述二维结构包含第一茎-环结构域(sl-1),所述第一茎-环结构域具有与seqidno:1有至少50%同一性、优选地与seqidno:1有至少75%同一性、更优选地与seqidno:1有至少90%同一性的序列。
在例举的来自asuchl1的倒转sineb2转座元件中,第一茎-环结构域(sl-1)包含seqidno:1,即seqidno:5(其是来自asuchl1的倒转sineb2转座元件的完整序列)的核苷酸64至92。
优选地,第一茎-环结构域(sl-1)的茎具有a型螺旋三维结构,如通过高分辨率nmr所示。
调节序列的二维结构优选地还包含第二茎-环结构域(sl-2)和第四内部环结构域(il-4)或第四茎-环结构域(sl-4),第二茎-环结构域(sl-2)包含具有2至4个配对核苷酸的茎和包含6至10个核苷酸的环,并且第四内部环结构域(il-4)包含14至24个核苷酸,并且第四茎-环结构域(sl-4)包含具有3至5个配对核苷酸的茎和包含6至10个核苷酸的环。
第二茎-环结构域(sl-2)的茎优选地富含g/c。第二茎-环结构域(sl-2)和/或第四内部环结构域的环是/优选地富含a/u的。
第二茎-环结构域(sl-2)的茎优选地具有3个配对核苷酸并且第二茎-环结构域(sl-2)的环优选地具有8个核苷酸。第四内部环结构域(il-4)优选地包含16至22个核苷酸。
本发明的功能性核酸分子优选地包含具有二维结构的靶结合序列,所述靶结合序列包含与seqidno:2有至少50%同一性、优选地与seqidno:2有至少75%同一性、更优选地与seqidno:2有至少90%同一性的序列。
在例举的来自asuchl1的倒转sineb2转座元件中,调节序列包含seqidno:2,即seqidno:5的核苷酸52至112。
调节序列的二维结构优选地还包含第三内部环结构域(il-3),其包含6至14个核苷酸、优选地7至13个核苷酸。
调节序列的二维结构优选地还包含第三茎-环结构域(sl-3),其包含具有1至3个配对核苷酸的茎和包含3至5个核苷酸的环。
本发明的功能性核酸分子优选地包含具有二维结构的调节序列,所述调节序列包含与seqidno:3有至少50%同一性、优选地与seqidno:3有至少75%同一性、更优选地与seqidno:3有至少90%同一性的序列。
在例举的来自asuchl1的倒转sineb2转座元件中,调节序列包含seqidno:3,即seqidno:5的核苷酸36至133。
调节序列的二维结构优选地还包含第二内部环结构域(il-2),其包含6至12个核苷酸、优选地7至11个核苷酸。第二内部环结构域(il-2)优选地是不对称的和/或富含a/u。
本发明的功能性核酸分子优选地包含具有二维结构的调节序列,所述调节序列包含与seqidno:4有至少50%同一性、优选地与seqidno:4有至少75%同一性、更优选地与seqidno:4有至少90%同一性的序列。
在例举的来自asuchl1的倒转sineb2转座元件中,调节序列包含seqidno:4,即seqidno:5的核苷酸22至150。
调节序列的二维结构优选地还包含第一内部环结构域(il-1),其包含6至12个核苷酸、优选地7至11个核苷酸。
本发明的功能性核酸分子优选地包含具有二维结构的调节序列,所述调节序列包含与seqidno:5有至少50%同一性、优选地与seqidno:5有至少75%同一性、更优选地与seqidno:5有至少90%同一性的序列。
在例举的来自asuchl1的倒转sineb2转座元件中,调节序列具有seqidno:5。
本发明的另一个功能性核酸分子包含靶结合序列,所述靶结合序列包含与待增强其蛋白质翻译的靶mrna序列反向互补的序列;和调节序列,所述调节序列包含与选自seqidno:15、seqidno:16、seqidno:17、seqidno:18、seqidno:19、seqidno:20、seqidno:21、seqidno:15、seqidno:7、seqidno:22和seqidno:23的序列具有至少50%、优选地至少75%、甚至更优选地至少90%同一性的序列。
icshape分析显示,上文公开的茎中配对核苷酸的数目和上文公开的环中核苷酸的数目可以超出公开的范围变动。
本发明的另一个功能性核酸分子包含靶结合序列,所述靶结合序列包含与待增强其蛋白质翻译的靶mrna序列反向互补的序列;和调节序列,所述调节序列包含与选自seqidno:7、seqidno:21、seqidno:17、seqidno:22、seqidno:65、seqidno:66和seqidno:67的序列具有至少50%、优选地至少75%、甚至更优选地至少90%同一性的序列。
调节序列优选地包含seqidno:7、seqidno:21、seqidno:17、seqidno:22、seqidno:65、seqidno:66和seqidno:67。
在一个备选实施方案中,功能性核酸包含调节序列,所述调节序列包含与seqidno:12具有至少50%、优选地至少75%、甚至更优选地至少90%同一性的序列,甚至更优选地该序列是seqidno:12。
本发明的另一个功能性核酸分子包含靶结合序列,所述靶结合序列包含与待增强其蛋白质翻译的靶mrna序列反向互补的序列;和调节序列,所述调节序列包含以下一个或多个基序:由与gcu(基序1)、agggc(基序3)、uggau(基序4)、uggugg(基序6)或ugga(基序7)具有50%、优选地75%、甚至更优选地90%同一性的rna序列形成并由其杂交的反向互补rna形成的基序,或由与aggagg(基序2)具有50%、优选地75%、甚至更优选地90%同一性的rna序列形成并由其部分杂交的互补性反向rna形成的基序、或由与ugg(基序5)具有50%、优选地75%、甚至更优选地90%同一性的rna序列形成并由其部分杂交的互补性反向rna形成的基序。
甚至更优选地,这些基序分别由gcu(基序1)、agggc(基序3)、uggau(基序4)、uggugg(基序6)和ugga(基序7)并由其杂交的反向互补性rna形成、由aggagg(基序2)并由其部分杂交的互补性反向rna形成、以及由ugg(基序5)并由其部分杂交的互补性反向rna形成。
上述功能性核酸分子的调节序列优选地包含由与uggugg(基序6)具有90%同一性的rna序列形成并由其杂交的反向互补性rna形成的基序。更优选地,基序是uggugg。
图24至图39、图43至图47和图48b中显示上述基序的优选组合。
更具体地,上述功能性核酸分子包含与选自seqidno:69、seqidno:70、seqidno:71、seqidno:72、seqidno:73、seqidno:74、seqidno:75、seqidno:76和seqidno:77的序列具有至少50%、优选地75%、甚至更优选地90%同一性的序列作为调节序列。甚至更优选地,该序列包含seqidno:69、seqidno:70、seqidno:71、seqidno:72、seqidno:73、seqidno:74、seqidno:75、seqidno:76和seqidno:77。在这些序列当中,优选的序列是seqidno:72,其代表最小功能性序列的实例,其中即便缺少全长sineup效应子结构域的几个基序,所述最小功能性序列维持增加靶rna翻译的能力。
本发明的功能性核酸分子优选地是环状分子。这种构象导致在细胞内部更难降解并且因此保持更长时间有活性的稳定得多的分子。
本发明还涉及包含上述功能性核酸分子的组合物。包括允许通过病毒载体(aav、慢病毒等)和非病毒载体(纳米粒子,脂质粒子等)递送上述功能性核酸分子的任何组合物。
实施例
实施例1
这个实例展示来自天然asuchl1的倒转sineb2序列中每个核苷酸的化学物质(dms和cmct)可及性。化学物质可及性严格依赖于分子的局部结构。
图1显示invsineb2/183rna构建体的化学足迹。左小图显示带c和a测序泳道连同核苷酸编号的荧光凝胶。具有经渐增dms浓度处理的invsineb2/183rna连同用阳性(+)和阴性(-)对照的dms泳道。右小图显示有核苷酸编号的带g和u测序泳道的荧光凝胶。具有经渐增cmct浓度处理的invsineb2/183rna连同用阳性(+)和阴性(-)对照的cmct泳道。实验如tijerinap.等人,(2007),natureprotocols2:2608-2623中所述实施。
实施例2
这个实施例显示功能性asuchl1的倒转sineb2效应子结构域的二维结构。该结构在茎-环(sl1、sl2和sl3)、内部环(il1、il2、il3和il4)和间插性茎结构中组织。该结构基于来自实施例1的数据生成。
图2显示asuchl1的倒转sineb2效应子结构域的二级结构。dms反应性核苷酸在带灰色背景的圆圈内以黑色字符显示,并且cmct反应性核苷酸在带黑色背景的圆圈内以白色字符。内部环和茎-环分别标记为ilx和slx仅圈选非反应性核苷酸。以灰色加阴影的区段对应于dna引物杂交位点。
实施例3
这个实施例表明,倒转sineb2序列的较短片段保留末端sl1结构。
图3显示倒转sineb2rna的末端缺失片段在溶液中的结构。图3a显示asuchl1的invsineb2/183效应子结构域的2d结构。正方形中指示末端移除的区域。图3b显示末端缺失片段的示意性结构。核苷酸位置相对于全长序列指示。图3c显示来自小鼠asuchl1的倒转sineb2的亚胺基质子区域。亚胺基1h信号归属于每个末端缺失的片段。显示片段#1(seqidno:9)、#2(seqidno:8)、#3(seqidno:7)和#4(seqidno:6)迹线。全部片段中均观察到sl1茎结构。
实施例4
这个实施例表明,sl1为功能性核酸分子(asuchl1)中倒转sineb2效应子结构域的翻译上调活性所需要。
将鼠神经母细胞瘤n2a细胞(atcc目录号atcc-ccl-131)用asuchl1和δsl1突变体构建体转染。在δsl1突变体构建体中,嵌合倒转sineb2的核苷酸68-77(seqidno:10)缺失。在δsl1突变体中,摧毁sl-1的茎环结构,但是保留整个倒转sineb2组合体。用空对照质粒转染对照细胞。转染后48小时,将细胞裂解并加工以获得蛋白质水平和rna水平。用抗uchl1抗体进行蛋白质印迹(图4a)。使用β-肌动蛋白作为上样对照。在蛋白质印迹图像上对β-肌动蛋白标准化并相对于空白对照样品,计算诱导倍数。使用特异性引物,通过qrt-pcr监测uchl1mrna(灰色标度)和asuchl1(白色标度)的表达。数据表明均数±st.dev。数据代表n=5独立重复。图4b显示asuchl1和δsl1对n2a细胞中内源uchl1mrna的翻译增强活性的图示(n=5)。*,p=0.01;ns,不显著(p>0.5)。
实验如zucchellis.等人,(2015),frontcellneurosci.9:174中所述实施。
实施例5
这个实施例表明,sl-1为人肝细胞性细胞系中asuchl1核酸分子翻译上调功能连同过量表达的mrna所需要。实际上,在人hepg2细胞(atcc目录号atcc-hb-8065)中用过量表达的uchl1-flag测试时,sl-1的缺失取消了asuchl1活性。
将人肝细胞hepg2细胞系用联合asuchl1和δsl1突变体构建体的uchl1-flag构建体转染。用空对照质粒转染对照细胞。转染后48小时,将细胞裂解并加工以通过蛋白质印迹获得蛋白质定量。用抗flag抗体进行蛋白质印迹以监测过量表达的蛋白质(图5a)。使用β-肌动蛋白作为上样对照。在蛋白质印迹图像上对β-肌动蛋白标准化并相对于空白对照样品,计算诱导倍数。asuchl1翻译增强子活性以粗体突出显示。将如上文公开那样处理的细胞加工以定量rna(图5b)。使用特异性引物,通过qrt-pcr监测uchl1mrna(右)和asuchl1(左)的表达。数据表明均数±st.dev。
实施例6
这个实施例表明,sl-1为靶向过量表达的mrna的功能性合成核酸分子(sineup-gfp)的翻译上调功能所需要。优选地还需要额外的结构性元件,因为stemdm突变体(其使茎去稳定化)已经削弱,但是未取消效应子结构域活性。
图6显示sl-1结构性元件在hek293t/17细胞(atcc目录号atcc-crl-11268)中促成sineup-gfp活性。图6a是来自倒转sineb2效应子结构域的末端发夹结构(sl-1)的放大图像。通过在sineup-gfp的sineb2效应子结构域中缺失,产生δsl1突变体。类似地,在sineup-gfp中引入位点定向突变(g70a和g73a,划圈)以产生stemdm突变体(seqidno:11)。在图6b中,如所示,将hek293t/17细胞用联合空载体(-)的egfp表达质粒,或sineup-gfp全长(fl)、δsl1或stemdm突变体转染。用抗gfp抗体进行蛋白质印迹,以监测过量表达的蛋白质。使用β-肌动蛋白作为上样对照。在蛋白质印迹图像上对β-肌动蛋白标准化并相对于空白对照样品,计算诱导倍数。sineup-gfp翻译增强子活性以红色突出显示。在图6c中,使用如图6b中那样处理的细胞来制备rna。使用特异性引物,通过qrt-pcr监测gfpmrna(灰色)和sineup-gfp(黑色)的表达。数据表明均数±st.dev。
实施例7
这个实施例表明,涵盖sl1结构区(seqidno:12)的倒转sineb2片段(38个核苷酸)在溶液中采取单一结构,如通过nmr所测定。在这个实验中,nmr数据验证了sl-1在较短片段中的结构(如实施例1、2和3中通过其他技术所观测)。
图7a显示在0和25℃获得的5%2h2o/95%h2o中38ntrna的1hnmr谱的亚胺基区域和芳族区域。归属亚胺基1h信号。图7b显示15nhsqcnmr谱的亚胺基区域。在25℃和0℃观察到的信号分别处于深灰色和浅灰色。图7c显示sineb2/38构建体的二级结构。碱基对连同在25℃和0℃可观察的亚胺基信号分别以深灰色和浅灰色加阴影。实验如sybrens.等人,(1998),prog.nucl.magn.reson.spectrosc.32:287-387中所述实施。
实施例8
这个实施例表明,可以在2dnoesynmr谱中分配nmr信号以正确重构倒转sineb238nt片段的3d结构。
图8显示相关共振的分配。invsineb2/38的2dnoesynmr谱(τm=250ms)的芳族异头区域。顺序行走绘为黑线。
实施例9
这个实施例展示了用来正确重构倒转sineb238nt片段的3d结构的nmr信号的关联性图。
图9显示noe关联性的简图。invsineb2/38分子的强烈(粗线)、中等(实线)和微弱(短划线)noe关联性的简图。
实施例10
这个实施例展示倒转sineb2效应子结构域的sl-1区域的高分辨率3d结构。发夹采取a型螺旋茎。环显示溶液中的动态特性。
图10显示sl1的高分辨率结构。图10a显示29nt发夹(seqidno:1)的10个最低能量结构的侧视图并且图10b显示其俯视图。图10c和图10d是具有g77、u78和g79碱基取向的环的代表性结构。
实施例11
这个实施例表明,附加结构性元件(il3-sl3区域、il4和基部茎结构)促成功能性核酸分子(尤其,sineup-gfp)中的倒转sineb2效应子结构域活性。
图11a是在倒转sineb2二级结构上注释的缺失序列δsl1(seqidno:10)、δpolii(seqidno:13)和δbox(seqidno:14)的示意图。在图11b中,如所示,将hek293t/17细胞用联合空载体(-)的egfp表达质粒,或sineup-gfp全长(fl)、δsl1、δpolii或δbox突变体转染。用抗gfp抗体进行蛋白质印迹,以监测过量表达的蛋白质。使用β-肌动蛋白作为上样对照。在蛋白质印迹图像上对β-肌动蛋白标准化并相对于空白对照样品,计算诱导倍数。sineup-gfp翻译增强子活性以粗体突出显示。结构突变体的贡献以红色显示。在图11c中,使用如图11b中那样处理的细胞来制备rna。使用特异性引物,通过qrt-pcr监测gfpmrna(灰色)和sineup-gfp(黑色)的表达。数据表明均数±st.dev。
实施例12
这个实施例表明,合成的新效应子结构域序列可以基于结构性特征(在il-4、sl-2、sl-1基部的间插性茎结构)设计。这些序列可以在mrna过量表达的功能性核酸分子中增强翻译上调活性。在一级核苷酸序列不同,但结构相同情况下获得增强的活性。
特别地,倒转sineb2的基于结构的变体显示如sineup-gfp中效应子结构域那样的增加的翻译上调活性。图12a显示在倒转sineb2二级结构上注释的基于结构的突变的示意图。在圆圈中:单点突变;在正方形中:结构性元件的序列交换(茎和环-碱基)。在图12b中,将hek293t/17细胞用联合空载体(-)的egfp表达质粒或作为阳性对照的sineup-gfp全长(fl)转染。如所示那样转染带有基于结构的突变的sineup-gfp构建体。用抗gfp抗体进行蛋白质印迹,以监测过量表达的蛋白质。使用β-肌动蛋白作为上样对照。图12c显示已经如b中进行的n=9实验中gfp蛋白质水平相对升高(左小图)和gfpmrna水平未改变(右小图)。每种sineup-gfp结构突变体(asgfp)的表达通过qrt-pcr监测并且纳入右小图。全部突变体均按相同水平表达。
实施例13
这个实施例表明,合成的新效应子结构域序列可以基于结构性特征(在il-4、sl-2、sl-1基部的间插性茎结构)设计并且在mrna过量表达的功能性核酸分子中获得增强的翻译上调活性。在一级核苷酸序列不同,但结构相同情况下获得增强的活性。
特别地,倒转sineb2的较短和基于结构的变体显示如靶向内源mrna的功能性核酸分子的效应子结构域那样的增加的翻译上调活性。
图13a是在倒转sineb2二级结构上注释的基于结构的突变的示意图(27-142对应于seqidno:21,44-120对应于seqidno:7,带a45g的44-120对应于seqidno:22和带a51c的44-120对应于seqidno:23)。显示单点突变。图13b显示靶向内源表达的mrna的功能性核酸分子的翻译上调活性,其中效应子结构域是具有改进结构性特征的合成性序列。取决于条件,备选性折叠出现并且可以变得更强,如seqidno:7。
实施例14
这个实施例表明,来自天然反义非编码性长rna的sine序列是功能性核酸分子中用于翻译上调部分重叠性mrna的效应子结构域。
特别地,来自不同小鼠aslncrnas的sineb2元件可以充当在minisineups中的效应子结构域并且在hek293t中上调gfp蛋白表达(rikenbrc目录号rcb2202)。
图14a显示来自天然功能性反义lncrnas的sineb2序列,其中所述序列被克隆入minisineup-gfp以代替来自asuchl1的原始的倒转sineb2序列作为效应子结构域。相对于启动性aug密码子,gfp靶向序列是-40/+4个核苷酸。序列按顺序是:seqidno:44、seqidno:47、seqidno:50至seqidno:62。图14b显示在有义egfp和minisineup-gfp共转染后(转染后24小时),hek293t细胞中的gfp蛋白质诱导倍数。使用imagej软件计算蛋白质印迹图像和相应的gfp条带强度(对actb表达水平标准化)。使用空载体作为阴性对照,而使用asuchl1(含有minisineup的asuchl1sineb2)作为阳性对照。样品标签显示从中分离sineb2序列的aslncrnas的名称。测试了代表四个亚家族(基于repeatmasker注释)(即,b3、b3a、mm2和mm1t)的sineb2序列。除非特别指出,否则sineb2元件来自b3亚家族。asuchl1+asuxt-b意指在ed中组合的asuxt的asuchl1sineb2和第二sineb2。3x表示sineb2的三个重复,并且sineb2/b3共有序列是取自repbase数据库的b3亚家族共有序列。n=5;误差线±sem;****p<0.00005;***p<0.0005;**p<0.005;*p<0.05;双尾studentt检验
图14c显示作为sineup-gfp中ed受测的sineb2序列。
图14d显示了使用imagej软件计算的采用抗gfp和抗actb抗体的蛋白质印迹图像和相应的gfp条带强度(对actb表达水平标准化)。使用actb作为上样对照。这里,asuchl1sineb2代表相对于egfp起始密码子具有-40/+32bd设计(+21nt间隔序列)和除alu和来自asuchl1的3′尾之外以asuchl1sineb2作为ed的sineup-gfp。astxnipsineb2和asuxt-sineb2b遵循相同的设计,例外是asuchl1sineb2由分别来自astxnip和asuxt(sineb2-b)的sineb2元件替换。asuchl1_δ5′-28nt与asuchl1sineb2相同,但是bd较短(相对于egfp起始密码子-40/+4)。direct=sineb2的正方向;δ=缺失。n=3;误差线±sem;**p<0.005;*p<0.05;双尾studentt检验
实施例15
这个实施例表明,功能性核酸分子中用于翻译上调功能的活性sine序列具有差的序列同一性。这个实施例显示效应子结构域功能的结构依赖性。
图15显示来自不同小鼠aslncrnas的sineb2元件的clustalw序列比对。在mega5软件中借助clustalw比对为sineup-gfp和minisineup-gfp中翻译上调功能所选定的小鼠sineb2序列(实施例14)。序列按顺序是:seqidno:44至seqidno:49和seqidno:52至seqidno:62。比对结果中的空位用“-”指示。碱基由四个不同灰阶突出显示。a框和b框由黑色方块代表。比对结果正在显示在sine的相同家族成员内部的序列变异。
实施例16
这个实施例表明,来自进化远缘物种的sine序列在具有翻译上调活性的反义功能性核酸分子中充当效应子结构域。
如图16a中所示,选择来自进化远缘物种并具有限定二级结构的sine序列(sunfen-jie等人,trendsingenetics,2011)用于分析。依据比对分析,这些序列显示差的序列同一性。序列表包括与16a图中所示dna序列相对应的rna序列(seqidno:24至seqidno:33)。序列表还包括倒转rna序列(seqidno:34至seqidno:43)。在图16b中,如所示,将hek293t/17细胞用联合空载体(-)的egfp表达质粒,或按正方向和倒转方向克隆的具有原始倒转sine序列(深蓝色,b2)或具有所示效应子结构域交换突变体的sineup-gfp转染。使用celigos自动化成像系统,记录gfp自身荧光。数据显示对空载体(pcdna3.1)转染的细胞(n>7次重复)标准化的gfp荧光的结果。数据表明均数±stdev。加框架的列显示其中如与sineb2阳性对照相比,sineup活性增强的情况。如图16c中所示,qrt-pcr分析显示,蛋白质上调伴随稳定的gfpmrna水平(转录后)出现。在celigos测量后,提取和纯化rna。采用特异性引物qrt-pcr分析gfpmrna和sineup-gfprnas。n=3,sineb2设定为1。
实施例17
这个实施例表明,来自进化远缘物种的sine序列的更强力翻译上调活性与更复杂的二级结构(茎环组合)相关。
在图17a中,将hek293t/17细胞用联合空载体(-)的egfp表达质粒,或具有原始倒转sine序列(sineb2)或具有所示效应子结构域交换突变体的sineup-gfp转染。用抗gfp抗体进行蛋白质印迹分析,以监测过量表达的蛋白质。使用β-肌动蛋白作为上样对照。在图17b中,针对β-肌动蛋白对照标准化时,通过图像分析计算gfp蛋白质量。来自n=5次重复的数据表述为均数±stdev。使用t检验,进行统计分析(显示p值)。图17c显示最有活性的来自各种物种的sine,连同其相对结构。总体上,sine结构用线(茎)和圆圈(发夹环)表述。应当指出,全部最有活性的sine均共有一个保守的顶端茎环结构。实际上,显示这个茎-环结构是进化远缘物种的sine中翻译上调活性的关键元件。
实施例18
这个实施例表明,功能性核酸分子的更强力的sine序列之间共有结构性基序(基序1、il-3/sl-3和基序2、sl-1)。
图18显示功能性核酸分子中具有翻译上调活性的来自进化远缘物种的sine序列的2d结构基序分析。图18a显示来自实施例2的倒转sineb2rna(seqidno:5)的2d结构。图18b显示b2共有sinerna(seqidno:30)的预测2d结构,所述结构由rna折叠webserver以默认设置确定。图18c显示使用酶促探测法,smaisinerna(seqidno:27)的2d结构。linert的推定性识别区由短划线加框(kawagoe-takaki等人gene365(2006),67-73)。图18d显示来自倒转sineb2和b2sine的2d结构的茎结构基序1(中间茎、il-3、sl-3)。基序1结构堵在中间环结构之间。图18e显示来自倒转sineb2、b2sine和smaisine的2d结构的茎结构基序2(sl-1)。基序2结构堵于边缘环结构附近。
再次应指出,图18a、图18b和图18c中的三个结构共有一个保守的顶端茎环结构。
实施例19
这个实施例表明,合成的新效应子结构域序列可以基于结构性特征(38nt、茎环结构sl1)设计并且这些序列在mrna过量表达的功能性核酸分子中拥有翻译上调活性。在一级核苷酸序列不同,但结构相同情况下获得活性。特别地,倒转sineb2的较短和基于结构的变体显示如靶向内源mrna的功能性核酸分子(59-96sl1sineup-gfp)的效应子结构域那样的翻译上调活性。图19a是在突出显示sl1(59-96nt对应于seqidno:12)的倒转sineb2二级结构上注释的基于结构的突变的示意图。图19b显示靶向内源表达的gfpmrna的功能性核酸分子的翻译上调活性,其中效应子结构域是具有保守结构性特征的较短合成性序列(38ntsl1)。如所示,将hek293t/17细胞用联合空载体(-)的egfp表达质粒,或1-183sineup-gfp(包含seqidno:5)、1-183sineup-gfp聚腺苷酸(seqidno:63)和sl159-96聚腺苷酸sineup-gfp(seqidno:64)转染。用抗gfp抗体进行蛋白质印迹,以监测过量表达的蛋白质。使用β-肌动蛋白作为上样对照。在蛋白质印迹图像上对β-肌动蛋白标准化并相对于空白对照样品,计算诱导倍数。图19c中显示来自4个独立实验的数据。sl1sineup-gfp翻译增强子活性以红色指示。数据表明均数±st.dev。
这个实验显示,第一茎-环结构域(sl-1)对于hek293t/17细胞中与gfpmrna重叠的反义lncrna(59-96sl1sineup-gfp)的翻译上调活性足够用。
实施例20
这个实施例表明,合成的新效应子结构域序列可以基于结构性特征(44-120ntsineup,含有il4、sl1和sl2并对应于seqidno:7)设计并且它们在mrna过量表达的功能性核酸分子中拥有翻译上调活性。在一级核苷酸序列不同,但结构相同情况下获得活性。特别地,倒转sineb2的较短和基于结构的变体显示如靶向内源mrna的功能性核酸分子的效应子结构域那样的翻译上调活性。另外,在具有截短型倒转sineb2元件44-120的sineup内部的突变u78g维持了相似的翻译上调活性,而三重突变gug77-79ccc明显地削弱活性,从而证明核苷酸77-79的功能重要性。图20a是在突出显示野生型序列和突变序列(具有u78g的44-120对应于seqidno:65;具有gug77-79ccc的44-120对应于seqidno:66)的截短型倒转sineb2二级结构上注释的基于结构的突变的示意图。图20b显示靶向内源表达的gfpmrna的功能性核酸分子的翻译上调活性,其中效应子结构域是在77-79核苷酸位置具有wt序列或突变型序列的较短合成性序列。将hek293t/17细胞用联合空载体(-)的egfp表达质粒或44-120sineup-gfp、具有u78g的44-120sineup-gfp和具有gug77-79ccc的44-120sineup-gfp转染。用抗gfp抗体进行蛋白质印迹,以监测过量表达的蛋白质。使用β-肌动蛋白作为上样对照。在蛋白质印迹图像上对β-肌动蛋白标准化并相对于空白对照样品,计算诱导倍数。44-120sineup-gfp活性既类似于整个倒转sineb2序列的sineup-gfp活性,又类似于具有截短型44-120效应子结构域的带u78g突变的44-120sineup-gfp。通过比较,具有gug77-79ccc的44-120sineup-gfp展示降低的活性。
这个实验显示,基于结构的较短形式的倒转sineb2足以翻译上调hek293t/17细胞中与gfpmrna(44-120sineup-gfp)重叠的反义lncrna。它还显示,倒转sineb2序列的核苷酸77-79对上调翻译能力重要。全部三个构建体均有功能并且二级结构以最佳方式呈递77-79环的三个核苷酸。即便这些三个核苷酸经过修饰,分子仍保留功能。
实施例21
这个实施例表明,合成的新效应子结构域序列可以基于结构性特征(44-120ntsineup,含有il4、sl1和sl2,对应于seqidno:7)设计并且它们在mrna内源表达的功能性核酸分子中拥有翻译上调活性。在一级核苷酸序列不同,但结构相同情况下获得活性。特别地,倒转sineb2的较短和基于结构的变体显示如靶向内源mrna的功能性核酸分子的效应子结构域那样的翻译上调活性。另外,在截短型倒转sineb2二级结构上注释的基于结构的突变可以增加翻译激活物能力(44-120sineup-dj-1变体,其在截短型倒转sineb2序列中具有以下6个点突变:a45g;u48c;u66g;u88c;u90c;u115c并称作44-120sineup-dj-1-强,对应于seqidno:67),而其他在截短型倒转sineb2二级结构上注释的基于结构的突变可以保持翻译上调能力不变或较低(44-120sineup-dj-1变体,其在截短型倒转sineb2序列中具有以下5个点突变:g49a;u66a;g68a;g116a;c119a并称作44-120sineup-dj-1-弱,对应于seqidno:68)。设计这两种变体背后的依据是,根据44-120sineup的结构,在两段dsrna中(44-50连同114-120和64-74连同82-92)存在5个碱基对错配(45-119;48-116;49-115;66-90;68-88)。在44-120sineup强中,已经设计突变体碱基对以建立g:c对。在44-120sineup弱中,已经设计突变体碱基对以建立u:a对。由于g:c对因相对于二个氢键,形成三个氢键(连同其他因素)而更稳定,故已经将具有5个g:c对的突变体44-120sineup指示为强,同时已经将具有5个u:a对的突变体44-120sineup指示为弱。
图21a是在截短型倒转sineb2二级结构上对44-120sineup-dj-1-强和44-120sineup-dj-1-弱注释的基于结构的突变的示意图。突变位置由箭头显示。图21b显示靶向内源表达的dj-1mrna的功能性核酸分子的翻译上调活性,其中效应子结构域是较短的合成性序列(44-120sineup-dj-1)或用6个点突变修饰(44-120sineup-dj-1-强)或用5个点突变修饰(44-120sineup-dj-1-弱)的相同序列。将hek293t/17细胞用联合空载体(-)的egfp表达质粒或44-120sineup-dj-1、44-120sineup-dj-1-强和44-120sineup-dj-1-弱转染。用抗dj-1抗体进行蛋白质印迹,以监测过量表达的蛋白质。使用β-肌动蛋白作为上样对照。在蛋白质印迹图像上对β-肌动蛋白标准化并相对于空白对照样品,计算诱导倍数。图21c中显示来自3个独立实验的数据。44-120sineup-dj-1活性类似于44-120sineup-dj-1-弱,而44-120sineup-dj-1-强以统计显著方式较高。数据表明均数±st.dev。
这个实验显示,基于结构的较短形式的倒转sineb2足以进行翻译上调(44-120sineup-dj-1)。另外,44-120sineup-dj-1-强相对于44-120sineup-dj-1呈现更高翻译激活物活性。44-120sineup-dj-1-弱呈现类似于或低于44-120sineup-dj-1的翻译上调活性。
实施例22
图23a显示minisineup-gfp中asuchl1sineb2的icshape数据指导的二级结构。使用icshape富集评分数据作为viennarna的rnafold工具中的软约束。线性映射方法用来导出配对概率并且选择zarringhalam等人(2012)描述的方法以将导引性拟能并入折叠算法。为了绘制二级结构,使用服务器forna。深色点,icshape富集评分≥0.8;浅色点,富集评分0.5-0.8。富集评分依据0至1评分评定,其中1表示核苷酸处于非配对状态的概率高并且0表示配对倾向性的概率高。sineb2的b框区域和a框区域用线条标记(分别在该图的上部分中和该图的下部分)。使用计算两个rna共同的精确匹配序列-结构基序的exparna工具(heyne,s.等人bioinformatics,2009,25:2095-2102),将这项研究中功能测试过的全部sineb2s的序列和icshape指导的2d结构与asuchl1sineb2的这个结构和序列配对比较。因此,随后筛选从全部配对比较衍生的基序文件,以抽取asuchl1sineb2的位置,所述位置存在于全部比较中长度至少2mer的一个或多个基序中。全部sineb2s之间共有的相似序列和结构基序的位置以带有其相应基序编号的正方形指示。由于考虑2mer和较长基序以鉴定共同区域并且sineb2序列高度可变,故大部分情况下,仅可以找到部分匹配。图23b显示图23a中所示共同序列和结构基序的放大视图。应当指出,基序2和3、和基序4、5、6和7是相似的并且分别主要地由agg和ugg三核苷酸组成。不同的基序编号基于其序列和结构毗邻情况和sineb2上的位置分配。归因于其相似序列组成,有时,查询序列中的一个命中匹配于多个基序,或一个基序匹配于查询序列中的多个位置。
实施例23
图24显示使用exparna,比较(a)sineup-gfp中和(b)minisineup-gfp中asuchl1sineb2的2d结构。不同的加阴影区域表示两种rna之间序列和结构的相应精确匹配。minisineup-gfp中的asuchl1sineb22d结构几乎是相同,这表明这种结构在细胞内部高度稳定和占优势。唯一的结构变异区域以黑色矩形标记。
实施例24
图25显示(a)sineup-gfp中和(b)minisineup-gfp中asuxtsineb2-b的2d结构的exparna精确模式匹配结果。不同的加阴影区域表示两种rna之间序列和结构的相应精确匹配。这里,sineups和minisineups之间总体结构相当不同,但是一些区域保留可能对sineup功能至关重要的结构(例如,-加框的区域)。
实施例25
图26显示(a)minisineup-gfp中asuchl1sineb2和(b)sineup-gfp中astxnipsineb2的2d结构的exparna精确模式匹配结果和基序分析结果。不同的加阴影区域表示正合序列的相应独特模式和两种rna之间的结构匹配。全部sineb2s之间共有的相似序列和结构基序的位置以带有其相应基序编号的正方形指示。归因于其相似的序列组成,在一些情况下,基序2和3可以重叠,并且基序4、5、6和7同样如此。
实施例26
图27显示(a)minisineup-gfp中asuchl1sineb2和(b)sineup-gfp中asuxtsineb2-b的2d结构的exparna精确模式匹配结果和基序分析结果。同上者适用。
实施例27
图28显示(a)minisineup-gfp中asuchl1sineb2和(b)minisineup-gfp中asuxtsineb2-b的2d结构的exparna精确模式匹配结果和基序分析结果。不同的加阴影区域表示正合序列的相应独特模式和两种rna之间的结构匹配。同上者适用。
实施例28
图29显示(a)asuchl1sineb2和(b)minisineup-gfp中sineb2/b3共有(反向互补物)的2d结构的exparna精确模式匹配结果和基序分析结果。同上者适用。
实施例29
图30显示(a)asuchl1sineb2和(b)minisineup-gfp中asnars2sineb2的2d结构的exparna精确模式匹配结果和基序分析结果。同上者适用。
实施例30
图31显示(a)asuchl1sineb2和(b)minisineup-gfp中asabhd11sineb2的2d结构的exparna精确模式匹配结果和基序分析结果。同上者适用。
实施例31
图32显示(a)asuchl1sineb2和(b)minisineup-gfp中asepb4.9sineb2的2d结构的exparna精确模式匹配结果和基序分析结果。同上者适用。
实施例32
图33显示(a)asuchl1sineb2和(b)minisineup-gfp中aswfdc5sineb2的2d结构的exparna精确模式匹配结果和基序分析结果。同上者适用。
实施例33
图34显示(a)asuchl1sineb2和(b)minisineup-gfp中aspgbd1sineb2的2d结构的exparna精确模式匹配结果和基序分析结果。同上者适用。
实施例34
图35显示(a)asuchl1sineb2和(b)minisineup-gfp中asgsk3βsineb2的2d结构的exparna精确模式匹配结果和基序分析结果。同上者适用。
实施例35
图36显示(a)asuchl1sineb2和(b)minisineup-gfp中asrhodsineb2-mm2的2d结构的exparna精确模式匹配结果和基序分析结果。同上者适用。
实施例36
图37显示(a)asuchl1sineb2和(b)minisineup-gfp中asrhodsineb2-b3a的2d结构的exparna精确模式匹配结果和基序分析结果。同上者适用。
实施例37
图38显示(a)asuchl1sineb2和(b)minisineup-gfp中ase4f1sineb2-mm2的2d结构的exparna精确模式匹配结果和基序分析结果。同上者适用。
实施例38
图39显示(a)asuchl1sineb2和(b)minisineup-gfp中ase4f1sineb2-mm1t的2d结构的exparna精确模式匹配结果和基序分析结果。同上者适用。
实施例39
图40显示as-uchl1sineb2的结构动力学。将sineb2rna结构分成更小片段的目的是,全长(fl)太大,以至于不能分析当前nmr磁力。该结构需要分成1hnmr谱的正确分配的亚胺基质子信号。划分的rna应当形成与fl相似的结构。tm和c和mc2的1hnmr谱表明它们的确如此。此外,相对于化学足迹测量,这些结构在nmr水质缓冲液中差异地形成(图2)。结构动力学可以在生理条件(如不同细胞区室)发生(sun等人,biorxiv2018.,https://doi.org/10.1101/399386)。
图40a显示借助rna折叠程序的来自as-uchl1的flsineb2和其结构部分的计算预测结构。fl分成三个结构(tm、c和mc2)。
图40b显示借助nmr1h谱时残基的分配情况。确认mc2结构、c结构、tm结构的几乎全部信号均是fl结构。这些结果表明,mc2、c和tm形成与fl结构相同的结构。
图40c显示,五个茎结构(左侧处打点正方形标记的茎)在奥氏核效应光谱法(noesy)(右)时分配。
实施例40
图41显示as-uchl1sineb2的结构动力学。
fl和c的计算预测均在46-49和58-61形成茎结构((a)中打点的圆圈),这种结构未在图1和图2中借助化学足迹法确认。此外,50-57处的环和18-123处的单链区域未在图1和图2中借助化学足迹法确认。但是,1hnmr谱和noesy确认46-49处和58-61处的茎结构(图40b和40c)与计算预测结构相同。使用a51>c突变体rna和sineb2rna的部分,借助1hnmr谱,分析了46-49和58-61的茎结构是否与环50-57处和单链区域18-123处的任何其他非结构化残基相关。换句话说,研究了化学足迹测量未确定的结构是否在nmr缓冲液条件下同时出现及彼此缔合。
该实施例展示了
a)借助rnafold程序的fl和c结构域(34-122)的计算预测结构。c结构分成fl结构的解折叠区域(118-123)。c结构处的5个残基(118-120)产生具有相反残基的茎。
b)三个小rna(34-120、34-121和34-122)和fl的1hnmr谱。34-120未显示45-49和58-61的茎(u61、g60、g49、u48和g59的垂线处的峰),但其他三个结构(fl、34-121和34-122)显示这些峰。图41b显示来自118-123的解折叠环对46-49和58-61处形成茎重要。图41b显示增加蛋白质合成的a51c的1hnmr谱(图12)。作为a51>c突变的结果,化学位移在u48和g59的峰(垂线)处发生。另外,fl处两个未分配的峰(u61和u48的左侧处的垂线)消失。这些峰位移表明,a51处的突变改变46-49处茎的结合位置并且升高蛋白质合成功能。总之,50-57处和118-123处的环结构在nmr缓冲液(20mmmes(ph6.0),50mmnacl、8%d2o,298k)的含水条件下与46-49处和58-61处茎的结构形成过程相互作用。图41表明,sineb2结构不是严格的,但是一些结构是动态的。a中的圆圈对应于b中的圆圈。
实施例41
图42显示ed长度和结构基序组合各异的合成sineup怎样上调gfp蛋白表达。
在有义egfp和(a)sineups(n=3),和(b)minisineups(n=4)共转染后(转染后24小时),hek293t/17细胞中的平均gfp蛋白质倍数变化。蛋白质印迹图像和使用imagej软件计算的相应的gfp条带强度(对actb表达水平标准化)。使用空载体作为阴性对照,而使用asuchl1(含有sineup或minisineup的asuchl1sineb2)作为阳性对照。sineupid之后为括号中的ed长度。误差线±sd;****p<0.00005;***p<0.0005;**p<0.005;*p<0.05;双尾studentt检验。
下文提供结构基序组合(基于这个实验中测试的在图23中描述的基序)连同构建体的完整特征。出于清晰理由,seqidnos仅表示效应子结构域的序列,所述序列在sineup中和相应的minisineup中相同。
sineup071/minisineup071(206nt):从5′端,部分基序1、部分基序2和3(重叠)、完整基序6、部分基序4和7
sineup071
接收质粒:pcdna3.1(-)
克隆限制性位点:xhoi-hindiii
特征:bd=-40/+32(+21nt间隔序列)
ed(seqidno:69)=在sacii和clai限制性位点之间;206nt长的序列来自基于asuchl1、asuxt-b和astxnipsineb2元件的全局序列和结构比对的共有2d结构(+alu和来自asuchl1的3′尾区域)
sineup长度=1380nt
minisineup071
接收质粒:pcdna3.1(-)
克隆限制性位点:xhoi-hindiii
特征:bd=-40/+4(+21nt间隔序列)
ed=在ecori和hindiii限制性位点之间;(seqidno:69)
sineup长度=289nt
sineup072/minisineup072(206nt):从5′端起,部分基序2、完整基序1、部分基序4、完整基序7
sineup072
接收质粒:pcdna3.1(-)
克隆限制性位点:xhoi-hindiii
特征:bd=-40/+32(+21nt间隔序列)
ed(seqidno:70)=在sacii和clai限制性位点之间;206nt长的序列来自基于asuchl1和asuxt-bsineb2元件的全局序列和结构比对的共有2d结构(+alu和来自asuchl1的3′尾区域)
sineup长度=1380nt
minisineup072
接收质粒:pcdna3.1(-)
克隆限制性位点:xhoi-hindiii
特征:bd=-40/+4(+21nt间隔序列)
ed=在ecori和hindiii限制性位点之间;seqidno:70
sineup长度=289nt
sineup073/minisineup073(156nt):从5′端起,完整基序1、部分基序2和3(重叠)、部分基序4、完整基序6、部分基序7
sineup073
接收质粒:pcdna3.1(-)
克隆限制性位点:xhoi-hindiii
特征:bd=-40/+32(+21nt间隔序列)
ed(seqidno:71)=在sacii和clai限制性位点之间;156nt长的序列来自基于asuchl1和asuxt-bsineb2元件的局部序列和结构比对的共有2d结构(+alu和来自asuchl1的3′尾区域)
sineup长度=1330nt
minisineup073
接收质粒:pcdna3.1(-)
克隆限制性位点:xhoi-hindiii
特征:bd=-40/+4(+21nt间隔序列)
ed=在ecori和hindiii限制性位点之间;seqidno:71
sineup长度=239nt
sineup074/minisineup074(68nt):从5′端起,部分1、7、6(重叠),部分2
sineup074
接收质粒:pcdna3.1(-)
克隆限制性位点:xhoi-hindiii
特征:bd=-40/+32(+21nt间隔序列)
ed(seqidno:72)=在sacii和clai限制性位点之间;68nt长的序列来自基于asuchl1、asuxt-b和astxnipsineb2元件的局部序列和结构比对的共有2d结构(+alu和来自asuchl1的3′尾区域)
sineup长度=1242nt
minisineup074
接收质粒:pcdna3.1(-)
克隆限制性位点:xhoi-hindiii
特征:bd=-40/+4(+21nt间隔序列)
ed=在ecori和hindiii限制性位点之间;seqidno:72
sineup长度=151nt
sineup075/minisineup075(110nt):从5′端起,完整1、2、3和6
接收质粒:pcdna3.1(-)
克隆限制性位点:xhoi-hindiii
特征:bd=-40/+32(+21nt间隔序列)
ed(seqidno:73)=在sacii和clai限制性位点之间;110nt;δ28-32、δ42-60、δ90-116、δ131-135asuchl1sineb2缺失突变体(+alu和来自asuchl1的3′尾区域)
sineup长度=1284nt
minisineup075
接收质粒:pcdna3.1(-)
克隆限制性位点:xhoi-hindiii
特征:bd=-40/+4(+21nt间隔序列)
ed=在ecori和hindiii限制性位点之间;seqidno:73
sineup长度=193nt
asuchl1和asuchl1_δ5′-28nt(167nt):阳性对照,全部7个基序
这个实验中测试的全部基序组合均上调gfp表达,尽管无一超过阳性对照诱导的蛋白质倍数变化。这表明,不同基序对sineup活性具有组合性影响。如实施例22中解释,大部分基序具有可能代偿任何遗漏基序并确保最小sineup活性水平的相似序列组成。全部7个基序均按其相应顺序获得最佳sineup效果,如从阳性对照(asuchl1sineb2)显而易见。有趣地,sineup074和minisineup074,尽管ed短并因此缺少基序,但增加gfp蛋白表达并且代表sineup活性所要求的最小功能性序列实例。
实施例42
图43显示(a)minisineup-gfp中asuchl1sineb2(icshape-导向)和(b)minisineup071ed(rnafold预测)的2d结构的exparna精确模式匹配结果。不同的加阴影区域表示正合序列的相应独特模式和两种rna之间的结构匹配。全部sineb2s之间共有的相似序列和结构基序的位置以带有其相应基序编号的正方形指示。归因于其相似的序列组成,在一些情况下,基序2和3可以重叠,并且基序4、5、6和7同样如此。
实施例43
图44显示(a)minisineup-gfp中asuchl1sineb2(icshape-导向)和(b)minisineup072ed(rnafold预测)的2d结构的exparna精确模式匹配结果。同上者适用。
实施例44
图45显示(a)minisineup-gfp中asuchl1sineb2(icshape-导向)和(b)minisineup073ed(rnafold预测)的2d结构的exparna精确模式匹配结果。同上者适用。
实施例45
图46显示(a)minisineup-gfp中asuchl1sineb2(icshape-导向)和(b)minisineup074ed(rnafold预测)的2d结构的exparna精确模式匹配结果。同上者适用。
实施例46
图47显示(a)minisineup-gfp中asuchl1sineb2(icshape-导向)和(b)minisineup075ed(rnafold预测)的2d结构的exparna精确模式匹配结果。同上者适用。
实施例47
图48显示来自asuchl1的sineb2的结构片段怎样上调gfp蛋白表达。
(a)在有义egfp和sineups(n=2)共转染后(转染后24小时),hek293t/17细胞中的平均gfp蛋白质倍数变化。蛋白质印迹图像和使用imagej软件计算的相应的gfp条带强度(对actb表达水平标准化)。使用空载体作为阴性对照。
这个实验中测试的全部结构片段均上调gfp表达,尽管无一超过asuchl1sineb2诱导的蛋白质倍数变化。这些片段形成与asuchl1sineb2结构相同的结构(图23)。
这表明,具有不同基序的结构对sineup活性具有组合性影响。如图23中解释,大部分基序具有可能代偿任何遗漏基序并确保最小sineup活性水平的相似序列组成。全部7个基序均按其相应顺序获得最佳sineup效果,如从阳性对照(asuchl1sineb2)显而易见。
(b)在nmr光谱溶液中确认这种asuchl1sineb2结构为备选结构。其他片段保持作为asuchl1sineb2较短结构形成。基序组合对应于新图23。
下文提供结构基序组合(基于图23中描述的基序)连同构建体的完整特征。出于清晰理由,seqidnos仅表示效应子结构域的序列。
asuchl1sineb2:从5′端起,完整基序1、部分基序2、完整基序3、部分基序5、完整基序6和完整基序7
24-150(126nt):从5′端起,完整基序3、部分基序4、部分基序5、完整基序6和完整基序7
接收质粒:pcdna3.1(-)
克隆限制性位点:xhoi-hindiii
特征:bd=-40/+32(+21nt间隔序列)
ed(seqidno:74)=在sacii和clai限制性位点之间;126nt长的序列来自asuchl1sineb2元件(+alu和来自asuchl1的3′尾区域)
sineup长度=1300nt
c_34-122(89nt):从5′端起,部分基序4、5和完整基序6和完整基序7
接收质粒:pcdna3.1(-)
克隆限制性位点:xhoi-hindiii
特征:bd=-40/+32(+21nt间隔序列)
ed(seqidno:75)=在sacii和clai限制性位点之间;89nt长的序列含于asuchl1sineb2元件内(+alu和来自asuchl1的3′尾区域)
sineup长度=1263nt
tm(72nt):从5′端起,完整基序1、完整基序2、完整基序3、完整基序4和部分基序6(重叠)
接收质粒:pcdna3.1(-)
克隆限制性位点:xhoi-hindiii
特征:bd=-40/+32(+21nt间隔序列)
ed(seqidno:76)=在sacii和clai限制性位点之间;72nt长的序列含于asuchl1sineb2元件内(+alu和来自asuchl1的3′尾区域)
sineup长度=1246nt
mc2(94nt):从5′端起,完整基序3、完整基序4、部分基序5和完整基序7。接收质粒:pcdna3.1(-)
克隆限制性位点:xhoi-hindiii
特征:bd=-40/+32(+21nt间隔序列)
ed(seqidno:77)=在sacii和clai限制性位点之间;94nt长的序列来自asuchl1sineb2元件(+alu和来自asuchl1的3′尾区域)
sineup长度=1268nt
实施例48
图49显示作为环状rna表达时,sineup有活性。在hek293t/17细胞中,环状sineup-dj-1(circsineup-dj-1)能够以结合结构域依赖性方式增加内源dj-1蛋白质水平,同时靶dj-1mrna水平无变化。
这个实施例表明,sineup序列可以是环状rna并且这些序列在有内源mrna的功能性核酸分子中拥有翻译上调活性。在一级核苷酸序列相同,但线性sineup-dj-1的5’端和3’端共价连接的情况下,获得活性。图49a是含有规范序列倒转sineb2和针对dj-1mrna的结合结构域的环状sineup-dj-1rna(circsineup-dj-1)的示意图。图49b显示靶向内源表达的dj-1mrna的功能性核酸分子的翻译上调活性,其中sineup处于环状形式。将hek293t/17细胞用空载体或环状sineup-dj-1和缺少针对dj-1mrna的反义序列的环状sineup-dj-1deltabd转染。用抗dj-1抗体进行蛋白质印迹,以监测过量表达的蛋白质。使用β-肌动蛋白作为上样对照。在蛋白质印迹图像上对β-肌动蛋白标准化并相对于空白对照样品,计算诱导倍数。图49c展示了定量环状sineup-dj-1rna水平和dj-1mrna水平的实时qpcr分析,从而证明该机制为转录后。
优点
sineb2结构性元件的鉴定允许一种结构指导的选择并设计在人类中活性和潜在实用性增加的sineup变体的策略。特别地,出于众多原因,其中有体内递送和稳定性,较短sineup变体有利。
起点商标作为专业知识产权交易平台,可以帮助大家解决很多问题,如果大家想要了解更多知产交易信息请点击 【在线咨询】或添加微信 【19522093243】 与客服一对一沟通,为大家解决相关问题。
与客服一对一沟通,为大家解决相关问题。
此文章来源于网络,如有侵权,请联系删除

 热门咨询
热门咨询



