
 商标分类
商标分类  商标转让
商标转让 
抗体变体及其用途的制作方法
 2021-02-02 13:02:35|
2021-02-02 13:02:35| 252|
252| 起点商标网
起点商标网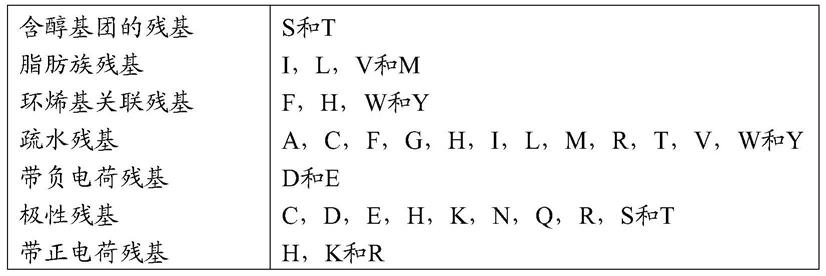
抗体变体及其用途
[0001]
本发明是基于申请日为2012年7月6日,申请号为“201610439143.1”,发明名称为“抗体变体及其用途”的专利申请的分案申请。
发明领域
[0002]
本发明涉及包含变体fc域的多肽及相关抗体。更具体地,本发明涉及包含fc域的抗体或多肽,其具有由fc域中一个或多个氨基酸修饰带来的修饰的效应子作用。
[0003]
发明背景
[0004]
由抗体fc域介导的效应子作用允许破坏外来实体,如杀灭病原体以及清除和降解抗原。抗体依赖性细胞介导细胞毒性(adcc)和抗体依赖性细胞介导吞噬(adcp)由fc区与具有fc受体(fcr)的细胞的结合起始,而补体依赖性细胞毒性(cdc)由fc区与c1q的结合起始,这起始补体激活的经典途径。
[0005]
每个igg抗体包含两个c1q结合位点,每个重链恒定(fc)区中各一个。然而,由于单体igg对c1q的亲和力非常弱(k
d
~10-4
m),溶液中的单个igg分子不激活补体(sledge等,1973j.biol.chem.248,2818-13;hughes-jones等,1979mol.immunol.16,697-701)。抗原驱动的igg关联可导致多价c1q分子更紧密结合(k
d
~10-8
m)和补体激活(burton等,1990mol.immunol.22,161-206)。相比之下,igm以共价连接的五或六聚体天然存在,与细胞表达或固定的抗原结合后,igm五聚体和六聚体可以有效的引起cdc。抗原结合是诱导igm构象变化以暴露c1q结合位点必需条件(feinstein等,1986,immunology today,169-174)。
[0006]
已经提出通过fc区ch2/ch3域的相互作用形成六聚体环结构,igg也能完成补体激活(burton等,1990trends in biochem.sci.15,64-69)。支持此六聚体igg结构存在的证据已经在二维(reidler等,1986i handbook of experimental immunology第4版.(weir,d.m.编),pp17.1-17.5.blackwell,edinburgh;pinteric等,1971immunochem.8,1041-5)和三维晶体中,以及对溶液中的igg1、igg2a和igg4和人fc找到(kuznetsov等,2000j struct.biol.131,108-115)。六聚体环的形成也在针对抗hiv-1gp120的b12人igg1κ抗体的晶体结构(pdb中的1hzh)中观察到(saphire等,science 2001aug 10;293(5532),1155-9)。在b12六聚体环中,六个易接近的c1q结合位点位于六聚体表面,一个来自于六个抗体中的每个抗体,而其他六个结合位点面向下。
[0007]
c1q类似六球头的郁金香束,包含系于六个胶原杆上的抗体结合区[perkins等,1985biochem j.228,13-26;poon等,1983j mol biol.168,563-77;reid等,1983biochem soc trans 11,1-12;weiss等,1986j.mol.biol.189,573-81]。发现c1q与1hzh晶体结构的b12六聚体组装配合,以使六个球头中每一个与六个c1q结合位点中的一个相接触(parren,faseb summer research conference,snowmass,co.,5-10july 2010;"crystal structure of an intact human igg:implications for hiv-1neutralization and effector function",erica ollmann saphire著,the scripps research institute,la jolla,california.november 2000)。晶体结构中对称相关的b12抗体之间观察到的fc界面中所选氨基酸的突变被发现降低c1q结合亲和力,表明了这些氨基酸对分子间fc:fc相互作
用的贡献。
[0008]
us 2011/0123440描述了改变的抗体fc区及其用途。改变的fc区有一个或多个氨基酸取代。
[0009]
us 2008/0089892描述了多肽fc区变体及包含这些fc区变体的组合物。
[0010]
us 2010/0184959描述了提供有改变的fc配体识别和/或效应子作用的fc多肽变体的方法。
[0011]
us 2010/015133描述了通过调节多肽关联来生产多肽的方法。
[0012]
us 2010/105873描述了生成多域蛋白疗法的综合方法。
[0013]
us 6,737,056描述了有改变的效应子作用的多肽变体。
[0014]
之前已做出努力以识别具有增强的效应子作用或其他修饰特性的抗体fc变体。这样的研究集中于,如,在igg同种型之间交换片段以产生嵌合igg分子(natsume等,2008cancer res 68(10),3863-72),或者在铰链区(dall'acqua等,2006j immunol 177,1129-1138)或在ch2域的c1q结合位点处或其附近,围绕残基d270、k322、p329和p331(idusogie等,2001j immunol 166,2571-2575;michaelsen等,2009scand j immunol 70,553-564和wo 99/51642)的氨基酸取代。例如,moore等(2010mabs 2(2),181-189)描述了检测s267e、h268f、s324t、s239d、i332e、g236a和i332e的各种组合的通过cdc或adcc的增强的效应子作用。影响与fc受体结合(wo 2006/105062,wo00/42072,美国专利6,737,056和美国专利7,083,784)或抗体物理特性(wo2007/005612 al)的其他fc突变也已经提出。
[0015]
尽管本领域中有这些和其他进步,然而,仍然有对新的和改进的基于抗体疗法的需求。
[0016]
发明概述
[0017]
本发明提供了相比其亲本多肽/抗体其具有增强的效应子作用的多肽和抗体变体。不限于理论,认为变体能在两个多肽/抗体分子fc区之间有更稳定的连接相互作用,由此提供更亲和的表面,其能导致增强的效应子作用,如提高的或更特异的cdc应答。特定变体的特征也在于改进的adcc应答、adcp应答、和/或其他增强的效应子作用。如本发明所述,多肽/抗体工程的这种微妙机制可以被应用于,例如提高基于抗体疗法的效力或特异性。
[0018]
因此本发明的一个方面涉及包含免疫球蛋白的fc域和结合区的亲本多肽的变体,其中变体在选自对应人igg1重链fc区中的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447的那些氨基酸残基的至少一个氨基酸残基处包含突变,条件是在s440的突变为s440y或s440w。
[0019]
本发明也提供至少一个这样的突变的用途,当多肽或抗体结合于例如抗原表达细胞、细胞膜或病毒粒子表面上的其抗原上时,提高由多肽或抗体介导的效应子作用。
[0020]
在一个本发明中被称为“单突变体”的方面,该变体相比亲本多肽或抗体具有提高的效应子作用。
[0021]
在一个本发明中被称为“双突变体”的方面,该变体在所述片段中包含至少两个突变,并相比仅包含两个突变中一个的变体、亲本多肽或抗体,或两者具有改进的效应子作用。
[0022]
在一个本发明中被称为“混合突变体”的方面,该变体在与相同或不同多肽或抗体在所述区段的不同氨基酸残基处包含突变的第二变体组合使用时,相比一种或多种变体、
第二变体和单独亲本多肽或抗体提供提高的效应子作用。
[0023]
通常,突变是氨基酸取代,如突变将亲本氨基酸残基交换为具有不同大小和/或促进新分子间fc:fc键形成或提高现存对相互作用强度的物理化学特性的氨基酸。对于根据本发明的突变的示例性氨基酸残基,连同示例性氨基酸取代示于表1和2a和b中。本发明不同方面的非限制性展示在图1中提供。
[0024]
本发明的这些和其他方面,具体是抗体变体的多种用途和治疗应用,在下面更详细地描述。
[0025]
附图简述
[0026]
图1:(a)六聚体形式igg分子的示意性展示。虚线圈显示了两个相邻igg分子的两个接近的fc:fc相互作用对。框中的箭头显示了观看b、c和d中图示的方向:两个邻近的fc分子旋转90
°
(在图的平面中)并从ch3域方向的fab臂观看。(b)观察到的寡聚增强突变对cdc的效果。示意性展示显示了根据本发明单突变体和双突变体方面具有增强效力的fc:fc相互作用对。(c)观察到的寡聚抑制突变对cdc的效果。示意性展示显示了根据本发明双突变体和混合突变体方面,相互补偿的至少两个寡聚抑制突变怎样组合入一个分子(双突变体方面)或者在两个分子上分离(混合突变体方面),以恢复或提高fc:fc相互作用。混合突变体实现依赖于两个抗体结合的特定效应子作用,这可以识别不同的靶标。(d)c1q结合抑制突变对cdc的理论效果。fc:c1q相互作用的示意性展示显示了由于c1q不能补偿引入抗体的缺陷,如果突变抑制c1q结合,其不能组合或混合以恢复cdc活性。
[0027]
图2:对应igg1重链中残基p247到k447的人igg1,igg1f,igg2,igg3和igg4 fc片段的比对,使用clustel 2.1软件,按kabat中所列eu索引编号。显示的序列代表人igg1重链恒定区域(seq id no:1;uniprot获取号p01857)和igg1m(f)(seq id no:5)的同种异型变体的残基130到330;igg2重链恒定区域的残基126到326(seq id no:2;uniprot获取号p01859);igg3重链恒定区域的残基177到377(seq id no:3;uniprot获取号p01860);和igg4重链恒定区域的残基127到327(seq id no:4;uniprot获取号p01861)。
[0028]
图3a和b:igg1(seq id no:6)、igg4(seq id no:8)和(部分)igg3(seq id no:7)骨架中的抗egfr抗体2f8的序列比对。描述了按照kabat和按照eu索引的氨基酸编号(两者都在kabat等,sequences of proteins of immunological interest,第5版,public health service,national institutes of health,bethesda,md.(1991)中描述)。
[0029]
图4:多聚体(如六聚体)排布中邻近分子(分别为fc和fc
’
)fc之间k439/s440相互作用详细视图,显示了野生型、未修饰的fc和fc
’
分子之间的相互作用。
[0030]
图5:多聚体(如六聚体)排布中邻近分子(分别为fc和fc
’
)fc之间k439/s440相互作用详细视图,显示了包含k439e和s440k突变的变体fc和fc
’
分子之间的相互作用。
[0031]
图6:用7d8 fc:fc突变体的c1q结合elisa。系列浓度的所示抗体被涂覆到微量滴定板孔上并与固定浓度的c1q温育。除了i253d,所有包被的变体与c1q结合的效率可与野生型7d8相比。显示了至少3个实验的示意。
[0032]
图7:7d8变体介导的对cd20阳性raji细胞的cdc。raji细胞与7d8变体(k439e,s440k,k439e/s440k双突变体,k439e+s440k混合)和系列浓度的c1q温育以通过测量细胞裂解测试cdc效力。显示了重复实验的示意图。
[0033]
图8:7d8变体(7d8-wt,k439e,s440k,k439e/s440k双突变体,k439e+s440k混合)介
导的对cd20阳性daudi细胞的cdc。测试了系列浓度的7d8变体的诱导cdc的效力。
[0034]
图9:cd38抗体humab005突变体介导的对cd38阳性细胞的cdc。(a)005突变体系列浓度对daudi细胞的cdc效力。(b)humab 005突变体系列浓度对raji细胞的cdc效力。(c)具有20%或50%nhs的humab 005的e345r突变体对wien133细胞的cdc效力。(d)具有20%或50%nhs的humab 005和7d8的e345r突变体对raji细胞的cdc效力。
[0035]
测试从瞬时转染中分离的未纯化抗体样品。使用模拟转染细胞的上清作为阴性对照。
[0036]
图10:与fc结合肽的竞争性实验中,cd38抗体humab 005(a)和cd20抗体humab 7d8(b)的野生型和e345r突变体的cdc。cdc后对与系列浓度fc结合dcawhlgelvwct肽(seq id no:7)温育的抗体调理的daudi细胞测量细胞裂解。使用从瞬时转染中分离的未纯化抗体样品。使用模拟转染细胞的上清作为阴性对照。
[0037]
图11:野生型cd38抗体humab 005和突变体igg1-005-e345r的cd38表达daudi细胞的adcc。表示为%裂解,显示了一个供体pbmc的adcc。
[0038]
图12a-c:在ph6,如通过elisa确定的,野生型的igg1-7d8和变体igg1-7d8-e345r与人、猴和小鼠fcrn的结合。
[0039]
图13:在scid小鼠中静脉注射后,野生型igg1-7d8和-e354r,-s440k和k322a变体的血浆浓度。
[0040]
图14a、b、c和d:对cd20和cd38阳性wien133细胞的cdc。
[0041]
图15a和b:在使用raji-luc#2d1细胞的皮下移植肿瘤模型中igg1-7d8-e345r的体内效力的评价。
[0042]
图16a和b:在使用raji-luc#2d1细胞的皮下移植肿瘤模型中igg1-005-e345r的体内效力的评价。
[0043]
图17:具有e345r突变的cd38/egfr双特异性抗体对cd38阳性、egfr阴性wien133细胞的cdc。
[0044]
图18a和b:具有或没有e345r突变的cd20/cd38双特异性抗体对cd20阳性、cd38阴性的wien133细胞或raji细胞的cdc。
[0045]
图19:具有e345r突变的egfr抗体2f8对egfr阳性a431细胞的cdc。
[0046]
图20a和b:e345r突变体抗体介导的cdc。
[0047]
图21:具有溶酶体标记lamp1(apc)的tf抗体(fitc)的共定位分析。
[0048]
图22a-d:引入e345r突变导致与在不同b细胞系中测试的野生型利妥昔单抗相比,cdc介导的杀伤增强。
[0049]
图22e:引入e345r导致与野生型利妥昔单抗相比最大cdc介导的杀伤提高,其独立于具有可比较cd20表达水平的不同b细胞系中补体调节蛋白cd46(a)、cd55(b)或cd59(c)的表达水平。
[0050]
图23a-d:cdc动力学。相比野生型抗体,e345r抗体导致更迅速和更大量的由cdc所致的靶细胞裂解。
[0051]
图24:cdc动力学。在双特异性cd38xcd20抗体中引入e345r突变导致更迅速和更大量的cdc介导的靶细胞裂解。
[0052]
图25:cdc动力学。在单价结合egfr阴性raji细胞的双特异性抗体cd38xegfr(a)和
cd20xegfr(b)中引入e345r突变导致更迅速和更大量的cdc介导的靶细胞裂解。
[0053]
图26a-f:野生型抗体与含(a-c)e345r和q386k或(d-f)e345r、e430g和q386k的突变体抗体的组合对wien133细胞的cdc。igg1-b12突变体不结合wien133细胞并被用作阴性对照抗体。
[0054]
图27a-b:含e345r突变的igg1、igg2、igg3和igg4同种型抗体的cdc效力。
[0055]
图28a-b:在野生型cd38抗体005中引入fc-fc稳定e345r突变导致离体cdc测定法中原代cll细胞杀伤增强(平均
±
均值的标准偏差)。
[0056]
发明详述
[0057]
如本发明中所述,令人惊奇地,不直接参与fc:c1q结合的氨基酸中的突变仍然能提高抗体的cdc,并也能改进抗体的其他fc介导的效应子作用。这支持了抗体分子如igg1抗体能形成随后被c1q连接的寡聚结构的假设。此外,尽管发现某些突变降低cdc诱导,相同或不同抗体分子中这样突变的某些组合导致cdc诱导的恢复,并显示对抗体寡聚的更多特异性,并由此促进更特异的cdc诱导。如实施例中所示,提高cdc应答的特定突变特征也在于改进的adcc应答,提高的亲和力,提高的内化和在鼠肿瘤模型系统中的体内效力。这些发现允许有增强cdc诱导能力,更有选择性的cdc诱导,和/或其他改进的效应子作用的新的基于抗体的疗法。
[0058]
本发明的抗体变体全包含抗原结合区以及全长或部分fc区,其在对应igg1中氨基酸残基p247到k447的区段中包含至少一个突变。不限于理论,基于图1中示意性展示并在本发明中被称为“单突变体”、“双突变体”和“混合突变体”的三个不同的原理,认为识别的突变导致更有效和/或更特异的cdc诱导。
[0059]
来自本发明变体的改进的c1q和/或cdc效应主要仅是在允许抗体寡聚体形成的检测中,如在抗原没有固定但位于流体膜上的基于细胞的检测中能检测到。此外,从更稳定的抗体寡聚体得到而不是从c1q直接结合位点的修饰得到的这些效应可以根据图1c所示的原理验证。
[0060]
定义
[0061]
术语“单突变体”应理解为相比亲本多肽或抗体具有提高的效应子作用的本发明变体。
[0062]
术语“双突变体”应理解为变体,其在所述片段中包含至少两个突变,并相比仅包含两个突变中一个的变体、亲本多肽或抗体或两者具有改进的效应子作用。
[0063]
术语“混合突变体”应理解为变体,其在与相同或不同多肽或抗体的包含所述区段的不同氨基酸残基处的突变的第二突变体组合使用时,相比一种或多种变体、第二变体和单独亲本多肽或抗体提供提高的效应子作用。
[0064]
术语“包含免疫球蛋白fc域和结合区的多肽”在本发明上下文中是指包含免疫球蛋白fc域和结合区的多肽,其能与例如位于细胞,细菌,或病毒粒子上任何分子,如多肽结合。免疫球蛋白的fc域定义为通常在用木瓜蛋白酶消化抗体后生成的抗体片段(这对本领域技术人员已知),其包括免疫球蛋白的两个ch2-ch3区和连接区,例如铰链区。抗体重链恒定区定义抗体同种型,例如igg1、igg2、igg3、igg4、iga1、iga2、ige。fc域介导抗体与被称为fc受体的细胞表面受体和补体系统蛋白的效应子作用。结合区可以是能连接到细胞、细菌、病毒粒子的多肽序列,如蛋白、蛋白配体、受体、抗原结合区或配体结合区。如果结合区是例
如受体,“包含免疫球蛋白fc域和结合区的多肽”可以作为免疫球蛋白fc域和所述结合区的融合蛋白来被制备。如果结合区是抗原结合区,“包含免疫球蛋白fc域和结合区的多肽”可以是抗体,像人抗体或仅有重链的抗体或scfv-fc-融合体。包含免疫球蛋白fc域和结合区的多肽通常包括连接区,例如铰链区,和免疫球蛋白重链的两个ch2-ch3区,因此“包含免疫球蛋白fc域和结合区的多肽”可以是“包含至少免疫球蛋白fc域和结合区的多肽”。术语“免疫球蛋白fc域”在本发明上下文中意指取决于抗体亚型的连接区,例如铰链,以及免疫球蛋白ch2和ch3区存在,例如人igg1、igg2、igg3、igg4、igd、iga1、igga2或ige。
[0065]
本发明中使用的术语“ch2区”或“ch2域”意指免疫球蛋白的ch2区。因此例如人igg1抗体的ch2区对应于根据eu编号系统的氨基酸228-340。然而,ch2区也可以是本发明所述的任何其他亚型。
[0066]
本发明中使用的术语“ch3区”或“ch3域”意指免疫球蛋白的ch3区。因此例如人igg1抗体的ch3区对应于根据eu编号系统的氨基酸341-447。然而,ch3区也可以是本发明所述的任何其他亚型。
[0067]
术语“免疫球蛋白”是指一类结构上相关的糖蛋白,其由两对多肽链,一对轻(l)低分子量链和一对重(h)链组成,所有四条之间通过二硫键内在相连。免疫球蛋白的结构已经被充分表征。见例如fundamental immunology ch.7(paul,w.,编第二版.raven press,n.y.(1989))。简而言之,每条重链通常由重链可变区(本发明中缩写为vh)和重链恒定区组成。重链恒定区通常由三个域,ch1,ch2,和ch3组成。重链之间通过所谓“铰链区”中的二硫键相连。每条轻链通常由轻链可变区(本发明中缩写为vl)和轻链恒定区组成。轻链恒定区通常由一个域,cl组成。vh和vl区可以被进一步再划分为高可变性区(或在序列和/或结构限定环形式中高度可变的高可变区),也被称为互补决定区(cdrs),其散布有更保守的区域,被称为框架区(frs)。每条vh和vl通常由三个cdrs和四个frs组成,其从氨基末端到羧基末端以以下顺序排列:fr1,cdr1,fr2,cdr2,fr3,cdr3,fr4(也见chothia和lesk j.mol.biol.196,901 917(1987))。除非另有说明或上下文相矛盾,本发明中恒定区序列的氨基酸根据eu-索引编号(在kabat,e.a.等,sequences of proteins of immunological interest.第五版-us department of health and human services,nih publication no.91-3242,pp 662,680,689(1991)中描述)。
[0068]
本发明上下文中术语“抗体”(ab)是指免疫球蛋白分子,免疫球蛋白分子片段,或两者的衍生物,其具有在典型生理条件下特异性结合抗原的能力,有显著时间段的半衰期,如至少约30分钟,至少约45分钟,至少约1小时,至少约2小时,至少约4小时,至少约8小时,至少约12小时,约24小时,约48小时,约3,4,5,6,7或更多天等,后任何其他相关功能限定时段(如足以诱导、促进、增强和/或调节与抗体结合抗原相关的生理应答的时间,和/或足以使抗体募集效应活性的时间)。本发明的抗体包含免疫球蛋白fc域和结合区。抗体通常含有两个ch2-ch3区和连接区,例如铰链区,例如至少fc域。因此本发明的抗体可以包含fc区和抗原结合区。免疫球蛋白分子的重链和轻链可变区包含与抗原相互作用的结合域。抗体的恒定或“fc”域可介导免疫球蛋白与宿主组织或因子的结合,包括免疫系统各种细胞(如效应细胞)和补体系统的组分如c1q,补体激活经典途径的第一组分。抗体也可以是多特异性抗体,如双特异性抗体或类似分子。术语“双特异性抗体”是指具有对至少两种不同,通常是不重叠表位的特异性的抗体。这样的表位可以在相同或不同的靶标上。如果表位在不同的
靶标位上,这样的靶标可以在相同细胞或不同细胞或细胞类型上。如上所述,除非另有说明或者明显与上下文相矛盾,在本发明中术语抗体包括抗体片段,其包含至少部分fc区并保持特异性结合抗原的能力。这样的片段可以通过任何已知技术提供,如酶切、肽合成和重组表达技术。已经显示了抗体的抗原结合功能可以由全长抗体的片段来进行。涵盖在术语“ab”或“抗体”中的结合片段实例包括,不限于,单价抗体(在wo2007059782中由genmab描述);重链抗体,其仅由两条重链组成并在例如骆驼中天然存在(例如,hamers-casterman(1993)nature 363:446);thiomabs(roche,wo2011069104),链交换工程域(seed或seed体),其是不对称的和双特异性的抗体样分子(merck,wo2007110205);triomab(fresenius,lindhofer等(1995j immunol 155:219));fcδadp(regeneron,wo2010151792),azymetric scaffold(zymeworks/merck,wo2012/058768),mab-fv(xencor,wo2011/028952),双可变域免疫球蛋白(abbott,dvd-ig,美国专利7,612,181);双域双头抗体(unilever;sanofi aventis,wo20100226923),二双抗体(imclone/eli lilly),钮-入-孔(knob-into-holes)抗体形式(genentech,wo9850431);duobody(genmab,wo2011/131746);静电驾驭抗体形式(amgen,ep1870459和wo 2009089004;chugai,us201000155133;oncomed,wo2010129304a2);双特异性igg1和igg2(rinat neurosciences corporation,wo11143545),crossmabs(roche,wo2011117329),luz-y(genentech),biclonic(merus),双靶向域抗体(gsk/domantis),识别两个靶标的二合一抗体(genentech,novlmmune),交联mabs(karmanos cancer center),covx体(covx/pfizer),igg样双特异性(imclone/eli lilly,shen,j.,等j immunol methods,2007.318(1-2):p.65-74),和dig体和pig体(pharmabcine),以及双亲和性再靶向分子(macrogenics的fc-dart或ig-dart,wo/2008/157379,wo/2010/080538),zybodies(zyngenia),普通轻链(crucell/merus,us7262028)或普通重链(novlmmune的κλ体)的方法,以及融合蛋白,其包含多肽序列,所述多肽序列融合于含fc域的抗体片段,像scfv融合体,像zymogenetics/bms的bsab,biogen idec的hercules(us007951918),emergent biosolutions/trubion的scorpions,ts2ab(medlmmune/az dimasi,n.,等j mol biol,2009.393(3):p.672-92),novartis的scfv融合体,changzhou adam biotech inc的scfv融合体(cn 102250246),roche的tvab(wo2012025525,wo2012025530),f-star的mab
2
(wo2008/003116)和双scfv-融合体。还应当理解,术语抗体,除另有指明外,也包括多克隆抗体、单克隆抗体(如人单克隆抗体)、抗体混合物(重组多克隆)例如,通过symphogen和merus(oligoclonics)开发的技术生成的,和抗体样多肽,如嵌合抗体和人源化抗体。生成的抗体可潜在地具有任何同种型。
[0069]
术语“全长抗体”,当在本发明中使用时是指抗体(如亲本或变体抗体),其包含对应通常在该同种型野生型抗体中发现的那些域的所有重和轻链恒定和可变域。
[0070]
本发明中所用的术语“人抗体”是包括抗体,其具有源自人种系免疫球蛋白序列的可变区和恒定区。本发明的人抗体可以包括不由人种系免疫球蛋白序列编码氨基酸残基(例如,通过体外随机或点特异突变或者通过体内体细胞突变引入的突变、插入或缺失),然而,本发明中所用的术语“人抗体”不意欲包括其中源自另一哺乳动物种类种系,如小鼠的cdr序列已经被移植到人骨架序列的抗体。
[0071]
本发明中所用术语“单克隆抗体”、“单克隆ab”、“单克隆抗体组成”,“mab”或类似是指单分子组成的抗体分子制剂。单克隆抗体组成显示对特定表位的单结合特异性和亲和
力。因此,术语“单克隆抗体”是指显示单结合特异性的抗体,其具有源自人种系免疫球蛋白序列的可变和恒定区。人mabs可以由杂交瘤细胞生成,杂交瘤细胞包括从转基因或转染色体非人动物,如转基因小鼠中获得的b细胞,b细胞包含重排以产生功能性人抗体的人重链转基因部分和轻链转基因部分并与永生细胞融合。
[0072]
本发明中所用的“同种型”是指按重链恒定区基因编码的免疫球蛋白类(例如iggl、igg2、igg3、igg4、igd、iga1、igga2、ige或igm或者其任何同种异型类型如igg1m(za)和igg1m(f))。此外,每种重链同种异型可与κ或λ轻链组合。
[0073]
术语“单价抗体”在本发明上下文中意为抗体分子,其仅能够仅以一个抗体结合域与抗原结合,例如具有单抗原-抗体相互作用,并因此不能抗原交联。
[0074]
本发明中所用的术语“靶标”在本发明上下文中应理解为包含fc域和结合区的多肽的结合区结合到的分子,当用于抗体结合的上下文中时包括所提出抗体导向的任何抗原。术语“抗原”和“靶标”关于抗体时可以互换使用,并对本发明任何方面或实施方案构成相同的含义或目的。
[0075]
本发明中所用的术语“结合”在抗体结合到预定抗原的上下文中通常是结合,当使用抗原作为配体和抗体作为分析物,通过例如表面等离子共振(spr)技术在biacore3000仪器中测定时,结合有对应约10-6
m或更小,例如10-7
m或更小,如10-8
m或更小,如10-9
m或更小,约10-10
m或更小,或约10-11
m或甚至更小的k
d
的亲和力,并且以对应于与结合到非预定抗原或密切相关抗原的非特异性抗原(例如bsa,酪蛋白)时其亲和力相比为至多1/10(at least ten-fold lower),如至多1/100,例如至多1/1000,如至多1/10000,例如至多1/100000的k
d
的亲和力与预定抗原结合。亲和力较低时的量取决于抗体的k
d
,这样当抗体的k
d
非常低(即抗体高度特异)时,则对抗原亲和力相比对非特异性抗原亲和力较低的量可以是至少10000倍。本发明中所用的术语“k
d”(m)是指特定抗体-抗原相互作用的解离平衡常数。
[0076]
本发明的“变体”或“抗体变体”或“亲本抗体变体”是相比“亲本抗体”包含一个或多个突变的抗体分子。类似的,本发明“变体”或“包含免疫球蛋白fc域和结合区的多肽的变体”或“包含免疫球蛋白fc域和结合区的亲本多肽的变体”是相比“包含免疫球蛋白fc域和结合区的亲本多肽”,包含一个或多个突变的“包含免疫球蛋白fc域和结合区的多肽”。不同的术语可以互换使用,并对本发明任何方面或实施方案构成相同的含义或目的。示例性的亲本抗体形式包括,不限于,野生型抗体,全长抗体或含fc的抗体片段,双特异性抗体,人抗体,或其任意组合。示例性的突变包括亲本氨基酸序列中的氨基酸缺失、插入或氨基酸取代。氨基酸取代可将天然氨基酸替换为另一种天然存在的氨基酸,或者非天然存在的氨基酸衍生物。氨基酸取代可以是保守或非保守的。在本发明上下文中,保守取代由下面三个表的一个或多个中反映的氨基酸类型之间的取代所定义:
[0077]
保守取代的氨基酸残基类型
[0078]
酸性残基asp(d)和glu(e)碱性残基lys(k),arg(r),和his(h)亲水不带电荷残基ser(s),thr(t),asn(n)和gln(q)脂肪族不带电荷残基gly(g),ala(a),val(v),leu(l)和ile(i)非极性不带电荷残基cys(c),met(m),和pro(p)芳香族残基phe(f),tyr(y),和trp(w)
[0079]
可选保守氨基酸残基取代类型
[0080]
1ast2de 3nq 4rk 5ilm6fyw
[0081]
可选的氨基酸残基物理和功能分类
[0082][0083][0084]
本发明上下文中,变体中的取代表示为:
[0085]
原氨基酸-位置-取代的氨基酸
[0086]
使用三字母码或单字母码,包括编码xaa和x来表示氨基酸残基。因此,标记“e345r”或“glu345arg”意为,在对应亲本抗体345位置氨基酸的变体氨基酸位置包含以精氨酸取代谷氨酸的取代的变体。当两者如下所示比对时,
[0087]
当位置本身不存在于抗体中,但变体包含氨基酸插入时,例如:
[0088]
位置-取代的氨基酸;使用标记,例如“448e”。
[0089]
这样的标记与同源多肽或抗体系列中的修饰特别相关联。
[0090]
类似的,当取代氨基酸残基的种类不重要时:
[0091]
原氨基酸-位置;或“e345”。
[0092]
对于原氨基酸和/或取代的氨基酸可以包含多于1个,但非全部氨基酸的修饰,位置345处谷氨酸取代为精氨酸、赖氨酸或色氨酸:
[0093]“glu345arg,lys,trp”或“e345r,k,w”或“e345r/k/w”,或“e345至r,k或w”可以在本发明上下文中互换使用。
[0094]
此外,术语“取代”包括取代为其他19个天然氨基酸的任一个,或取代为其他氨基酸,如非天然氨基酸。例如,345位置处氨基酸e的取代包括下面取代的每一个:345a,345c,345d,345g,345h,345f,345i,345k,345l,345m,345n,345q,345r,345s,345t,345v,345w和345y。顺便说一下,这相当于名称345x,其中x指任意氨基酸。这些取代也可以被称为e345a,
e345c等,或e345a,c等或e345a/c/等。同样的情况适用于本发明中涉及的每一个的类似情况和每个位置,本发明中具体地包括这样取代的任一个。
[0095]“对应于”另一个序列中的氨基酸或片段的一个序列中的氨基酸或片段是(i)使用标准序列比对程序如align,clustalw或类似的,通常以默认设置与另一氨基酸或片段比对,和(ii)具有与seq id no:1至少50%,至少80%,至少90%,或至少95%的序列同一性的,氨基酸或片段。例如可以使用图2和3中所显示的序列比对以鉴定igg2、igg3或igg4 fc序列中对应于igg1 fc序列中特定氨基酸的任意氨基酸。
[0096]
本发明涉及变体,即亲本抗体,和/或变体抗体,其与seq id nos:1,2,3,4和5的氨基酸p247至k447具有一定程度的同一性,这样的亲本和/或变体抗体在下文中被称为“同源抗体”。
[0097]
为了本发明的目的,通过needleman-wunsch比对(即全局比对)的程序“比对”来确定两个氨基酸序列之间的同一性程度以及两个核苷酸序列之间的同一性程度。该程序被用于多肽以及核苷酸序列的比对。默认得分矩阵blosum50被用于多肽比对,而默认同一性矩阵被用于核酸序比对,间隙的第一个残基罚分是多肽-12和核苷酸-16,间隙的其他残基的罚分是多肽-2和核苷酸-4。
[0098]“比对”是fasta包版本v20u6的一部分(见w.r.pearson和d.j.lipman(1988),"improved tools for biological sequence analysis",pnas85:2444-2448,以及w.r.pearson(1990)"rapid and sensitive sequence comparaison with fastp and fasta",methods inenzymology 183:63-98)。fasta蛋白比对使用smith-waterman算法,对间隙大小无限制(见"smith-waterman algorithm",t.f.smith和m.s.waterman(1981)j.mol.biolo.147:195-197)。
[0099]
本发明中所用的术语“效应细胞”是指相对免疫应答识别和激活阶段,参与免疫应答效应阶段的免疫细胞。示例性的免疫细胞包括骨髓或淋巴来源的细胞,例如淋巴细胞(如包含溶细胞性t细胞(ctls)在内的b细胞和t细胞),杀伤细胞,天然杀伤细胞,巨噬细胞,单核细胞,嗜酸性粒细胞,多形核细胞,如中性粒细胞,粒细胞,肥大细胞,和嗜碱性粒细胞。一些效应细胞表达fc受体(fcrs)或补体受体并执行特异免疫功能。在一些实施方案中,效应细胞如,例如天然杀伤细胞,能诱导adcc。例如,表达fcrs的单核细胞,巨噬细胞,中性粒细胞,树突状细胞和kupffer(枯否氏)细胞参与靶细胞的特异性杀伤和将抗原呈递到免疫系统其他组分上,或结合到呈递抗原的细胞上。在一些实施方案中,能得到靶细胞激活c3片段累积的抗体驱动经典补体激活可以进一步增强adcc。c3裂解产物是表达在骨髓细胞上的补体受体(crs),如cr3的配体。效应细胞上crs的补体片段识别可以促进增强的fc受体介导adcc。在一些实施方案中,抗体驱动的经典补体激活导致靶细胞上的c3片段。这些c3裂解产物可以促进直接补体依赖性细胞细胞毒性(cdcc)。在一些实施方案中,效应细胞可以吞噬靶标抗原,靶标颗粒或靶细胞。特定fcr或补体受体在效应细胞上的表达可能受体液因子如细胞因子调节。例如,已经发现fcγri的表达被干扰素γ(ifnγ)或g-csf上调。这样增强的表达提高了带有fcγri的细胞对靶标的细胞毒性。效应细胞可以吞噬靶标抗原或吞噬或裂解靶细胞。在一些实施方案中,抗体驱动的经典补体激活导致靶细胞上的c3片段。这些c3裂解产物可以通过效应细胞促进直接吞噬或者通过增强抗体介导的细胞吞噬间接吞噬。
[0100]
本发明中所用术语“载体”意思是指能诱导连接入载体的核酸片段转录的核酸分
子。载体的一种类型是“质粒”,其为环形双链dna环形式。载体的另一种类型是病毒载体,其中核酸片段可以被连接入病毒基因组。某些载体能在其被引入的宿主细胞中自主复制(例如有细菌复制原点的细菌载体和附加体哺乳动物载体)。其他载体(如非附加体哺乳动物载体)可以在引入宿主细胞时被整合入宿主基因组,并由此随宿主基因组一起复制。此外,某些载体能引导可操作地与其连接的基因的表达。这样的载体本发明中被称为“重组表达载体”(或简称“表达载体”)。一般来说,用于重组dna技术中的表达载体通常是质粒形式。在本说明书中,由于质粒是最常用的载体形式,“质粒”和“载体”可以互换使用。然而,本发明是要包括发挥等同功能的表达载体的这样其他形式,如病毒载体(如复制缺陷型逆转录病毒,腺病毒和腺相关病毒)。
[0101]
本发明中所用术语“重组宿主细胞”(或简称“宿主细胞”)意指表达载体已经被引入其中的细胞。应当理解,这样的术语意不仅指特定对象细胞,也指这样细胞的子代。由于突变或环境影响在后代中可能出现某些修饰,因此这样的子代实际上可能不会与亲本细胞相同,但仍然包括在本发明中所用术语“宿主细胞”的范围内。重组宿主细胞包括,例如,转染细胞,如cho细胞,hek-293细胞,per.c6,ns0细胞,和淋巴细胞,以及原核细胞如大肠杆菌,以及其它真核宿主如植物细胞和真菌。
[0102]
本发明中所用术语“转染细胞”包括表达抗体或靶标抗原的重组宿主细胞,如cho细胞,per.c6,ns0细胞,hek-293细胞,植物细胞或真菌,包括酵母细胞。
[0103]
术语“制剂”是指抗体变体的制剂和不同抗体变体的混合物,当与关联细胞(例如细胞表面上表达的抗原),细胞膜,病毒粒子或其他结构接触的抗原相互作用时,其可具有提高的形成寡聚体的能力,由此能得到提高的c1q结合,补体激活,cdc,adcc,adcp,其他fc介导的效应子作用,内化,下调,细胞凋亡,抗体-药物缀合物(adc)摄取,亲和力或其任意组合。示例性的检测提供在实施例中,例如,c1q结合亲和力(实施例4),cdc(实施例5,6和10,16,19,22,23,24,25);adcc(实施例12)和体内效力(实施例20,21)。根据本发明被称为“单突变体”,“双突变体”,和“混合突变体”方面的突变体,与其示例性制备过程和使用方法在下面更详细的描述。
[0104]
本发明中所用的术语“亲和力(affinity)”是一个分子,例如抗体,与另一个,例如靶标或抗原,在单个位点,如抗体与抗原的个体抗原结合位点的单价结合,的结合强度。
[0105]
本发明中所用的术语“亲和力(avidity)”是两个结构之间,如同时与靶标相互作用的抗体多个抗原结合位点之间,或例如抗体与c1q之间,的多个结合位点的结合强度。当存在多于一种结合相互作用时,仅当所有结合位点解离时两个结构才会解离,并因此,解离速率将会低于单个结合位点,并由此相比单个结合位点结合的强度(亲和力(affinity))提供了更有效的总结合强度(亲和力(avidity))。
[0106]
本发明中所用的术语“寡聚体”是指,相比至少原则上是由不限数目的单体组成的聚合物,由多于一个但有限数目的单体单元(如抗体)组成的分子。示例性的寡聚体是二聚体,三聚体,四聚体,五聚体和六聚体。希腊语前缀经常用于表示寡聚体中的单体单元数目,例如由四个单元组成的四聚体和由六个单元组成的六聚体。
[0107]
本发明中所用的术语“寡聚化”意指将单体转化为有限程度的聚合的过程。本发明中,观察到fc域的寡聚发生是在含fc域的多肽,如抗体,优选但不限于在细胞表面的,靶标结合之后。抗体的寡聚可以,例如使用细胞表面c1q结合检测(如实施例4和9中所述),c1q效
力检测(如实施例5中所述),和实施例6,10和19中所述的补体依赖细胞毒性来评价。
[0108]
本发明中所用的术语“c1q结合”意指在c1q结合的上下文中,c1q结合到连接了其抗原的抗体上。连接到其抗原的抗体应理解为在本发明所描述的上下文中在体内和体外发生。c1q结合可以,例如通过使用人造表面上固定的抗体(例如,如实施例3所描述的elisa用塑料板),通过使用到细胞或病毒表面上预定抗原的连接(如实施例4和9中所述)来评价。c1q与抗体寡聚体的结合应理解为导致高亲和力结合的多价相互作用。
[0109]
本发明中所用的术语“补体激活”是指经典补体途径的激活,其是被补体组分c1q与连接了其抗原的抗体的结合所激活。c1q是经典补体级联早期事件中的第一个蛋白,经典补体级联包含最终导致称为c3转化酶的酶活性形成的一系列裂解反应,c3转化酶将补体组分c3裂解为c3b和c3a。c3b与膜上的c5共价结合以形成c5b,其顺次激活补体激活的晚期事件,其中末期补体组分c5b,c6,c7,c8和c9组装成膜攻击复合物(mac)。补体级联导致孔的形成,由于孔的形成引起细胞裂解,也被称为cdc。补体激活可以通过使用c1q效力(如实施例5中所述),cdc动力学(如实施例28,29和30中所述),cdc检测(如实施例6,10,19,25,27和33中所述),或者通过beurskens等2012年4月1日vol.188no.7 3532-3541中描述的c3b和c4b细胞沉积方法来评价。
[0110]
本发明中所用的术语“补体依赖性细胞毒性”(“cdc”)意指抗体介导的补体激活过程,由于mac组装所产生的膜上的孔,该过程导致抗体与细胞或病毒粒子上的其抗原连接后细胞或病毒粒子的裂解。cdc可以通过体外检测,如实施例6,10,19,25,27和33所述的其中正常人血清被用作补体源的cdc检测,或者实施例5所述的其中正常人血清已被限制在c1q中的c1q效力检测来评价。
[0111]
本发明中所用的术语“抗体依赖性细胞介导细胞毒性”(“adcc”)意指通过表达能识别所连接抗体恒定区的fc受体的细胞,来进行的抗体包被的靶细胞或病毒粒子杀伤机制。adcc可以使用方法,如实施例12中描述的adcc检测来确定。
[0112]
本发明中所用的术语“抗体依赖性细胞吞噬”(“adcp”)意指通过吞噬细胞内化来进行的抗体包被的靶细胞或病毒粒子消除机制。内化的抗体包被的靶细胞或病毒粒子被包含在称为吞噬体的囊泡中,其随后与一个或多个溶酶体融合,以形成吞噬溶酶体。adcp可以通过使用van bij等journal of hepatology volume 53,issue 4,2010年10月,677-685页中描述的,使用巨噬细胞作为效应细胞和视觉显微术的体外细胞毒性检测,或者实施例14中所述的例如pmn金黄色葡萄球菌吞噬来评价。
[0113]
本发明中所用的术语“补体依赖性细胞细胞毒性”(“cdcc”)意指通过表达能识别补体3(c3)裂解产物的补体受体的细胞,来进行的靶细胞或病毒粒子杀伤机制,其中作为抗体介导补体激活的结果,补体3(c3)裂解产物共价连接到靶细胞或病毒粒子上。cdcc可以用与对adcc所描述的类似的方式来评价。
[0114]
本发明中所用的术语“下调”意指例如通过抗体与受体的结合,来减少分子,如细胞表面的抗原或受体数目的过程。
[0115]
本发明中所用的术语“内化”意指抗体或含fc的多肽被从细胞表面和/或从周围介质中,例如通过内吞,内化入靶标表达细胞中的任何机制。抗体的内化可以使用测量内化抗体量的直接检测法(如,例如实施例26中所述的溶酶体共定位检测)来评价。
[0116]
本发明中所用的术语“抗体-药物缀合物”是指具有对至少一类型恶性细胞特异性
的抗体或含fc的多肽,药物,和将药物连接到例如抗体上的连接物。在恶性细胞存在时连接物是可裂解或不可裂解的;其中抗体-药物缀合物杀伤恶性细胞。
[0117]
本发明中所用的术语“抗体-药物缀合物摄取”是指抗体-药物缀合物被连接到细胞的靶标上,后被细胞膜摄取/吞入,并由此吸入细胞的过程。如wo 2011/157741所述抗体-药物缀合物摄取可以被评价为“抗体介导的内化和体外杀伤检测中抗tf adc的细胞杀伤”。
[0118]
本发明中所用的术语“细胞凋亡”是指可以发生在细胞中的程序性细胞死亡(pcd)过程。生物化学事件导致特征性细胞改变(形态学)和死亡,这些改变包括空泡化,细胞皱缩,核碎裂,染色质浓缩,和染色体dna片段化。抗体结合到某些受体可以诱导凋亡。
[0119]
fc受体结合可以如实施例12所述间接测量。
[0120]
本发明中所用的术语“fcrn”意指为fc受体的新生fc受体。其首先在啮齿类动物中被发现,为能够将igg从母乳中通过新生啮齿动物肠道上皮细胞输送到新生动物血流中的独特受体。进一步的研究揭示了了人中的类似受体。然而,在人中,其被发现在胎盘中以帮助促进母体igg向生长的胎儿中输送,其也已经显示在检测igg周转中发挥作用。fcrn在6.0-6.5的ph,但不在中性或更高的ph结合igg。因此,fcrn可以从微酸性ph的肠腔(肠的内部)中结合igg,并确保向ph中性到碱性(ph7.0-7.5)的基底侧有效单向输送。该受体在igg成人救助中也通过其在内皮细胞内吞途径中的发生起作用。酸性内涵体中的fcrn受体结合通过胞饮作用内化的igg,将其再循环到细胞表面,将其释放到碱性ph的血液中,由此防止其经历溶酶体降解。该机制也对血液中相比其他同种型较高的igg半衰期提供了解释。实施例13描述了显示elisa中在ph 6.0igg结合到fcrn的检测。
[0121]
本发明中所用的术语“蛋白a”意指最初在细菌金黄色葡萄球菌(staphylococcus aureus)的细胞壁上发现的56kda mscramm表面蛋白。其由spa基因编码,而其调节受dna拓扑结构,细胞渗透压,和称为arls-arlr的双组分系统控制。由于其结合免疫球蛋白的能力,也已发现了其在生物化学研究中的用途。其由折叠入三螺旋束中的5个同源ig结合域组成。每个域能结合来自许多哺乳动物种类的蛋白,最显著的是iggs。其结合大多数免疫球蛋白的重链fc区(重叠fcrn受体的保守结合位点),并也与人vh3家族的fab区域相互作用。通过血清中的这些相互作用,igg分子通过其fc区而不是单纯通过其fab区域结合细菌,由此细菌破坏调理,补体激活和吞噬。
[0122]
本发明中所用的术语“蛋白g”意指在组c和g链球菌细菌中表达的免疫球蛋白结合蛋白,其很类似蛋白a但有不同的特异性。其是65kda(g148蛋白g)和58kda(c40蛋白g)细胞表面蛋白,并已经发现其通过其结合到fc区在纯化抗体中的应用。
[0123]
本发明中所用的术语“ch2区”或“ch2域”意指免疫球蛋白的ch2区。因此例如人igg1抗体的ch2区对应根据eu编码系统的氨基酸228-340。
[0124]
本发明中所用的术语“ch3区”或“ch3域”意指免疫球蛋白的ch3区。因此例如人igg1抗体的ch2区对应根据eu编码系统的氨基酸341-447。
[0125]
本发明中所用的术语“变构突变”意思是指在按kabat中所列eu索引编号,含fc域的多肽中氨基酸p247,和e430的修饰,例如插入,取代和缺失。
[0126]
本发明中所用的术语“疏水钮突变”意指在按kabat中所列eu索引编号,含fc域的多肽中氨基酸i253,和s254,和q311的修饰,例如插入,取代和缺失。疏水钮由delano wl,等,science 287,(2000),1279-1283页,如1281页描述。
[0127]
本发明中所用的术语“n-末端ch3螺旋突变”意指按kabat中所列eu索引编号,含fc域的多肽中氨基酸r355,和d356,和e356,和e357,和m358,和l358,以及t359,更具体的d356,和e356以及t359的修饰,例如插入、取代和缺失。
[0128]
本发明中所用的术语“c-末端ch3β链突变”意指按kabat中所列eu索引编号,含fc域的多肽中氨基酸y436,和t437,和q438,和k439,和s440以及l441,更具体的y436和k439,以及s440的修饰,例如插入、取代和缺失。
[0129]
影响抗体效应子作用的方法
[0130]
应当理解本发明中描述的关于亲本抗体,第一亲本抗体或第二亲本抗体的所有实施方案也应当理解为关于包含免疫球蛋白fc域和结合区的亲本,第一亲本或第二多肽的实施方案。
[0131]
一方面本发明涉及提高亲本多肽的效应子作用的方法,所述亲本多肽包含免疫球蛋白fc域和结合域,该方法包括向亲本多肽中选自对应于人类igg1重链fc区中e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447的那些氨基酸残基的至少一个氨基酸残基中引入突变,条件是s440上的突变为s440y或s440w。
[0132]
在一个实施方案中亲本多肽可以是抗体。
[0133]
因此,本发明涉及提高亲本抗体的效应子作用的方法,包括向亲本抗体中的选自对应于人类igg1重链fc区中e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447的那些氨基酸残基中的至少一个氨基酸残基中引入突变,条件是s440上的突变为s440y或s440w。
[0134]
提及的“d/e356”在本发明上下文中是指人igg1序列中的同种异型变体。在人igg1的igg1m(za)同种异型中,位置356的氨基酸是d,而在人igg1的igg1m(f)同种异型中,位置356的氨基酸是e。
[0135]
根据本发明的方法或用途向亲本抗体中引入突变得到了变体或变体抗体。因此可以执行本发明的方法以获得本发明描述的任何变体或变体抗体。
[0136]
从本发明的方法或用途获得的变体抗体相比亲本抗体具有提高的效应子作用。通常,抗体对效应子作用的效果可以通过ec50值确定,ec50值是获得最大裂解半数值所需的抗体浓度。
[0137]
最大裂解是使用饱和量抗体时所获得的裂解,其中饱和意思是指抗体的所有抗原都被抗体连接时的抗体量。
[0138]
术语“提高效应子作用”或“改进效应子作用”本发明上下文中指相比亲本抗体,变体抗体的ec50值有降低。ec50值的降低可以是例如至多或约1/2,如至多或约1/3,或至多或约1/5,或至多或约1/10。可选的,“提高效应子作用”或“改进效应子作用”意为在亲本抗体裂解少于所有细胞100%的条件下,裂解的细胞最大量增加(其中细胞总量被设为100%)例如从所有细胞的10%到100%,如约10%,约20%,约30%,约40%,约50%,约60%,约70%,约80%,约90%,和约100%。
[0139]
可通过将igg1-005或igg1-7d8重链可变域克隆入变体并在cdc检测中测试其效力,来检测变体提高或改进效应子作用,如对daudi(实施例6)和wien(实施例10)所描述的。使用igg1-7d8 hc可变域和daudi细胞,提高被限定为与在研究条件下igg1-7d8的ec50相比小于1/2的ec50,如约1/2,约1/3,约1/5,约1/10或小于1/10的ec50值,ec50为观察到最大裂
解半数的浓度。使用igg1-005 hc可变域和daudi细胞,提高被限定为与在研究条件下igg-005的ec50相比小于1/2的ec50,如约1/2,约1/3,约1/5,约1/10或小于1/10的ec50值,ec50为观察到最大裂解半数的浓度。使用igg1-7d8 hc可变域和wien133细胞,提高被限定为与在研究条件下igg1-7d8的ec50相比小于1/2的ec50,如约1/2,约1/3,约1/5,约1/10或小于1/10的ec50值,ec50为观察到最大裂解半数的浓度。使用igg1-005 hc可变域和wien133细胞,提高被限定为最大裂解的提高从所有细胞的10%到100%,如提高约10%,约20%,约30%,约40%,约50%,约60%,约70%,约80%,约90%,和约100%。cdc效力的提高也可以被限定为与在研究条件下igg-005的ec50相比,小于1/2的ec50,如约1/2,约1/3,约1/5,约1/10或小于1/10的ec50值,ec50为在wien133细胞的裂解可以检测到的条件下,观察到最大裂解半数的浓度。
[0140]
本发明的发明人惊奇地发现,在这些特定位置的突变具有对变体抗体效应子作用的改进效果,所述抗体变体是根据本发明的方法向亲本抗体能够引入突变而获得(例如实施例19中所示)。不限于理论,认为从上述位置基团取代至少一个氨基酸会激发寡聚化。抗体结合有更高的亲和力(实施例2举例说明;igg-7d8-e345r直接标记得到相比igg-7d8-wt提高的与daudi细胞的结合),这使得抗体与细胞结合更长的时间并由此能有不同的效应子作用,例如提高的c1q结合,c1q效力cdc,adcc,内化,adcp,和/或体内效力。这些效果已经被实施例4(c1q结合在细胞上),实施例5(cdc检测中的c1q效力),实施例6,7,27,28和29(cdc检测),实施例12(adcc),实施例26(内化),以及实施例21和22(体内效力)来举例说明。
[0141]
因此选自对应于人类igg1重链fc区中e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447的那些氨基酸残基中的氨基酸残基中的突变在本发明上下文中也可以被称为“单突变”方面或“效应增强突变”。
[0142]
在另一方面,本发明也提供了表1中的一种或多种突变,如选自对应于人类igg1重链fc区中e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447的那些氨基酸残基的氨基酸残基中的突变的用途,当与在细胞,细胞膜,病毒粒子,或其他颗粒上的其抗原结合时,其能提高效应子作用,例如(i)c1q结合,(ii)补体激活,(iii)cdc,(iv)寡聚体形成,(v)寡聚体稳定性,(vi)抗体依赖性细胞介导细胞毒性(adcc),(vii)fcrn结合,(viii)fc-γ受体结合,(ix)蛋白a结合,(x)蛋白g结合,(xi)抗体依赖性细胞吞噬(adcp),(xii)补体依赖的细胞细胞毒性(cdcc),(xiii)补体增强细胞毒性,(xiv)抗体介导的与受调理抗体的补体受体结合,(xv)内化,(xvi)下调,(xvii)诱导细胞凋亡,(xviii)调理以及(xix)(i)至(xviii)中任意的组合中的一个或多个。在(iv)或(v)的一个实施方案中,寡聚体为六聚体。在一个实施方案中,抗体的至少一个其他效应子作用,如c1q结合,补体激活,补体依赖性细胞毒性(cdc),抗体依赖性细胞介导细胞毒性(adcc),fcrn结合,fc-γ受体结合,蛋白a结合,蛋白g结合,adcp,补体依赖的细胞细胞毒性(cdcc),补体增强细胞毒性,抗体介导的与受调理抗体的补体受体结合,抗体介导的细胞吞噬(adcp),内化,细胞凋亡,和/或与受调理抗体的补体受体结合也或者任选地提高,如具体为fcrn结合,adcc,fc-γ受体结合,蛋白a结合,蛋白g结合,adcp,cdcc,补体增强细胞毒性,调理及其任意组合。
[0143]
在一个实施方案中,当亲本抗体与抗原表达细胞、细胞膜或病毒粒子上的其抗原结合时,亲本抗体的效应子作用被提高。
[0144]
本发明的发明人也已显示了向亲本抗体对应人igg1重链fc区中k439或s440的氨基酸残基处引入突变降低了亲本抗体的效应子作用(实施例5,6和10)。
[0145]
本发明的另一个方面涉及降低亲本多肽的效应子作用的方法,所述亲本多肽包含免疫球蛋白fc结构域和结合域,该方法包括向亲本多肽中的选自对应于人类igg1重链fc区的k439和s440的那些氨基酸突变中的至少一个氨基酸残基中引入突变,条件是s440上的突变不是s440y或s440w,如,其中对应于人类igg1重链fc区中k439的位置中的突变是k439d/e,和/或对应于人类igg1重链fc区中s440的位置中的突变是s440k/h/r。
[0146]
在一个实施方案中亲本多肽可以是抗体。
[0147]
因此,在另一方面,本发明涉及涉及降低亲本抗体的效应子作用的方法,包括向亲本抗体中的选自对应于人类igg1重链fc区的k439和s440的那些氨基酸突变中的至少一个氨基酸残基中引入突变,条件是s440上的突变不是s440y或s440w,如其中对应于人类igg1重链fc区中k439的位置中的突变是k439d/e,和/或对应于人类igg1重链fc区中s440的位置中的突变是s440k/h/r。
[0148]
如实施例6中所示,作为“单突变体”的位置k439e或s440k的氨基酸取代,相比根据本发明方法的第一突变中任一个,降低了cdc。
[0149]
从所述降低效应子作用的方法获得的变体抗体相比亲本抗体具有降低的效应子作用。通常,抗体对效应子作用的效果可以通过ec50值测量,ec50值是获得最大裂解半数值所需的抗体浓度。
[0150]
最大裂解是使用饱和量抗体时所获得的裂解,其中饱和意指抗体的所有抗原都被抗体连接时的抗体量。
[0151]
术语“降低效应子作用”本发明上下文中指相比亲本抗体,变体抗体的ec50值有提高。ec50值的提高可以是例如至少或约2倍,如至少或约3倍,至少或约5倍,至少或约10倍。可选的,“降低效应子作用”意为在亲本抗体裂解少于所有细胞100%的条件下,裂解的细胞最大量有降低,降低例如从所有细胞的10%到100%,如约10%,约20%,约30%,约40%,约50%,约60%,约70%,约80%,约90%,和约100%。
[0152]
可以通过将igg1-005或igg1-7d8重链可变域克隆入变体并在cdc检测中测试其效力,来检测变体降低效应子作用,如对daudi(实施例6)和wien(实施例10)所描述的。使用igg1-7d8 hc可变域和daudi细胞,降低被定义为与在研究条件下igg1-7d8的ec50相比小于1/2的ec50,如约1/2,约1/3,约1/5,约1/10或小于1/10的ec50值,ec50为观察到最大裂解半数的浓度。使用igg1-005 hc可变域和daudi细胞,降低被定义为与在研究条件下igg-005的ec50相比小于1/2的ec50,如约1/2,约1/3,约1/5,约1/10或小于1/10的ec50值,ec50为观察到最大裂解半数的浓度。使用igg1-7d8 hc可变域和wien133细胞,降低被定义为与在研究条件下igg1-7d8的ec50相比小于1/2的ec50,如约1/2,约1/3,约1/5,约1/10或小于1/10的ec50值,ec50为观察到最大裂解半数的浓度。使用igg1-005 hc可变域和wien133细胞,降低被定义为最大裂解的降低从所有细胞的10%到100%,如降低约10%,约20%,约30%,约40%,约50%,约60%,约70%,约80%,约90%,和约100%。cdc效力的降低也可以被定义为与在研究条件下igg-005的ec50相比小于1/2的ec50,如约1/2,约1/3,约1/5,约1/10或小于1/10的ec50值,ec50为在wien133细胞的裂解可以检测到的条件下,观察到最大裂解半数的浓度。
[0153]
在一个实施方案中,当亲本抗体与抗原表达细胞、细胞膜或病毒粒子上的其抗原结合时,效应子作用被降低。
[0154]
因此,在另一个方面,本发明涉及抗体变体中至少更多突变的用途,所述抗体变体在选自对应于人类igg1重链fc区中的k439和s440的那些氨基酸残基中的一个氨基酸残基中包含突变,条件是s440上的突变不是s440y或s440w,当与抗原表达细胞、细胞膜或病毒粒子上的其抗原结合时,其能恢复抗体变体的效应子作用。其中
[0155]
第一突变在对应于人类igg1重链fc区中的k439的氨基酸残基处,而第二突变在对应于人类igg1重链fc区中的s440的氨基酸残基处,或者
[0156]
第一突变在对应于人类igg1重链fc区中的s440的氨基酸残基处,而第二突变在对应于人类igg1重链fc区中的k439的氨基酸残基处。
[0157]
在一个实施方案中,亲本抗体是单特异性,双特异性,或多特异性抗体。
[0158]
如果亲本抗体是包含ch2-ch3区的单特异性抗体,尽管为了大多数实用目的,根据本发明的提高或降低效应子作用的突变存在于两个ch2-ch3区中,根据本发明的突变原则上可以仅存在于ch2-ch3区的一个之中。
[0159]
如果亲本抗体是包含ch2-ch3区的双特异性抗体,尽管为了大多数实用目的,根据本发明的提高或降低效应子作用的突变存在于双特异性抗体的第一和第二ch2-ch3区中,根据本发明的突变原则上可以仅存在于ch2-ch3区的一个之中;即,在第一或第二ch2-ch3区中。
[0160]
单特异性,双特异性,或多特异性抗体的适合的例子包括任何本发明中描述的那些抗体。
[0161]
在特定的实施方案中,亲本或第一和/或第二抗体可以是双特异性抗体,如wo11/1311746中描述的异二聚体蛋白,在此其被以引用的方式纳入本发明。
[0162]
在一个实施方案中,亲本抗体是双特异性抗体,其包括了含免疫球蛋白的第一ch2-ch3区和第一抗原结合区的第一多肽,以及含免疫球蛋白的第二ch2-ch3区和第二抗原结合区的第二多肽,其中第一和第二抗原结合区结合相同抗原或不同抗原上的不同表位。
[0163]
在进一步的实施方案中所述第一ch2-ch3区在选自对应于人igg1重链fc区k409,t366,l368,k370,d399,f405,和y407的那些位置中的位置包含更多的氨基酸取代;而其中所述第一ch2-ch3区在选自对应于人类igg1重链fc区f405,t366,l368,k370,d399,y407,和k409的那些位置中的位置包含更多的氨基酸取代,而其中在第一ch2-ch3区的所述更多的氨基酸取代与第二ch2-ch3区的所述更多的氨基酸取代不同。
[0164]
在进一步的实施方案中,所述第一ch2-ch3区在对应于人igg1重链fc区k409的位置包含氨基酸取代;而所述第二ch2-ch3区在对应于人igg1重链fc区f405的位置包含氨基酸取代。
[0165]
在一个实施方案中,所述方法包括将选自对应于人igg1重链fc区中的e345,e430,s440,q386,p247,i253,s254,q311,d/e356,t359,e382,y436和k447的那些氨基酸残基中的至少一个氨基酸残基处的突变,引入到第一和第二ch2-ch3区的每一个,条件是s440上的突变是s440y或s440w。
[0166]
在进一步的实施方案中,选自对应于人igg1重链fc区中的e345,e430,s440,q386,p247,i253,s254,q311,d/e356,t359,e382,y436和k447的那些氨基酸残基中的至少一个氨
基酸残基处导入第一和第二ch2-ch3区的突变,条件是s440上的突变是s440y或s440w,可以在相同的氨基酸残基位置或不同的位置。在进一步的实施方案中其可以是相同氨基酸残基位置的相同或不同的突变。
[0167]
在另一个实施方案中,所述方法包括将选自对应于人igg1重链fc区中的e345,e430,s440,q386,p247,i253,s254,q311,d/e356,t359,e382,y436和k447的那些氨基酸残基中的至少一个氨基酸残基处的突变,引入到第一或第二ch2-ch3区,条件是s440上的突变是s440y或s440w。
[0168]
表1中所列的任何突变可以被引入双特异性抗体。实施例24显示了将e345r突变导入双特异性cd20xegfr抗体增强了cdc效力。实施例23、29和30也描述了包含根据本发明突变的一些不同的双特异性抗体。
[0169]
在一个实施方案中所述方法包括在除s440和k447之外的一个或多个位置引入突变,并进一步在如下位置引入突变
[0170]
(i)在对应于人igg1重链fc区中的k439和s440的氨基酸残基的每一个上,条件是s440上的突变不是s440y或s440w,
[0171]
(ii)在对应于人igg1重链fc区中的k447和448的氨基酸残基的每一个上,如在人igg1重链fc区上的k447k/r/h和448e/d,优选在人igg1重链fc区上的k447k和448e,或者,
[0172]
(iii)在对应于人igg1重链fc区中的k447、448和449的氨基酸残基的每一个上,如在人igg1重链fc区上的k447d/e、448k/r/h和449p,优选在人igg1重链fc区上的k447e、448k和449p。
[0173]
在一个实施方案中,所述方法包括在非s440的一个或多个位置引入突变,并在对应于人igg1重链fc区中的k439和s440的氨基酸残基的每一个上进一步引入突变,条件是s440上的突变不是s440y或s440w。
[0174]
在亲本抗体对应于人igg1重链fc区中的k439和s440的两个氨基酸残基中引入突变,条件是s440上的突变不是s440y或s440w,在本发明中也被称为“双突变体”方面。如别处所述,s440y或s440w突变已经被发现当引入亲本抗体时能提高效应子作用。
[0175]
其他地方也描述了本发明发明人已经发现在对应于人igg1重链fc区中的k439或s440的氨基酸残基上引入所鉴定的突变,导致效应子作用的降低(实施例5、6、10)。然而,当在对应于人igg1重链fc区中的k439和s440的两个氨基酸残基上引入抑制突变时,效应子作用的降低被恢复,由此使其与在k439和s440突变处没有突变的亲本抗体的效应子作用相似。然而,不限于任何理论,认为k439和s440突变的存在限制了效应子作用诱导为寡聚复合物,所述寡聚复合物仅由单独包含k439和s440两个突变的抗体组成。因此,如果治疗抗体中包括k439和s440突变,不限于任何理论,认为当这样的治疗抗体被施用给患者时,效应子作用诱导被限制为寡聚抗体复合物,所述寡聚抗体复合物包含含有k439/s440突变的治疗抗体但不含患者自身抗体,患者自身抗体不含k439和s440突变,由此限制了由治疗抗体与患者自身抗体相互作用引起的任何潜在副作用。
[0176]
当将位置k439和/或s440的突变与第一突变组合时,获得了cdc的增强并提高了cdc的特异性。
[0177]
因此在另一个方面,本发明涉及提高包含免疫球蛋白fc域和结合区的至少第一和第二亲本多肽的组合的特异性的方法,包括:
[0178]
a)
[0179]
(i)向第一亲本多肽对应于人igg1重链fc区中的k439位置的氨基酸残基中引入突变;和
[0180]
(ii)向第二亲本多肽对应于人igg1重链fc区中的s440位置的氨基酸残基中引入突变,条件是s440上的突变不是s440y或s440w,
[0181]
b)
[0182]
(i)向第一亲本多肽对应于人igg1重链fc区中的k447d/e位置的氨基酸残基中引入突变;和
[0183]
(ii)向第二亲本多肽对应于人igg1重链fc区中的k447k/r/h和448p位置的氨基酸残基中引入突变;或者
[0184]
c)
[0185]
(i)向第一亲本多肽对应于人igg1重链fc区中的k447d/e位置的氨基酸残基中引入突变;和
[0186]
(ii)向第二亲本多肽对应于人igg1重链fc区中的k447k/r/h,448k/r/h和449p位置的氨基酸残基中引入突变。
[0187]
在一个实施方案中,亲本多肽,第一亲本多肽和第二亲本多肽可以各为抗体。
[0188]
因此,在进一步的方面,本发明也涉及提高至少第一和第二亲本抗体的组合的特异性的方法,包括
[0189]
(i)向第一亲本抗体对应于人igg1重链fc区中的k439位置的氨基酸残基中引入突变;和
[0190]
(ii)向第二亲本抗体对应于人igg1重链fc区中的s440位置的氨基酸残基中引入突变,条件是s440上的突变不是s440y或s440w。
[0191]
如实施例10中所示,相比任何野生型或天然存在的抗体,第一和第二变体抗体会有相互寡聚的偏好。
[0192]
特异性的提高是关于“效应子作用诱导”的。因此在一个实施方案中所述方法是通过至少第一和第二亲本抗体的组合来提高效应子作用的诱导特异性的方法。
[0193]
进行通过至少第一和第二亲本抗体的组合来提高特异性,或效应子作用的诱导特异性的方法,获得了至少第一突变和第二突变抗体的组合。
[0194]
通过在亲本抗体k439或s440上引入突变,由此获得的变体抗体相比亲本抗体具有降低的效应子作用。然而,本发明其他地方也描述了k439和s440处的突变能相互补偿以恢复包含两个突变的抗体的效应子作用。k439和s440处突变相互补偿的能力可以被类似的用于两个抗体中。因此,当k439处的突变被引入第一亲本抗体而s440处的突变被引入第二亲本抗体时,或者相反,第一和第二突变抗体组合使用就不再会看到效应子作用的降低。术语“提高特异性”或“改进特异性”在本上下文中是指由包含k439处突变的第一变体抗体和包含s440处突变的第二变体抗体的组合所诱导的效应应答,高于由包含k439处突变的第一变体抗体或包含s440处突变的第二变体抗体所诱导的效应应答。
[0195]
通过k439和s440处两个氨基酸取代的引入,使寡聚体的特异性提高。
[0196]
当将位置k439和/或s440的突变与第一突变组合时,获得了cdc的增强并提高了cdc的特异性。
[0197]
在一个实施方案中,至少第一和第二亲本抗体结合到相同的表位。
[0198]
在一个实施方案中,至少第一和第二亲本抗体结合到相同抗原上的不同表位。
[0199]
在一个实施方案中,至少第一和第二亲本抗体结合到不同靶标上的不同表位。
[0200]
在一个实施方案中,第一和第二亲本抗体具有相同或不同的vl和vh序列。
[0201]
在一个实施方案中,至少第一和第二亲本抗体的组合包括一种第一亲本抗体和一种第二亲本抗体。
[0202]
在一个实施方案中,第一和第二亲本抗体的组合连接到抗原表达细胞、细胞膜或病毒粒子上的其抗原上时,特异性提高。
[0203]
因此,在另一方面,本发明也涉及抗体两个或更多个氨基酸残基处突变的用途,当抗体连接到抗原表达细胞、细胞膜或病毒粒子上的其抗原上时,所述突变增加例如抗体诱导的效应子作用的特异性,其中,
[0204]
第一突变在对应于人igg1重链fc区中的k439的氨基酸残基处;
[0205]
第二突变在对应于人igg1重链fc区中的s440的氨基酸残基处。
[0206]
在进一步的方面,本发明涉及提高至少第一和第二亲本多肽组合的效应子作用的方法,其中至少第一和第二亲本多肽各包含免疫球蛋白fc域和结合区,其中所述方法包括
[0207]
(i)向至少第一和/或第二亲本多肽的选自下组的一个或多个氨基酸残基处引入突变:
[0208]
(a)提供变构突变ch2-ch3区内的氨基酸残基,
[0209]
(b)ch2-ch3区的疏水钮内的氨基酸残基,
[0210]
(c)n-末端ch3螺旋内的氨基酸残基,
[0211]
(d)c-末端ch3β链内的氨基酸残基,条件是在对应人igg1重链fc区中s440的突变的情况下,突变为s440y或s440w,和
[0212]
(e)对应人igg1重链fc区中e345、e382或q386的氨基酸残基。
[0213]
在一个实施方案中,第一和/或第二亲本多肽可以各为抗体。
[0214]
因此在一个实施方案中,本发明涉及提高至少第一和第二亲本抗体的组合的效应子作用的方法,其中至少第一和第二亲本抗体各包含免疫球蛋白fc域和抗原结合区,其中所述方法包括
[0215]
(i)向至少第一和/或第二亲本抗体的选自下组的一个或多个氨基酸残基处引入突变:
[0216]
(a)提供变构突变ch2-ch3区内的氨基酸残基,
[0217]
(b)ch2-ch3区的疏水钮内的氨基酸残基,
[0218]
(c)n-末端ch3螺旋内的氨基酸残基,
[0219]
(d)c-末端ch3β链内的氨基酸残基,条件是在对应人igg1重链fc区中s440的突变的情况下,突变为s440y或s440w,和
[0220]
(e)对应人igg1重链fc区中e345、e382或q386的氨基酸残基。
[0221]
通过进行该方法,获得了至少第一和第二变体抗体的组合。通过该方法获得的至少第一和第二变体抗体当组合时具有相比第一和第二亲本抗体的组合提高的效应子作用。
[0222]
术语“提高的效应子作用”应按本发明中所描述的理解。
[0223]
第一和/或第二亲本抗体可以是本发明所描述的任何亲本抗体。
[0224]
具体地,可以进行提高第一和第二抗体组合的效应子作用的方法,以获得具有本发明所描述变体抗体任何特征的第一和第二抗体。本发明发明人已发现将突变引入选自(a)、(b)、(c)、(d)和/或(e)的氨基酸残基得到的第一和第二变体抗体的组合,相比第一和第二亲本抗体的组合具有提高的效应子作用。
[0225]
在一个实施方案中,至少第一和第二亲本抗体结合到相同的表位。
[0226]
在一个实施方案中,至少第一和第二亲本抗体结合到相同抗原上的不同表位。
[0227]
在一个实施方案中,至少第一和第二亲本抗体结合到不同靶标上的不同表位。
[0228]
在一个实施方案中,至少第一和第二亲本抗体具有相同或不同的vl和vh序列。
[0229]
在一个实施方案中,至少第一和第二亲本抗体的组合包括一种第一亲本抗体和一种第二亲本抗体。
[0230]
在一个实施方案中,至少第一和第二亲本抗体的组合包括更多的亲本抗体,如第三,第四或第五亲本抗体。
[0231]
在一个实施方案中,(a)提供变构突变ch2-ch3区内的氨基酸残基是选自对应于人igg1重链fc区p247或e430的那些氨基酸残基的氨基酸残基。
[0232]
在一个实施方案中,(b)ch2-ch3区的疏水钮内的氨基酸残基是选自对应于人igg1重链fc区i253、s254和q311的那些氨基酸残基的氨基酸残基。
[0233]
在一个实施方案中,(c)n-末端ch3螺旋内的氨基酸残基是选自对应于人igg1重链fc区d/e356和t359的那些氨基酸残基的氨基酸残基。
[0234]
在一个实施方案中,(d)c-末端ch3β链内的氨基酸残基是选自对应y436和s440的那些氨基酸残基的氨基酸残基。
[0235]
(b)、(c)、(d)和(e)中的氨基酸残基是为位于两抗体fc:fc界面处,因此抗体fc部分可以与另一抗体fc部分相互作用,两抗体相互接近。
[0236]
因此,在进一步的实施方案中,至少第一和/或第二亲本抗体中的突变是在选自对应于人igg1重链fc区中的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447的那些氨基酸残基中至少一个氨基酸残基处,条件是s440上的突变是s440y或s440w。
[0237]
在一个实施方案中(i)包括在第一和第二亲本抗体中引入突变。
[0238]
在另一个实施方案中所述方法包括:
[0239]
(i)向第一亲本抗体中对应于人igg1重链fc区中的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447的那些氨基酸残基中至少一个氨基酸残基处引入突变,条件是s440上的突变是s440y或s440w,
[0240]
(ii)提供第二亲本抗体,其在选自对应于人igg1重链fc区中的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436、和k447的那些氨基酸残基的氨基酸残基处不包含突变。
[0241]
在一个实施方案中,所述方法包括在除s440之外的至少一个氨基酸残基处引入突变,其中所述方法进一步包括在非s440的一个或多个位置处引入突变的步骤,而且其中所述方法进一步包括如下步骤
[0242]
(i)向第一亲本抗体对应于人igg1重链fc区中的k439位置的氨基酸残基处引入第二突变;和
[0243]
(ii)向第二亲本抗体对应于人igg1重链fc区中的s440位置的氨基酸残基处引入第二突变,条件是s440上的突变不是s440y或s440w;其中步骤(ii)和(iii)可替换地可为
[0244]
(i)向第一亲本抗体对应于人igg1重链fc区中的s440位置的氨基酸残基处引入第二突变,条件是s440上的突变不是s440y或s440w;和
[0245]
(ii)向第二亲本抗体对应于人igg1重链fc区中的k439位置的氨基酸残基处引入第二突变。
[0246]
对于本发明那些实施方案,其中第二亲本在选自对应于人igg1重链fc区中的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447的那些氨基酸残基的氨基酸残基处不包含突变,步骤(ii)中的术语“第二突变”可以是第一突变,例如第二亲本抗体仅包含步骤(ii)中引入的突变。步骤(i)和(ii)中的术语“第二突变”也不是意欲限制引入第一和/或第二亲本抗体中的突变数目。
[0247]
在一个实施方案中,亲本抗体,第一亲本抗体和第二亲本抗体可各选自包括,但不限于单特异性、双特异性和多特异性抗体的组。双特异性可以是例如异二聚体蛋白。
[0248]
在一个实施方案中,第一和第二亲本抗体是单特异性抗体,其可以例如连接相同或不同的表位。如果第一和第二亲本抗体连接不同的表位,其可以在相同或不同的抗原上。
[0249]
在另一个实施方案中,第一亲本抗体是单特异性抗体,而第二亲本抗体是双特异性或多特异性抗体,反之亦然。
[0250]
在另一个实施方案中,第一和第二亲本抗体是双特异性或多特异性抗体。在一个实施方案中,第一和第二双特异性或多特异性抗体是相同或不同的抗体。在一个实施方案中,第一和第二双特异性或多特异性抗体结合相同或不同抗原上不同的表位。因此,在一个实施方案中,至少第一和第二亲本抗体是结合相同或不同抗原上不同表位的双特异性或多特异性抗体。
[0251]
在另一个实施方案中,第一亲本抗体是单特异性抗体,而第二亲本抗体是双特异性抗体,反之亦然。单特异性可以结合相同表位,而双特异性(或部分双特异性)或单特异性和双特异性抗体可以结合相同或不同抗原上不同的表位。双特异性抗体可以结合相同或不同抗原上不同的表位。
[0252]
在一个实施方案中,所述至少第一和第二亲本抗体各为双特异性抗体,双特异性抗体包括含免疫球蛋白的第一ch2-ch3区和第一抗原结合区的第一多肽,以及含免疫球蛋白的第二ch2-ch3区和第二抗原结合区的第二多肽,其中第一和第二抗原结合区结合相同或不同抗原上的不同表位,和其中所述第一ch2-ch3区在选自对应于人igg1重链fc区k409、t366、l368、k370、d399、f405和y407的那些位置的位置处包含进一步的氨基酸取代;和其中所述第二ch2-ch3区在选自对应于人igg1重链fc区f405、t366、l368、k370、d399、y407和k409的那些位置的位置处包含进一步的氨基酸取代;而其中第一ch2-ch3区中的所述进一步的氨基酸取代与第二ch2-ch3区中的所述进一步的氨基酸取代不同。
[0253]
在进一步的实施方案中,所述第一ch2-ch3区在对应于人igg1重链fc区k409的位置处包含氨基酸取代;而所述第二ch2-ch3区在对应于人igg1重链fc区f405的位置处包含氨基酸取代。
[0254]
在本发明方法和/或用途的一个实施方案中,亲本抗体,无论其是否为亲本抗体,第一亲本抗体或第二亲本抗体,除了已经被发现能影响效应子作用的本发明那些突变之外
还包含其他突变。这样的突变可以与影响效应子作用的本发明突变同时被引入,或者其可以被顺序引入。本发明的方法或用途不限于同时或顺序引入突变。双特异性抗体可以是任何双特异性抗体,而由于预见到可以使用不同形式,本发明的方法或用途不限于任何特定的双特异性抗体形式。
[0255]
组合包含能提高效应子作用的一个所述突变的第一抗体与不包含这样突变的第二抗体的方法可以如实施例31所示增加组合的效应子作用。因此,不限于任何理论,认为例如该方法可以被用于组合已被证明安全但不足够有效的作为第二抗体的治疗抗体,与包含突变的第一抗体,并由此得到有效的组合。
[0256]
因此在一个实施方案中,在选自对应于人igg1重链fc区中的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447的那些氨基酸残基的氨基酸残基处不包含突变的第二亲本抗体是治疗抗体。在特定的实施方案中其为具有适合的安全性谱(profile)的治疗抗体。在一个实施方案中其为具有适合的安全性谱但不是足够有效的治疗抗体。
[0257]
在对应人igg1重链fc区中e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447的那些氨基酸残基的中所选氨基酸残基处不包含突变的适合的第二抗体的例子包括但不限于任何下面的:(90y)clivatuzumab tetraxetan;(90y)tacatuzumab tetraxetan;(99mtc)fanolesomab;(99mtc)nofetumomab merpentan;(99mtc)pintumomab;3f8;8h9;abagovomab;阿巴西普;阿昔单抗;actoxumab;阿达木单抗;adecatumumab;阿非莫单抗;aflibercept;afutuzumab;alacizumab pegol;albiglutide;ald518;阿法赛特;阿仑单抗;alirocumab;altumomab;altumomab pentetate;alvircept sudotox;amatuximab;amg714/humax-il15;anatumomab mafenatox;anrukinzumab(=ima-638);apolizumab;arcitumomab;aselizumab;atacicept;atinumab;atlizumab(=托珠单抗);atorolimumab;baminercept;bapineuzumab;巴利昔单抗;bavituximab;bectumomab;belatacept;belimumab;benralizumab;bertilimumab;besilesomab;贝伐单抗;bezlotoxumab;biciromab;bifarcept;bivatuzumab;bivatuzumab mertansine;blinatumomab;blosozumab;brentuximab vedotin;briakinumab;briobacept;brodalumab;canakinumab;cantuzumab mertansine;cantuzumab ravtansine;caplacizumab;capromab;capromab pendetide;carlumab;catumaxomab;cc49;cedelizumab;certolizumab pegol;西妥昔单抗;ch.14.18;citatuzumab bogatox;cixutumumab;clazakizumab;clenoliximab;clivatuzumab tetraxetan;conatumumab;conbercept;cr6261;crenezumab;dacetuzumab;达利珠单抗;dalantercept;dalotuzumab;daratumumab;demcizumab;地诺单抗;detumomab;dorlimomab aritox;drozitumab;dulaglutide;ecromeximab;依库珠单抗;edobacomab;依决洛单抗;依法利珠单抗;efungumab;elotuzumab;elsilimomab;enavatuzumab;enlimomab;enlimomab pegol;enokizumab;ensituximab;epitumomab;epitumomab cituxetan;依帕珠单抗;erlizumab;ertumaxomab;依那西普;etaracizumab;etrolizumab;exbivirumab;fanolesomab;faralimomab;farletuzumab;fasinumab;fbta05;felvizumab;fezakinumab;ficlatuzumab;figitumumab;flanvolumab;fontolizumab;foralumab;foravirumab;fresolimumab;fulranumab;galiximab;ganitumab;gantenerumab;gavilimomab;吉妥珠单
抗;吉妥珠单抗奥唑米;gevokizumab;girentuximab;glembatumumab;glembatumumab vedotin;戈利木单抗;gomiliximab;gs6624;抗-cd74抗体;如wo2011/110642公开的抗cmet抗体;如wo2011/147986或wo2011/147982公开的抗her2抗体;如wo2004/058797公开的抗il-8抗体;如wo2004/045512公开的抗tac抗体;如wo2010/066803或wo 2011/157741公开的抗组织因子(tf)抗体;ibalizumab;替伊莫单抗;icrucumab;igovomab;imciromab;inclacumab;indatuximab ravtansine;英夫利昔单抗;inolimomab;inotuzumab ozogamcin;intetumumab;碘(i241)girentuximab;匹伊单抗;iratumumab;itolizumab;ixekizumab;keliximab;labetuzumab;lebrikizumab;lemalesomab;lenercept;lerdelimumab;lexatumumab;libivirumab;林妥珠单抗;lorvotuzumab mertansine;lucatumumab;lumiliximab;mapatumumab;maslimoma;马妥珠单抗;mavrilimumab;美泊利单抗;metelimumab;milatuzumab;minretumomab;mirococept;米妥莫单抗;mogamulizumab;morolimumab;motavizumab;moxetumomab;pasudotox;莫罗单抗-cd3;nacolomab tafenatox;namilumab;naptumomab estafenatox;narnatumab;那他珠单抗;nebacumab;necitumumab;nerelimomab;尼妥珠单抗;nivolumab;nofetumomab;merpentan;obinutuzumab;ocaratuzumab;ocrelizumab;odulimomab;奥法木单抗;olaratumab;olokizumab;奥马珠单抗;onartuzumab;奥那西普;oportuzumab monatox;oregovomab;otelixizumab;oxelumab;ozoralizumab;pagibaximab;帕利珠单抗;帕尼单抗;panobacumab;pascolizumab;pateclizumab;patritumab;pegsunercept;pemtumomab;帕妥珠单抗;pexelizumab;pintumomab;placulumab;ponezumab;priliximab;pritumumab;pro 140;quilizumab;racotumomab;radretumab;rafivirumab;ramucirumab;兰尼单抗;raxibacumab;regavirumab;reslizumab;rg1507/humax-igflr;rg1512/humax-pselectin;rilonacept;rilotumumab;利妥昔单抗;robatumumab;roledumab;romosozumab;rontalizumab;rovelizumab;ruplizumab;samalizumab;sarilumab;satumomab;satumomab pendetide;secukinumab;sevirumab;sibrotuzumab;sifalimumab;siltuximab;siplizumab;sirukumab;solanezumab;solitomab;sonepcizumab;sontuzumab;sotatercept;stamulumab;sulesomab;suvizumab;tabalumab;tacatuzumab tetraxetan;tadocizumab;talizumab;tanezumab;taplitumomab paptox;tefibazumab;telimomab aritox;tenatumomab;teneliximab;teplizumab;teprotumumab;tgn1412;ticilimumab(=tremelimumab);tigatuzumab;tnx-650;托珠单抗(=atlizumab);toralizumab;torapsel;托西莫单抗;tralokinumab;曲妥珠单抗;曲妥珠单抗emtansine;trbs07;trebananib;tregalizumab;tremelimumab;tucotuzumab celmoleukin;tuvirumab;ublituximab;urelumab;urtoxazumab;ustekinumab;vapaliximab;vatelizumab;vedolizumab;veltuzumab;vepalimomab;vesencumab;visilizumab;volociximab;vorsetuzumab mafodotin;votumumab;扎妥木单抗;zanolimumab;ziralimumab;和zolimomab aritox。
[0258]
在本发明方法和用途的一个实施方案中,在对应于人igg1重链fc区中的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447的至少一个氨基酸残基,或一个或多个氨基酸残基处的突变,条件是s440上的突变是s440y或s440w,可以在表1所列任何示例性和优选的氨基酸位置。因此表1所列每个氨基酸位置是至少一个氨基酸处单独的、非限制性的突变实施方案。
[0259]
可以根据本发明的方法引入本发明所描述的任何突变或其组合。
[0260]
选自示例性或优选氨基酸取代的突变可以以适当的检测方法测试,所述检测方法允许抗原连接抗体的寡聚物形成并检测增强的c1q结合,补体激活,cdc,adcc和/或内化,如实施例中所描述的那些。例如c1q结合亲和力可以根据类似于实施例4所描述检测方法的检测方法,使用表达抗体变体的抗原的细胞来确定。示例性示例性的cdc检测方法在实施例5、6、10、16、19、22、23、24或25中提供。示例性示例性adcc检测方法在实施例12中提供。示例性示例性内化检测方法在实施例26中提供。最后,为区分直接参与c1q结合的氨基酸残基的突变与影响寡聚物形成的突变,根据例如实施例3的elisa检测中的c1q结合可以与根据例如实施例4的基于细胞检测中的c1q结合相比较。
[0261]
在进一步的实施方案中,所述突变选自对应于人igg1重链fc区中的e345、e430、s440和q386的那些突变,条件是s440上的突变是s440y或s440w。
[0262]
在可选的实施方案中,在至少一个氨基酸残基,或一个或多个氨基酸残基处的突变是在对应于人igg1重链fc区e382和h433的氨基酸残基处。
[0263]
在特定的实施方案中,一个突变在对应于人igg1重链fc区中的e345的氨基酸残基处。
[0264]
在特定的实施方案中,一个突变在对应于人igg1重链fc区e430的氨基酸残基处。
[0265]
在特定的实施方案中,一个突变在对应于人igg1重链fc区中的s440的氨基酸残基处,条件是s440上的突变是s440y或s440w。
[0266]
在特定的实施方案中,一个突变在对应于人igg1重链fc区q386的氨基酸残基处。
[0267]
在可选的实施方案中,一个突变在对应于人igg1重链fc区e382或h433的氨基酸残基处。
[0268]
在一个实施方案中,在至少一个氨基酸残基处的突变可以是氨基酸取代、氨基酸缺失或氨基酸插入。
[0269]
在一个实施方案中,在至少一个氨基酸残基处的突变可以是氨基酸缺失。
[0270]
在一个实施方案中,在至少一个氨基酸残基处的突变可以是氨基酸插入。
[0271]
在一个实施方案中,在至少一个氨基酸残基处的突变可以是氨基酸取代。
[0272]
在一个实施方案中,在至少一个氨基酸残基处的突变可以选自表1所列的任何氨基酸取代,氨基酸缺失。此外,表1所列每个特定氨基酸残基处每个优选的氨基酸取代是该用途单独和特定非限制性实施方案。示例性示例性氨基酸取代包括将e残基交换为r残基,和将h残基交换为r残基。
[0273]
在进一步的实施方案中,在至少一个氨基酸残基处的突变是选自对应于人igg1重链fc区中的e345x、e430x、s440y或w,和q386k的那些氨基酸取代的氨基酸取代,其中x指任何氨基酸,例如天然氨基酸或非天然发生的氨基酸。x可以具体的指任何20个天然发生氨基酸。
[0274]
因此,在一个实施方案中,突变在选自对应于人igg1重链fc区中的e345、e430、s440到y或w,和q386的那些氨基酸残基的至少一个氨基酸残基处,优选的其中突变是至少一个下列氨基酸取代:e345到r、q、n或k,e430到t、s或g,s440到y或w,或者q386到k。
[0275]
因此,在一个实施方案中,e345x可以是e345r、q、n、k、y、a、c、d、f、g、h、i、l、m、p、s、t、v、w或y;具体是e345a、d、g、h、k、n、q、r、s、t、y或w,或更具体是e345d、k、n、q、r或w;或甚至
更具体是e345r、q、n、k或y。在另一进一步的实施方式中e430x可以是e430t、s、g、f、h、a、c、d、i、k、l、m、n、p、q、r、v、w或y;具体是e430t、s、g、f或h。在优选的实施方案中,氨基酸取代选自包含e345r、e345q、e345n、e345k、e345y、e430t、e430s、e430g、e430f、e430h、s440w和s440y的组。在进一步的实施方案中,至少一个氨基酸残基处的突变选自e345r和e430g。在进一步的实施方案中,至少一个氨基酸残基处的突变是e345r。在进一步的实施方案中,至少一个氨基酸残基处的突变是e430g。
[0276]
在可选的实施方案中,至少一个氨基酸残基处的突变是选自对应于人igg1重链fc区i253、h310、q311、e382、g385、h433、n434、y436、q438的那些突变,如e382和h433。在进一步可选的实施方案中,至少一个氨基酸残基处的突变可以是选自i253e、n、q、s或t,例如i253n或q;h310n、q、w或y,例如h310q;q311e或r,e382d、h、k、r、n、q、s、t、w或y,例如:e382d、q、k或r;g385e、h、k、n、q、r、s、t、w或y,例如g385d、e、k或r;h433r;n434d、e、h、k、q、r、s、t、w或y,例如n434h、k、q或r;y436a、e、f、h、i、k、l、m、n、q、r、s、t或v,例如y436n、q、s或t;q438a、e、g、h、k、n、q、r、s、t、w或y,或例如q438 n、s或t的那些氨基酸取代的氨基酸取代。
[0277]
因此,在更进一步的可选实施方案中,至少一个氨基酸残基处的突变可以是选自对应于人igg1重链fc区p247g、i253v、s254l/v、q311l/w、d/e356g/r、t359r、e382l/v和y436i的那些氨基酸取代的氨基酸取代,例如具体是e382l、v、d、q、k或r或者h433r。在进一步的可选实施方案中,至少一个氨基酸残基处的突变选自e382r和h433r。在可选实施方案中,突变是e382r。在另一个可选实施方案中,突变是h433r。
[0278]
在另一个实施方案中,可选的,按通过将根据实施例3的elisa检测中c1q结合与根据实施例4的基于细胞检测中的c1q结合相比较来确定,突变不在直接参与c1q结合的氨基酸残基处。
[0279]
在一个实施方案中,突变不在对应于i253、n434或q311的氨基酸残基处,可选的不在对应h433的氨基酸残基处,或氨基酸取代不是h433a。
[0280]
在一个实施方案中,至少一个突变是一个突变,即不多于一个突变被引入亲本抗体。
[0281]
在另一个实施方案中,根据本发明的方法或用途包括在至少两个,如两个,三个,四个,五个或更多的表1氨基酸残基处引入突变。
[0282]
可以根据本发明的方法引入本发明所描述的任何突变组合。
[0283]
在一个实施方案中,根据本发明的方法或用途包括向亲本抗体中选自对应于人igg1重链fc区中的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447的那些氨基酸残基的至少两个氨基酸残基处引入突变,条件是s440上的突变是s440y或s440w。
[0284]
在一个实施方案中,根据本发明的方法或用途包括向亲本抗体中选自对应于人igg1重链fc区中的e345、e430、q386和s440的那些氨基酸残基的至少两个氨基酸残基处引入突变,条件是s440上的突变是s440y或s440w,如其中至少两个氨基酸处的突变选自下列:e345到r、q、n或k,e430到t、s或g,s440到y或w,或者q386到k。
[0285]
在可选的实施方案中,第一突变的位置可以选自由位置i253、h310、q311、e345、e382、g385、h433、n434、y436和q438组成的组。
[0286]
在一个实施方案中,方法进一步包括向抗体对应于第一和/或第二fc区中e345、
e430、p247、i253、s254、q311、d/e356、t359、e382、q386、y436或k447的氨基酸残基处引入更多或第三突变。
[0287]
例如多于一个,如两个,三个,四个,或五个,具体是两个或三个突变被引入到亲本抗体选自对应于人igg1重链fc区中的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447的那些氨基酸残基的氨基酸残基处。例如,可以突变至少一个对应于人igg1重链fc区中的e345、e430和s440的氨基酸残基,如e345、e430和s440的两个或全部,可选的与一个或多个表1所列其他氨基酸处的突变组合。至少两个突变可以是位置e345的任何氨基酸残基取代组合上位置e430或s440的任何氨基酸取代,或者可以是位置e430的任何氨基酸残基取代组合上位置s440的任何氨基酸取代。
[0288]
在进一步的实施方案中,两个或三个突变被引入到亲本抗体选自对应于人igg1重链fc区中的e345、e430、s440和q386的那些氨基酸残基的氨基酸残基处。
[0289]
在一个实施方案中,多于一个突变可以具体是氨基酸取代。
[0290]
因此,根据本发明,方法或用途包括向抗体导入选自下列由p247g、i253v、s254l、q311l/w、e345x、d/e356g/r、t359r、e382l/v、q386k、e430x、y436i和s440y/w组成的组的至少一个,如一个、两个、三个、四个、五个或六个氨基酸取代。在优选的实施方案中,氨基酸取代选自由e345x、e430x、s440y/w和q386k组成的组。
[0291]
在可选的实施方案中,至少两个,如两个、三个、四个或五个突变在选自对应于人igg1重链fc区h310、g385、h433、n434和q438的那些氨基酸残基的氨基酸残基处。
[0292]
在另一个可选实施方案中,至少一个突变,可选的两个或三个突变,选自由e345r、e382r和h433r组成的组。在另一个可选实施方案中,可以突变对应于人igg1重链fc区e382和h433的至少一个氨基酸残基,如两个,任选地与一个或多个表1所列其他氨基酸处的突变组合。
[0293]
在本发明方法和/或用途的某些实施方案中,对应于k439和/或s440的氨基酸残基中的突变被引入选自由亲本抗体,第一亲本抗体,第二亲本抗体及其组合组成的组的抗体。如上所述,显示了在对应于人igg1重链fc区中的k439和s440的氨基酸残基处引入突变,将抗体间的分子间相互作用限制到包含突变的那些(实施例4、5、6、10)。取决于k439和s440是否被引入相同的亲本抗体或者在第一个第二亲本抗体中,这些方面也被称为“双突变体”和“混合突变体”方面。
[0294]
在本发明一个实施方案中,对应于人igg1重链fc区中的k439的氨基酸残基处的突变是氨基酸取代。
[0295]
在本发明一个实施方案中,对应于人igg1重链fc区中的s440的氨基酸残基处的突变是氨基酸取代。
[0296]
在本发明所有实施方案中,其中在对应于人igg1重链fc区中的k439和s440位置的突变,无论是在相同多肽或抗体上,或者在第一和第二多肽或抗体上,可以被下述中的突变所替代:
[0297]
(i)在每个对应于人igg1重链fc区中的k447和448的氨基酸残基上,如人igg1重链fc区中的k447k/r/h和448e/d,优选igg1重链fc区中的k447k和448e,或者
[0298]
(ii)在每个对应于人igg1重链fc区中的k447、448和449的氨基酸残基上,如人igg1重链fc区中的k447d/e、k448k/r/h和449p,优选igg1重链fc区中的k447e、448k和449p。
[0299]
因此这样突变的组合包括任何表2a和2b描述的那些。
[0300]
在本发明一个实施方案中,在对应于人igg1重链fc区中的k439和s440氨基酸残基处的突变均是氨基酸取代。
[0301]
在一个实施方案中,在对应于人igg1重链fc区中的k439氨基酸残基处的突变是取代为选自e和d的氨基酸的氨基酸取代。
[0302]
在另一个实施方案中,突变是k439e。
[0303]
在一个实施方案中,在对应于人igg1重链fc区中的s440氨基酸残基处的突变是取代为选自k、r和h的氨基酸的氨基酸取代。
[0304]
在另一个实施方案中,突变是s440k。
[0305]
因此在进一步的实施方案中,被引入亲本抗体对应于人igg1重链fc区中的k439和s440的氨基酸残基处的突变是选自k439e和d,以及s440k、r和h的氨基酸取代。
[0306]
因此在进一步的实施方案中,被引入亲本抗体对应于人igg1重链fc区中的k439和s440的氨基酸残基处的突变是氨基酸取代k439e和s440k。
[0307]
本发明的某些方法和用途包括第一和第二亲本抗体。
[0308]
因此在进一步的实施方案中,被引入第一亲本抗体对应于k439氨基酸残基处的突变是选自k439e和d的氨基酸取代,例如k439e,而被引入第二亲本抗体对应于s440氨基酸残基处的突变时选自s440k、r和h的氨基酸取代,例如s440k。第一和第二亲本抗体中的突变可以被同样相反的引入,即,其也可以是对应于s440氨基酸残基处的突变被引入第一亲本多肽,而对应于k439氨基酸残基处的突变被引入第二亲本多肽,其中突变可以是上述优选的氨基酸取代。
[0309]
在根据本发明的方法或用途的一个实施方案中,当抗体被连接到其抗原上时效应子作用被提高。
[0310]
在进一步的实施方案中,当抗体被连接到其抗原上时效应子作用被提高,其中抗原是在抗原表达细胞、细胞膜或病毒粒子上。在一个实施方案中,igg1重链fc区包括seq id no:1残基130-330的序列。
[0311]
亲本抗体可以是本发明描述的任何亲本抗体。本上下文中的亲本抗体也是第一亲本和第二亲本抗体。
[0312]
在一个实施方案中,亲本抗体是人igg1、igg2、igg3或igg4、iga1、iga2、igd或ige抗体。
[0313]
在一个实施方案中,亲本抗体是人全长抗体,如人全长igg1抗体。
[0314]
在一个实施方案中,亲本抗体,第一亲本抗体和第二亲本抗体是人igg1抗体,例如igg1m(za)或igg1m(f)同种异型体,任选地包括含seq id no:1或5的fc区。
[0315]
在一个实施方案中,亲本抗体是人igg2抗体,任选地包括含seq id no:2的fc区。
[0316]
在一个实施方案中,亲本抗体是人igg3抗体,任选地包括含seq id no:3的fc区。
[0317]
在一个实施方案中,亲本抗体是人igg4抗体,任选地包括含seq id no:4的fc区。
[0318]
在一个实施方案中,亲本抗体是双特异性抗体。
[0319]
在一个实施方案中,亲本抗体是本发明描述的任何抗体,例如包含至少部分fc区的抗体片段,单价抗体(在wo2007059782中由genmab描述);重链抗体,其仅由两条重链组成并在例如骆驼中天然存在(例如,hamers-casterman(1993)nature 363:446);thiomabs
(roche,wo2011069104),链交换改造域(seed或seed体),其是不对称的和双特异性的抗体样分子(merck,wo2007110205);triomab(fresenius,lindhofer等(1995j immunol 155:219));fcδadp(regeneron,wo2010151792),azymetric scaffold(zymeworks/merck,wo2012/058768),mab-fv(xencor,wo2011/028952),双可变域免疫球蛋白(abbott,dvd-ig,美国专利7,612,181);双域双头抗体(unilever;sanofi aventis,wo20100226923),二双抗体(imclone/eli lilly),钮-入-孔抗体形式(genentech,wo9850431);duobody(genmab,wo2011/131746);静电驾驭抗体形式(amgen,ep1870459和wo 2009089004;chugai,us201000155133;oncomed,wo2010129304a2);双特异性igg1和igg2(rinat neurosciences corporation,wo11143545),crossmabs(roche,wo2011117329),luz-y(genentech),biclonic(merus),双靶向域抗体(gsk/domantis),识别两个靶标的二合一抗体(genentech,novlmmune),交联mabs(karmanos cancer center),covx体(covx/pfizer),igg样双特异性(imclone/eli lilly,shen,x,等j immunol methods,2007.318(1-2):p.65-74),和dig体和pig体(pharmabcine),以及双亲和性再靶向分子(macrogenics的fc-dart或ig-dart,wo/2008/157379,wo/2010/080538),zybodies(zyngenia),用普通轻链(crucell/merus,us7262028)或普通重链(novlmmune的κλ体)的方法,以及融合蛋白,其包含多肽序列,所述多肽序列融合于含fc域的抗体片段,像scfv融合体,像zymogenetics/bms的bsab,biogenidec的hercules(us007951918),emergent biosolutions/trubion的scorpions,ts2ab(medlmmune/az dimasi,n.,等j mol biol,2009.393(3):p.672-92),novartis的scfv融合,changzhou adam biotech inc的scfv融合体(cn102250246),roche的tvab(wo 2012025525,wo2012025530),f-star的mab
2
(wo2008/003116)和双的scfv-融合体。还应当理解,术语抗体,除另有指明外,也包括多克隆抗体,单克隆抗体(如人单克隆抗体),抗体混合物(重组多克隆)例如,通过symphogen和merus(oligoclonics)开发的技术生成的,抗体样多肽,例如嵌合抗体和人源化抗体。生成的抗体可以潜在的具有任何同种型。
[0320]
可选的选自由单价抗体,重链抗体,链交换改造域(seed),triomab,双可变域免疫球蛋白(dvd-ig),钮-入-孔抗体,迷你抗体,双亲和性再靶向分子fc-dart或ig-dart);luz-y抗体,双克隆抗体,双靶向(dt)-ig抗体,二合一抗体,交联mab,mab
2
,covx体,igg样双特异性抗体,ts2ab,bsab,hercules抗体,tvab,scfv/fc融合抗体,scoprion,与fc域融合的scfv片段和与fc域融合的双scfv片段。
[0321]
在另一个实施方案中,抗原在细胞表面表达。
[0322]
在另一个实施方案中,细胞是人肿瘤细胞。
[0323]
在进一步的实施方案中,抗原选自下组:erbb1(egfr),erbb2(her2),erbb3,erbb4,muc-1,cd4,cd19,cd20,cd38,cd138,cxcr5,c-met,herv-包膜蛋白,骨膜蛋白,bigh3,sparc,bcr,cd79,cd37,egfrviii,igfr,l1-cam,axl,组织因子(tf),cd74,epcam和mrp3。
[0324]
在另一个实施方案中,抗原与细胞膜相关联。
[0325]
在另一个实施方案中,抗原与病毒粒子相关联,任选地其中抗原包含在病毒粒子的蛋白衣壳或脂质包膜中。
[0326]
在另一个实施方案中,抗体是人抗体,任选地结合至少一个选自cd20和cd38的抗原。
[0327]
在另一个实施方案中,抗体结合与7d8和005中至少一个相同的表位,任选地包含7d8和005中至少一个的可变重和/或可变轻链区域。
[0328]
在根据所公开发明的任何用途中,没有任何本发明突变的抗体可以是任何亲本抗体。因此,本发明的用途提供了这样亲本抗体的任何变体。
[0329]
在本发明进一步的实施方案中,效应子作用是选自c1q结合,补体激活,补体依赖性细胞毒性(cdc),抗体依赖性细胞介导的细胞毒性(adcc),fcrn结合,fc受体结合包括fc-γ受体结合,蛋白a结合,蛋白g结合,抗体依赖性细胞吞噬(adcp),补体依赖性细胞细胞毒性(cdcc),补体增强细胞毒性,调理作用,含fc多肽的内化,靶标下调、adc摄取、诱导细胞凋亡、细胞死亡、细胞周期停滞、以及其任意组合所组成组的fc介导的效应子作用。
[0330]
在具体的实施方案中,效应子作用是c1q结合,补体激活(c1q功效),补体依赖性细胞毒性(cdc),抗体依赖性细胞介导的细胞毒性(adcc),fc受体结合,例如fc-γ受体结合,含fc多肽的内化或其任意组合。
[0331]
在一个实施方案中,效应子作用是c1q结合。
[0332]
在一个实施方案中,效应子作用是补体激活(c1q功效)。
[0333]
在一个实施方案中,效应子作用是补体依赖性细胞毒性(cdc)。
[0334]
在一个实施方案中,效应子作用是抗体依赖性细胞介导的细胞毒性(adcc)。
[0335]
在一个实施方案中,效应子作用是fc受体结合,例如包括fc-γ受体结合
[0336]
在一个实施方案中,效应子作用是含fc多肽的内化。
[0337]
在一个实施方案中,效应子作用是补体依赖性细胞毒性(cdc)和抗体依赖性细胞介导的细胞毒性(adcc)的组合。
[0338]
在另一个实施方案中,一个或多个突变增加了选自fcrn结合,adcc,fc-γ受体结合,蛋白a结合,蛋白g结合,adcp,补体依赖性细胞细胞毒性(cdcc),补体增强细胞毒性,抗体介导受调理的抗体结合到补体受体,及其任意组合的更多效应子作用。
[0339]
在另一方面,本发明涉及增加亲本抗体制剂与c1q亲和力的方法,包含突变该抗体fc区中至少一个氨基酸的步骤,其中至少一个氨基酸选自由e345、e430、s440、p247、i253、s254、q311、d/e356、t359、e382、q386、y436和k447组成的组。
[0340]
本发明中所用的术语“c1q结合”,当用于亲本抗体的变体或抗体的上下文中时,包括由变体或抗体与包括免疫系统各种细胞(如效应细胞)在内的宿主组织或因子结合所介导的补体激活经典途径的第一组分任何机制。抗体的c1q结合可以通过elisa评价(如,例如实施例3和4中所用的c1q结合elisa),或者c1q效力可以通过cdc检测评价(如,例如实施例5中所用的cdc检测)。在进一步的实施方案中,抗体的c1q结合亲和力根据实施例4中描述的检测方法来确定。
[0341]
在根据所公开发明的所有方法中,没有任何本发明突变的抗体可以是任何亲本抗体。因此,本发明的方法提供了这样亲本抗体的任何变体。
[0342]
通过本发明方法和/或用途获得的亲本抗体,第一亲本抗体,第二亲本抗体,或其变体可以连接带本发明所描述的任何靶标上。
[0343]
本发明针对抗的抗原或靶标的实例为:5t4;adam-10;adam-12;adam17;afp;axl;angpt2炭疽抗原;bsg;caix;caxii;ca72-4;癌相关抗原ctaa16.88;ccl11;ccl2;ccr4;ccr5;ccr6;cd2;cd3e;cd4;cd5;cd6;cd15;cd18;cd19;cd20;cd22;cd24;cd25;cd29;cd30;
cd32b;cd33;cd37;cd38;cd40;cd40lg;cd44;cd47;cd52;cd56;cd66e;cd72;cd74;cd79a;cd79b;cd80;cd86;cd98;cd137;cd147;cd138;cd168;cd200;cd248;cd254;cd257;cdh3;cea;ceacam5;ceacam6;ceacam8;claudin4;cs-1;csf2ra;cspg-4;ctla4;cripto;dll4;ed-b;efna2;egfr;内皮缩血管肽b受体;enpp3;epcam;erbb2;erbb3;fapα;fcγri;fcer2;fgfr3;纤维蛋白-β链;flt1;folh 1;folr1;frp-1;gd3神经节苷脂;gdf2;glp1r;磷脂酰肌醇聚糖-3;gpnmb;hbv(乙型肝炎病毒);hcmv(人巨细胞病毒);热休克蛋白90同源物[白色念珠菌];单纯疱疹病毒的gd糖蛋白;hgf;hiv-1;hiv-1iiib gp120 v3环;hla-drb(hla-drβ);人类呼吸道合胞病毒;糖蛋白f;icam-1;ifna1;ifna1;ifnb1双特异性;ige fc;igf1r;ighe连接区;il12b;il13;il15;il17a;il1a;il1b;il2ra;il4;il5;il5ra;il6;il6r;il9;白细胞介素2受体β亚单位;itga2;itga2b itgb3;itga4itgb7;itga5;itgal;itgav_itgb3;itgb2;kdr;l1cam;lewis-y;脂质a;脂多糖lps域;lta;met;mmp14;mmpl5;mst1r;mstn;muc1;muc4;muc16;muc5ac;nca-90粒细胞抗原;连接素4;ngf;nrp;ny-eso-1;ox40l;plac-1;plgf;pdgfra;pd1;pdl1;psca;磷脂酰丝氨酸;ptk-7;铜绿假单胞菌血清型iats o11;rsv(人类呼吸道合胞病毒,糖蛋白f);ror1;rtn4;sell;selp;steap1;志贺样毒素ii b亚单位[大肠杆菌];slam7;slc44a4;sost;表皮葡萄球菌的脂磷壁酸;t细胞受体α_β;tf;tgfb1;tgfb2;tmeff2;tnc;tnf;tnfrsf10a;tnfrsf10b;tnfrsf12a;tnfsf13;tnfsf14;tnfsf2;tnfsf7;trailr2;trop2;tyrp1;vap-1和波形蛋白。
[0344]
诱导效应应答的方法
[0345]
应当理解本发明所描述的关于亲本抗体,第一亲本抗体或第二亲本抗体所有实施方案,也应被理解为涉及包含免疫球蛋白fc域和结合区的亲本,第一亲本或第二亲本多肽的实施方案。
[0346]
在进一步的主要方面,本发明涉及诱导对细胞、细胞膜或病毒粒子的效应应答的方法,所述细胞、细胞膜或病毒粒子表达包含免疫球蛋白fc域和结合区的亲本多肽所结合到的靶标,所述方法包括
[0347]
(i)提供已经被按照权利要求1到24任一项突变的亲本多肽或者至少第一亲本多肽与第二亲本多肽的组合;和
[0348]
(ii)将步骤(i)的突变亲本多肽或步骤(i)的至少第一亲本多肽与第二亲本多肽的突变组合的制剂与表达抗原的细胞、细胞膜或病毒粒子,在人补体或效应细胞的存在下接触。
[0349]
在一个实施方案中,任何或所有的亲本多肽,第一亲本多肽与第二亲本多肽可以是抗体。
[0350]
因此在一个实施方案中,本发明涉及使用本发明描述的抗体变体诱导对与抗原或多种抗原相关联的细胞、细胞膜、病毒粒子或其他颗粒的效应应答,例如补体激活,cdc或其他效应应答的的方法。本发明也涉及诱导对表达亲本抗体所结合抗原的细胞、细胞膜或病毒粒子的效应应答的方法,包括
[0351]
(i)提供已经被按照本发明描述的任何方法突变的亲本抗体或者至少第一亲本抗体与第二亲本抗体的组合;和
[0352]
(ii)将步骤(i)的突变亲本抗体或步骤(i)的至少第一亲本抗体与第二亲本抗体的突变组合的制剂与表达抗原的细胞、细胞膜或病毒粒子,在人补体或效应细胞的存在下
接触。
[0353]
亲本抗体,第一亲本抗体和第二亲本抗体可以各选自本发明描述的任何亲本抗体,具体是关于影响抗体效应子作用方法的任何上述那些抗体。
[0354]
在一个实施方案中,抗原在细胞表面表达。
[0355]
在一个实施方案中,细胞是人肿瘤细胞。
[0356]
在进一步的实施方案中,抗原选自下组:erbb1(egfr),erbb2(her2),erbb3,erbb4,muc-1,cd4,cd19,cd20,cd38,cd138,cxcr5,c-met,herv-包膜蛋白,骨膜蛋白,bigh3,sparc,bcr,cd79,cd37,egfrviii,igfr,l1-cam,axl,组织因子(tf),cd74,epcam和mrp3。
[0357]
在另一个实施方案中,抗原与细胞膜相关联。
[0358]
在另一个实施方案中,抗原与病毒粒子相关联,任选地其中抗原包含在病毒粒子的蛋白衣壳或脂质包膜中。
[0359]
在另一个实施方案中,抗体是人抗体,任选地结合至少一个选自cd20和cd38的抗原。
[0360]
在另一个实施方案中,抗体结合与7d8和005中至少一个相同的表位,任选地包含7d8和005中至少一个的可变重和/或可变轻链区域。
[0361]
在本发明进一步的实施方案中,诱导的效应应答是,补体依赖性细胞毒性(cdc),fc-介导的效应应答,其选自选自如下的fc-介导的效应应答:c1q结合,补体激活,补体依赖性细胞毒性(cdc),抗体依赖性细胞介导的细胞毒性(adcc),fcrn结合,fc受体结合包括fc-γ受体结合,蛋白a结合,蛋白g结合,抗体依赖性细胞吞噬(adcp),补体依赖性细胞细胞毒性(cdcc),补体增强细胞毒性,调理作用,含fc多肽的内化,靶标下调,adc摄取,诱导细胞凋亡,细胞死亡,细胞周期停滞,以及其任意组合。
[0362]
在具体的实施方案中,效应应答是c1q结合,补体激活(c1q功效),补体依赖性细胞毒性(cdc),抗体依赖性细胞介导的细胞毒性(adcc),fc受体结合,例如fc-γ受体结合,含fc多肽内化或其任意组合。
[0363]
在一个实施方案中,效应应答是c1q结合。
[0364]
在一个实施方案中,效应应答是补体激活(c1q功效)。
[0365]
在一个实施方案中,效应应答是补体依赖性细胞毒性(cdc)。
[0366]
在一个实施方案中,效应应答是抗体依赖性细胞介导的细胞毒性(adcc)。
[0367]
在一个实施方案中,效应应答是fc受体结合,例如包括fc-γ受体结合。
[0368]
在一个实施方案中,效应应答是包含fc的多肽内化。
[0369]
在一个实施方案中,效应应答是补体依赖性细胞毒性(cdc)和抗体依赖性细胞介导的细胞毒性(adcc)的组合。
[0370]
在另一个实施方案中,所述方法增加选自以下的进一步效应应答:fcrn结合,adcc,fc-γ受体结合,蛋白a结合,蛋白g结合,adcp,补体依赖性细胞毒性(cdcc),补体增强的细胞毒性,通过抗体介导的与受调理抗体的补体受体的结合,及其任意组合。
[0371]
在另一方面,本发明涉及一种增加亲本抗体制剂对c1q亲和力的方法,包括将抗体fc区至少一个氨基酸突变的步骤,其中所述至少一种氨基酸选自e345、e430、s440、p247、i253、s254、q311、d/e356、t359、e382、q386、y436和k447。
[0372]
本发明针对的抗原或靶标的实例是:5t4;adam-10;adam-12;adam17;afp;axl;angpt2炭疽抗原;bsg;caix;caxii;ca 72-4;癌相关抗原ctaa16.88;ccl11;ccl2;ccr4;ccr5;ccr6;cd2;cd3e;cd4;cd5;cd6;cd15;cd18;cd19;cd20;cd22;cd24;cd25;cd29;cd30;cd32b;cd33;cd37;cd38;cd40;cd40lg;cd44;cd47;cd52;cd56;cd66e;cd72;cd74;cd79a;cd79b;cd80;cd86;cd98;cd137;cd147;cd138;cd168;cd200;cd248;cd254;cd257;cdh3;cea;ceacam5;ceacam6;ceacam8;claudin4;cs-1;csf2ra;cspg-4;ctla4;cripto;dll4;ed-b;efna2;egfr;内皮缩血管肽b受体;enpp3;epcam;erbb2;erbb3;fapα;fcγri;fcer2;fgfr3;纤维蛋白iiβ链;flt1;folh 1;folr1;frp-1;gd3神经节苷脂;gdf2;glp1r;磷脂酰肌醇聚糖-3;gpnmb;hbv(乙型肝炎病毒);hcmv(人巨细胞病毒);热休克蛋白90同源物[白色念珠菌(candida albicans)];单纯疱疹病毒gd糖蛋白;hgf;hiv-1;hiv-1iiib gpl20 v3环;hla-drb(hla-drβ);人呼吸道合胞病毒,糖蛋白f;icam1;ifna1;ifna1;ifnb1双特异性;ige fc;igf1r;ighe连接区;il12b;il13;il15;il17a;il1a;il1b;il2ra;il4;il5;il5ra;il6;il6r;il9;白介素-2受体β亚单位;itga2;itga2b itgb3;itga4 itgb7;itga5;itgal;itgav_itgb3;itgb2;kdr;l1cam;lewis-y;脂质a,脂多糖lps的域;lta;met;mmp14;mmpl5;mst1r;mstn;muc1;muc4;muc16;muc5ac;nca-90粒细胞抗原;连接素4;ngf;nrp;ny-eso-1;ox40l;plac-1;plgf;pdgfra;pd1;pdl1;psca;磷脂酰丝氨酸;ptk-7;铜绿假单胞菌血清型iats o11;rsv(人呼吸道合胞病毒,糖蛋白f);ror1;rtn4;sell;selp;steap1;志贺样毒素ii b亚单位[大肠杆菌];slam7;slc44a4;sost;表皮葡萄球菌脂胞壁酸;t细胞受体α_β;组织因子(tf);tgfb1;tgfb2;tmeff2;tnc;tnf;tnfrsf10a;tnfrsf10b;tnfrsf12a;tnfsf13;tnfsf14;tnfsf2;tnfsf7;trailr2;trop2;tyrp1;vap-1;和波形蛋白。
[0373]
在一个实施方案中,细胞是肿瘤细胞或细菌细胞。
[0374]
在另一个实施方案中,抗原选自erbb1(egfr),erbb2(her2),erbb3,erbb4,muc-1,cd4,cd19,cd20,cd25,cd32,cd37,cd38,cd74,cd138,cxcr5,c-met,herv-包膜蛋白,骨膜蛋白,bigh3,sparc,bcr,cd79,egfrviii,igfr,ll-cam,epcam和mrp3。
[0375]
在进一步的实施方案中,抗原是cd20或cd38。
[0376]
在另一个实施方案中,igg1亲本抗体是人igg1抗体。
[0377]
在另一个实施方案中,亲本抗体选自7d8和005。
[0378]
在一个实施方案中,细胞是人肿瘤细胞。
[0379]
在另一个实施方案中,第一和第二抗原分别选自erbb1(egfr),erbb2(her2),erbb3,erbb4,muc-1,cd4,cd19,cd20,cd25,cd32,cd37,cd38,cd74,cd138,cxcr5,c-met,herv-包膜蛋白,骨膜蛋白,bigh3,sparc,bcr,cd79,egfrviii,igfr,l1-cam,axl,组织因子(tf),epcam和mrp3。
[0380]
在另一个实施方案中,第一和第二亲本抗体是完全人的,任选的其中所述第一和第二亲本抗体分别结合选自cd20和cd38的抗原。
[0381]
在进一步的实施方案中,第一和第二亲本抗体分别选自7d8和005。
[0382]
在甚至进一步的实施方案中,细胞是细菌细胞。
[0383]
在另一个实施方案中,细菌细胞选自金黄色葡萄球菌(s.aureus),表皮葡萄球菌(s.epidermidis),肺炎葡萄球菌(s.pneumonia),炭疽芽孢杆菌(bacillus anthracis),铜绿假单胞菌(pseudomonas aeruginosa),衣原体属(chlamydia),大肠杆菌(e.coli),沙门
氏菌属(salmonella),志贺氏菌属(shigella),耶尔森氏菌属(yersinia),鼠伤寒沙门氏菌(s.typhimurium),脑膜炎奈瑟氏菌(neisseria meningitides)和结核分枝杆菌(mycobacterium tuberculosis)。
[0384]
在另一个实施方案中,第一和/或第二抗原是脂胞壁酸(lta),任选的其中所述第一和第二亲本抗体中的至少一种的是帕昔单抗。
[0385]
在另一个实施方案中,抗原在病毒粒子上表达。
[0386]
在另一个实施方案中,第一和第二抗体结合相同抗原。
[0387]
在另一个实施方案中,第一和第二抗体包括相同的vh序列,vl序列,或vh和vl序列两者。
[0388]
对于本发明的目的来说,表达抗原或否则与抗原结合的靶细胞可以是任意原核或真核细胞。示例性抗原表达细胞包括,但不限于,哺乳动物细胞,尤其是人细胞,例如人癌细胞;和单细胞机体例如细菌,原生动物;和单细胞真菌例如酵母细胞。包括抗原或否则与抗原结合的细胞膜包括来源于抗原表达细胞的部分和/或破坏的细胞膜。与病毒粒子或病毒颗粒有关的抗原可以包括在病毒粒子的蛋白外壳和/或脂质包膜中,或与病毒粒子的蛋白外壳和/或脂质包膜结合。
[0389]
靶细胞可以是例如人肿瘤细胞。合适的肿瘤抗原包括本发明描述的任意靶标或抗原,但不限于,erbb1(egfr),erbb2(her2),erbb3,erbb4,muc-1,cd4,cd19,cd20,cd25,cd32,cd37,cd38,cd74,cd138,cxcr5,c-met,herv-包膜蛋白,骨膜蛋白,bigh3,sparc,bcr,cd79,egfrviii,igfr,l1-cam,axl,组织因子(tf),epcam和mrp3。优选的抗原包括cd20,cd38,her2,egfr,igfr,cd25,cd74和cd32。示例性抗体包括wo 2004/035607公开的抗cd20抗体7d8,wo 06/099875公开的抗cd38抗体005,wo 2004/035607公开的抗cd20抗体11b8,wo 06/099875公开的抗cd38抗体003,wo 02/100348公开的抗egfr抗体2f8。本发明提供其他具体抗体的实例。
[0390]
可选的,靶细胞可以是细菌细胞,例如,例如,金黄色葡萄球菌(s.epidermidis),表皮葡萄球菌(s.epidermidis),肺炎葡萄球菌(s.pneumonia),炭疽杆菌(bacillus anthracis),铜绿假单胞菌(pseudomonas aeruginosa),衣原体属(chlamydia),大肠杆菌(e.coli),沙门氏菌属(salmonella),志贺氏菌属(shigella),耶尔森氏菌属(yersinia),鼠伤寒沙门氏菌(s.typhimurium),脑膜炎奈瑟菌(neisseria meningitides)和结核分枝杆菌(mycobacterium tuberculosis)。示例性抗原包括脂胞壁酸(lta),和示例性抗体包括帕昔单抗。
[0391]
可选的,靶标可以存在于病毒、真菌细胞或其他颗粒的表面,例如,例如,西尼罗河病毒,登革热病毒,丙肝病毒(hcv),人类免疫缺陷病毒(hiv),人乳头瘤病毒,eb病毒,疱疹病毒,痘病毒,禽流感病毒,rvs,曲霉属,白色念珠菌,隐球菌属(cryptococcus),和组织胞浆菌属(histoplasma)。
[0392]
在一个实施方案中,接触步骤(ii)在体外发生。
[0393]
在一个实施方案中,接触步骤(ii)在体内发生。
[0394]
在另一个实施方案中,步骤(ii)包括给受试者施用变体。
[0395]
在进一步的实施方案中,受试者患有癌症,细菌感染,或病毒感染。上述实施方案的接触步骤(ii)可以在体外或体内发生。在后一情况下,步骤(ii)可以进一步包括给受试
者(任选的患有癌症或细菌感染的受试者)施用一种制剂或多种制剂。关于治疗应用的更多细节将在下文提供。
[0396]
第一和第二抗体包括可以结合相同或不同表位的抗原结合区。这类表位可以在相同或不同的靶标上。
[0397]
在一个实施方案中,第一和第二抗体结合不同靶标上的不同表位。这类靶标可以在相同细胞或细胞类型上表达,或可以在不同细胞或细胞类型上表达。在这类实施方案中,效应子作用的增强只针对表达两种靶标的细胞或细胞类型,从而降低对不是待治疗疾病病因的细胞或细胞类型的任何附带损害的风险。
[0398]
不受任何理论约束,普遍认为cdc的增强可限于同时表达两种特定靶标/抗原的靶细胞,条件是第一和第二抗体结合在相同细胞上发现的表位,从而利用靶标的组合表达来提高增强的cdc诱导的选择性。
[0399]
靶标在不同的细胞或细胞类型上表达的情况下,不受理论约束的认为,按照第一和第二抗体的任意顺序的施用将促进cdc增强,并且还可能通过“募集”表达第二靶标的第二细胞或细胞类型来提高其他效应子作用。
[0400]
在其中使用第一和第二抗体的组合的实施方案中,可以在人补体和/或效应细胞存在的条件下将细胞与突变的第一和第二亲本抗体同时、分开或顺序接触来进行步骤(ii)。
[0401]
在另一方面,本发明涉及一种提高亲本抗体制剂的cdc诱导能力的方法,包括将抗体fc区至少一个氨基酸突变的步骤,其中所述至少一种氨基酸选自e345、e430、s440、p247、i253、s254、q311、d/e356、t359、e382、q386、y436和k447。
[0402]
在可选的方面,本发明涉及一种诱导针对表达igg1亲本抗体结合的抗原的细胞、细胞膜或病毒粒子的效应应答(任选的cdc应答)的方法,包括
[0403]
(i)提供在选自以下的至少一个氨基酸残基中包含突变的抗体:igg1重链fc区的e345、e430、s440、p247、i253、s254、q311、d/e356、t359、e382、q386、y436和k447;和
[0404]
(ii)在人补体或效应细胞存在的情况下,将抗体制剂与细胞、细胞膜或病毒粒子接触。
[0405]
在另一个可选实施方案中,所述方法进一步包括施用第一抗体,其在选自对应于第一抗体fc区的e345、e430、s440、p247、i253、s254、q311、d/e356、t359、e382、q386、y436和k447的氨基酸残基的至少一个氨基酸残基中包括第一突变;
[0406]
施用第二抗体,其在选自对应于第二抗体fc区的e345、e430、s440、p247、i253、s254、q311、d/e356、t359、e382、q386、y436和k447的氨基酸残基的至少一个氨基酸残基中包括第二突变;
[0407]
其中第一和第二抗体可以同时、分开或顺序施用。第一和第二抗体可以结合相同或不同靶标上的相同或不同表位。在相同或不同的细胞或细胞类型中可以发现靶标。
[0408]
在另一个可选的方面,本发明涉及一种提高igg1亲本抗体制剂的cdc诱导能力的方法,包括将抗体fc区的至少一个氨基酸突变,其中所述至少一个氨基酸选自e345、e382和h433。
[0409]
在另一个可选的方面,本发明涉及一种诱导针对表达igg1亲本抗体结合的抗原的细胞、细胞膜或病毒粒子的效应应答(任选的cdc应答)的方法,包括以下步骤:
[0410]
(i)提供亲本抗体的变体,其在抗体fc区的至少一个氨基酸中包含突变,所述至少一个氨基酸选自e345、e382和h433;和
[0411]
(ii)在人补体或效应细胞存在的情况下,将变体制剂与细胞接触。
[0412]
在另一个可选的方面,本发明涉及一种诱导针对表达igg1亲本抗体结合的抗原的细胞的效应应答(任选的cdc应答)的方法,包括以下步骤:
[0413]
(i)提供亲本抗体的变体,其在抗体的fc区包括k439e和s440k突变;和
[0414]
(ii)在人补体或效应细胞存在的情况下,将变体制剂与细胞接触。
[0415]
在另一个可选的方面,本发明涉及一种诱导针对表达第一igg1亲本抗体结合的第一抗原和第二亲本抗体结合的第二抗原的细胞、细胞膜或病毒粒子的cdc应答的方法,包括以下步骤:
[0416]
(i)提供第一亲本抗体包括k439e突变的第一变体和第二亲本抗体包括s440k突变的第二变体;和
[0417]
(ii)在人补体和/或效应细胞存在的条件下,将细胞与第一和第二变体同时、分开或顺序地接触。
[0418]
在另一个可选的方面,本发明提供一种诱导针对靶细胞、细胞膜、病毒粒子或其他颗粒的cdc或其他效应应答的方法,所述靶细胞、细胞膜、病毒粒子或其他颗粒与igg1或igg3抗体结合的抗原有关,包括步骤:(i)提供抗体变体,其在对应于igg1抗体fc区的e345、e430或s440的至少一个氨基酸中包含突变;和(ii)在人补体和/或效应细胞存在的条件下将变体制剂与细胞接触。
[0419]
在另外可选的方面,本发明提供一种诱导针对靶细胞、细胞膜、病毒粒子或其他颗粒的adcc或adcp,或靶细胞、细胞膜、病毒粒子或其他颗粒的吞噬的方法,所述靶细胞、细胞膜、病毒粒子或其他颗粒与igg1或igg3抗体结合的抗原有关,包括步骤:(i)提供抗体变体,其在对应于igg1抗体fc区的e345、e430或s440的至少一个氨基酸中包含突变;和(ii)在效应细胞存在的条件下将变体制剂与细胞接触。
[0420]
本发明还提供一种诱导针对与igg1或igg3抗体结合的抗原有关的靶细胞、细胞膜、病毒粒子或其他颗粒的cdc或其他效应应答的方法,包括步骤:(i)提供抗体变体,其包括在抗体fc区的k439的突变(k439e)和s440的突变(s440k或s440r);和(ii)在人补体和/或效应细胞存在的条件下将变体制剂与细胞接触。
[0421]
本发明还提供一种诱导针对靶细胞、细胞膜或病毒粒子的cdc或其他效应应答的方法,所述靶细胞、细胞膜或病毒粒子表达第一igg1抗体结合的第一抗原和第二抗体结合的第二抗原,包括步骤:(i)提供第一变体(其是包括k439e突变的第一抗体)和第二变体(其是包括s440k或s440r突变的第二抗体);和(ii)在人补体或效应细胞存在的条件下将细胞与第一和第二变体制剂同时、分开或顺序地接触。
[0422]
在单独和具体的实施方案中,第一和第二抗体结合(i)不同抗原,(ii)相同抗原上的不同表位,(iii)抗原上相同表位,和(iv)抗原上的相同表位并包括相同的vh和/或vl序列。
[0423]
在一个实施方案中,第一和第二抗体进一步包括e345,e430和s440的一个或更多个的突变,例如e345r。在一个实施方案中,第一和第二抗体进一步包括e345,e382和h433的一个或更多个的突变,例如e345r。
[0424]
其他方法
[0425]
在另一个主要方面,本发明涉及一种鉴定增强抗体结合c1q的效应子作用的抗体突变的方法,包括步骤:
[0426]
(i)制备至少一种抗体,其在选自e345、e430、s440、k439、p247、i253、s254、q311、d/e356、t359、e382、q386、y436和k447的至少一个氨基酸中包含突变;
[0427]
(ii)当结合抗原表达细胞的表面时,与亲本抗体比较,评估所述抗体的c1q活性;和
[0428]
(iii)挑选具有c1q亲和力增加的任意变体的突变。
[0429]
在一个实施方案中,至少一种抗体包括选自e345r、e345q、e345n、e345k、e345y、e430t、e430s、e430g、e430f、e430h、s440w和s440y的至少一个氨基酸取代。
[0430]
仍在另一个主要方面,本发明涉及一种鉴定增加抗体诱导cdc应答能力的亲本抗体突变的方法,包括步骤:
[0431]
(i)制备至少一种亲本抗体变体,其在选自e345、e430、s440、k439、p247、i253、s254、q311、d/e356、t359、e382、q386、y436和k447的至少一个氨基酸中包含突变;
[0432]
(ii)在效应细胞或补体存在的条件下,当结合抗原表达细胞的表面时,与亲本抗体比较,评估所述变体诱导的cdc应答;和
[0433]
(iii)挑选具有c1q应答增加的任意变体的突变。
[0434]
在一个实施方案中,至少一个氨基酸选自e345、e382和h433。
[0435]
在一个实施方案中,至少一种抗体包括选自e345r、e345q、e345n、e345k、e345y、e430t、e430s、e430g、e430f、e430h、s440w和s440y的至少一个氨基酸取代。
[0436]
在另一个方面,本发明涉及一种增加igg1亲本抗体制剂对c1q亲和力的方法,包括将抗体fc区的至少一个氨基酸突变,其中所述至少一个氨基酸选自e345、e382和h433。
[0437]
本发明的抗体
[0438]
亲本抗体
[0439]
如本发明所述,本发明尤其涉及在免疫球蛋白的ch2和/或ch3区域(例如在抗体重链)包括一个或更多个突变的亲本抗体变体。在修饰前将用作本发明起始原料的“亲本”抗体,其可以是野生型抗体,可以例如通过kohler等,nature 256,495(1975)首次描述的杂交瘤方法产生,或通过重组dna方法产生。使用描述于例如clackson等,nature 352,624 628(1991)和marks等,j.mol.biol.222,581 597(1991)的技术,也可以从噬菌体抗体文库分离单克隆抗体。单克隆抗体可以获自任意合适的来源。因此,例如,单克隆抗体可以获自从鼠脾b细胞制备的杂交瘤,所述鼠脾b细胞获自用感兴趣的抗原免疫的小鼠,所述感兴趣的抗原例如采用以下形式:在表面表达抗原的细胞,或编码感兴趣抗原的核酸。单克隆抗体也可以获自来源于免疫的人或非人哺乳动物(例如兔,大鼠,狗,灵长类动物,等等)的抗体表达细胞的杂交瘤。
[0440]
亲本抗体可以是例如嵌合或人源化抗体。在另一个实施方案中,抗体是人抗体。可以利用转基因或转染色体小鼠(例如humab小鼠,携带部分人免疫系统而不是小鼠系统)来产生人单克隆抗体。humab小鼠包含编码未重排的人重链(μ和γ)和κ轻链免疫球蛋白序列的人免疫球蛋白基因小位点(minilocus),以及使内源μ和κ链基因座失活的靶向突变(lonberg,n.等,nature368,856 859(1994))。因此,小鼠显示降低的小鼠igm或κ表达,并且
响应于免疫,导入的人重链和轻链转基因,经历类型转换和体细胞突变以产生高亲和力人igg,k单克隆抗体(lonberg,n.等(1994),上文;综述于lonberg,n.handbook of experimental pharmacology 113,49 101(1994),lonberg,n.和huszar,d.,intern.rev.immunol.vol.13 65 93(1995)以及harding,f.和lonberg,n.ann.n.y.acad.sci 764 536 546(1995))。humab小鼠的制备详述于taylor,l.等,nucleic acids research 20,6287 6295(1992),chen,j.等,international immunology 5,647 656(1993),tuaillon等,j.immunol.152,29122920(1994),taylor,l.等,international immunology 6,579 591(1994),fishwild,d.等,nature biotechnology 14,845 851(1996)。还可参见us 5,545,806,us5,569,825,us 5,625,126,us 5,633,425,us 5,789,650,us 5,877,397,us5,661,016,us 5,814,318,us 5,874,299,us 5,770,429,us 5,545,807,wo98/24884,wo 94/25585,wo 93/1227,wo 92/22645,wo 92/03918和wo01/09187。根据公知技术,可以使用来自这些转基因小鼠的脾细胞产生分泌人单克隆抗体的杂交瘤。
[0441]
此外,本发明的人抗体或本发明来自其他种属的抗体可以通过展示类技术来鉴定,包括,但不限于,噬菌体展示,逆转录病毒展示,核糖体展示,哺乳动物展示,酵母展示和本领域已知的其他技术,并且所获分子可以进行附加的成熟,例如亲和力成熟,因为这类技术是本领域公知的。描述于实施例17的具体策略适用于任意抗体以使用噬菌体展示来制备并获得本发明的变体。
[0442]
亲本抗体不限于具有天然的例如人fc域的抗体,但是它还可以是具有除本发明那些之外的其他突变的抗体,例如例如影响糖基化或使抗体成为双特异性抗体的突变。术语“自然抗体”表示不包括任何遗传导入突变的任意抗体。包括天然发生的修饰(例如不同的同种异型)的抗体因此就本发明来说被理解为“天然抗体”,从而可以被理解为亲本抗体。这种抗体可以作为根据本发明的一个或更多个突变的模板,从而提供本发明的变体抗体。包括除本发明那些之外的其他突变的亲本抗体的实例是描述于wo2011/131746(genmab)的双特异性抗体,利用还原条件以促进包括igg4样ch3区域的两种抗体的半分子替换,从而形成双特异性抗体而无聚集体的伴发形成。亲本抗体的其他实例包括但不限于双特异性抗体例如异二聚体双特异性∶triomabs(fresenius);双特异性igg1和igg2(rinat neurosciences corporation);fc
△
adp(regeneron);钮-入-孔(genentech);静电驾驭(amgen,chugai,oncomed);seed体(merck);azymetric scaffold(zymeworks);mab-fv(xencor);和luz-y(genentch)。其他示例性亲本抗体形式包括,但不限于,野生型抗体,全长抗体或包含fc的抗体片段,人抗体,或其任意组合。
[0443]
亲本抗体可以结合任意靶标,本发明这种靶标或抗原的实例可以是,并且不限于,针对的是:5t4;adam-10;adam-12;adam 17;afp;axl;angpt2炭疽抗原;bsg;caix;caxii;ca 72-4;癌相关抗原ctaa16.88;ccl11;ccl2;ccr4;ccr5;ccr6;cd2;cd3e;cd4;cd5;cd6;cd15;cd18;cd19;cd20;cd22;cd24;cd25;cd29;cd30;cd32b;cd33;cd37;cd38;cd40;cd40lg;cd44;cd47;cd52;cd56;cd66e;cd72;cd74;cd79a;cd79b;cd80;cd86;cd98;cd137;cd147;cd138;cd168;cd200;cd248;cd254;cd257;cdh3;cea;ceacam5;ceacam6;ceacam8;claudin4;cs-1;csf2ra;cspg-4;ctla4;cripto;dll4;ed-b;efna2;egfr;内皮缩血管肽b受体;enpp3;epcam;erbb2;erbb3;fapα;fcγri;fcer2;fgfr3;纤维蛋白iiβ链;flt1;folh1;
folr1;frp-1;gd3神经节苷脂;gdf2;glp1r;磷脂酰肌醇聚糖-3;gpnmb;hbv(乙型肝炎病毒);hcmv(人巨细胞病毒);热休克蛋白90同源物[白色念珠菌(candida albicans)];单纯疱疹病毒gd糖蛋白;hgf;hiv-1;hiv-1iiib gpl20 v3环;hla-drb(hla-drβ);人呼吸道合胞病毒,糖蛋白f;icam1;ifna1;ifna1;ifnb1双特异性;ige fc;igf1r;ighe连接区;il12b;il13;il15;il17a;ilia;il1b;il2ra;il4;il5;il5ra;il6;il6r;il9;白介素-2受体β亚单位;itga2;itga2b itgb3;itga4itgb7;itga5;itgal;itgav_itgb3;itgb2;kdr;l1cam;lewis-y;脂质a,脂多糖lps的域;lta;met;mmp14;mmpl5;mst1r;mstn;muc1;muc4;muc16;muc5ac;nca-90粒细胞抗原;连接素4;ngf;nrp;ny-eso-1;ox40l;plac-1;plgf;pdgfra;pd1;pdl1;psca;磷脂酰丝氨酸;ptk-7;铜绿假单胞菌血清型iats o11;rsv(人呼吸道合胞病毒,糖蛋白f);ror1;rtn4;sell;selp;steap1;志贺样毒素ii b亚单位[大肠杆菌];slam7;slc44a4;sost;表皮葡萄球菌脂胞壁酸;t细胞受体α_β;tf;tgfb1;tgfb2;tmeff2;tnc;tnf;tnfrsf10a;tnfrsf10b;tnfrsf12a;tnfsf13;tnfsf14;tnfsf2;tnfsf7;trailr2;trop2;tyrp1;vap-1;和波形蛋白。
[0444]
亲本抗体可以是任意同种型的任意人抗体,例如igg1、igg2、igg3、igg4、iga1、iga2、ige和igd,任选的人全长抗体,例如人全长igg1抗体。亲本抗体可以包括根据seq id nos:1,2,3,4和5中任一项的序列。
[0445]
用于本发明的单克隆抗体,例如亲本和/或变体,可以通过kohler等,nature 256,495(1975)首先描述的杂交瘤方法产生,或通过重组dna方法产生。还可以使用在例如clackson等,nature 352:624-628(1991)和marks等,j.mol.biol.222:581-597(1991)中描述的技术,从噬菌体抗体文库分离单克隆抗体。单克隆抗体可以获自任意合适的来源。因此,例如,单克隆抗体可以从鼠脾b细胞制备的杂交瘤获得,所述鼠脾b细胞获自用感兴趣的抗原(例如采用以下形式:在表面表达抗原的细胞,或编码感兴趣抗原的核酸)免疫的小鼠。单克隆抗体还可以从来源于免疫的人或非人哺乳动物(例如大鼠,狗,灵长类动物,等等)的抗体表达细胞的杂交瘤获得。
[0446]
在一个实施方案中,抗体是人抗体。可以利用携带部分人免疫系统而不是小鼠系统的转基因或转染色体小鼠来产生针对任意抗原的人单克隆抗体。这种转基因和转染色体小鼠分别包括在本发明中被称为小鼠和km小鼠的小鼠,在本发明中统称为“转基因小鼠”。
[0447]
小鼠包含编码未重排人重链(μ和γ)和κ免疫球蛋白轻链序列的人免疫球蛋白基因小基因座,以及使内源μ和κ链基因座失活的靶向突变(lonberg,n.等,nature 368,856-859(1994))。因此,小鼠显示小鼠igm或κ表达的降低,并且响应于免疫,引入的人重链和轻链转基因,经历类型转换和体细胞突变以产生高亲和力人igg,κ单克隆抗体(lonberg,n.等(1994),上文;综述于lonberg,n.handbook of experimental pharmacology 113,49-101(1994),lonberg,n.和huszar,d.,intern.rev.immunol.vol.13 65-93(1995)以及harding,f.和lonberg,n.ann.n.y.acad.sci 764 536-546(1995))。小鼠的制备详细描述于taylor,l.等,nucleic acids research 20,6287-6295(1992),chen,j.等,international immunology 5,647-656(1993),tuaillon等,j.immunol.152,2912-2920(1994),taylor,l.等,international immunology 6,579-591(1994),fishwild,d.等,nature biotechnology 14,845-851(1996)。还可参见us 5,545,
806,us 5,569,825,us 5,625,126,us5,633,425,us 5,789,650,us 5,877,397,us 5,661,016,us 5,814,318,us5,874,299,us 5,770,429,us 5,545,807,wo 98/24884,wo 94/25585,wo93/1227,wo 92/22645,wo 92/03918和wo 01/09187。
[0448]
hco7、hco12、hco17和hco20小鼠在其内源轻链(κ)基因具有jkd破坏(描述于chen等,embo j.12,821-830(1993)),在其内源重链基因(描述于wo 01/14424的实施例1)和kco5人κ轻链转基因(描述于fishwild等,nature biotechnology 14,845-851(1996))具有cmd破坏。此外,hco7小鼠具有hco7人重链转基因(描述于us 5,770,429),hco12小鼠具有hco12人重链转基因(描述于wo 01/14424的实施例2),hco17小鼠具有hco17人重链转基因(描述于wo 01/09187的实施例2)和hco20小鼠具有hco20人重链转基因。所获小鼠在纯合的用于破坏内源小鼠重链和κ轻链基因座的背景下表达人免疫球蛋白重链和κ轻链转基因。
[0449]
在km小鼠品系中,如chen等,embo j.12,811-820(1993)所描述,内源小鼠κ轻链基因已经被纯合破坏,并且如wo 01/09187的实施例1所描述,内源小鼠重链基因已经被纯合破坏。如fishwild等,nature biotechnology 14,845-851(1996)所描述,这种小鼠品系携带有人κ轻链转基因kco5。如wo02/4347814所描述,这种小鼠品系还携带由14号染色体片段hcf(sc20)组成的人重链转染色体。如wo/2009/097006所述,通过将hco12与kco5[j/k](balb)杂交,产生hco12-balb/c小鼠。根据公知技术,可以使用来自这些转基因小鼠的脾细胞产生分泌人单克隆抗体的杂交瘤。
[0450]
此外,利用本领域公知的技术,通过展示类技术(包括但不限于,噬菌体展示,逆转录病毒展示,核糖体展示和其他技术)鉴定,可以从人抗体或其他种属的抗体获得任意抗原结合区,并且所获分子可以经历附加的成熟,例如亲和力成熟,这类技术是本领域公知的(参见例如hoogenboom等,j.mol.biol.227,381(1991)(噬菌体展示),vaughan等,nature biotech 14,309(1996)(噬菌体展示),hanes和plucthau,pnas usa 94,4937-4942(1997)(核糖体展示),parmley和smith,gene 73,305-318(1988)(噬菌体展示),scott tibs 17,241-245(1992),cwirla等,pnas usa 87,6378-6382(1990),russel等,nucl.acids research 21,1081-1085(1993),hogenboom等,immunol.reviews 130,43-68(1992),chiswell和mccafferty tibtech 10,80-84(1992),和us 5,733,743)。如果展示技术用于产生非人抗体,则这种抗体可以被人源化。
[0451]
在另一个方面,本发明涉及一种包括fc域和结合区的亲本多肽。在本发明的上下文中应理解涉及亲本抗体的所有实施方案类似地适用于“亲本多肽”。
[0452]
根据本发明的突变可以是,但不限于,一个或更多个氨基酸的缺失、插入或取代。氨基酸的这种取代可以是利用任意天然存在的或非天然氨基酸的取代。
[0453]“单突变体”[0454]
应理解本发明中所有实施方案参考亲本抗体进行描述,还应将第一亲本抗体或第二亲本抗体理解为有关包括免疫球蛋白和结合区的亲本、第一亲本或第二亲本多肽的实施方案。
[0455]
根据本发明“单突变体”方面的抗体或多肽变体包括在表1所示的至少一个氨基酸残基的突变,通常是氨基酸取代,表1列出了根据人igg1抗体的eu索引进行编号的每个氨基酸残基,连同igg2、igg3和igg4亲本抗体对应位置的氨基酸以及“示例性”和“优选的”氨基酸取代。图2显示了对应于残基p247到k447的igg2区段,对应于残基p247到k447的igg3 fc
区段,和igg1中对应于残基p247到k447的igg4区段。
[0456]
表1
-“
单突变体”方面的示例突变位点和氨基酸取代
[0457][0458]
从表1可以看出,导致实施例19的wien133细胞的细胞裂解增加的氨基酸取代被包括作为“优选的取代”。
[0459]
在一个方面,本发明涉及一种亲本多肽的变体,其包括免疫球蛋白的fc域和结合区,其中所述变体包括在选自对应于以下氨基酸残基的至少一个氨基酸残基的突变:人igg1重链fc区的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447,条件是s440的突变是s440y或s440w。
[0460]
在一个实施方案中,变体多肽可以是变体抗体。
[0461]
因此在另一个方面,本发明涉及一种抗体的变体,其包括免疫球蛋白的抗原结合区和fc域,其中所述变体包括在选自对应于以下氨基酸残基的至少一个氨基酸残基的突变:人igg1重链fc区的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447,条件是s440的突变是s440y或s440w。可选的,所述氨基酸残基选自对应于以下的氨基酸残基:人igg1重链fc区的h310、g385、h433、n434、q438和k439。
[0462]
对应于人igg1重链fc区中e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447的每一氨基酸残基可以如上所述根据以下分组:
[0463]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0464]
(b)在ch2-ch3区的疏水钮(knobs)内的氨基酸残基,
[0465]
(c)n端ch3螺旋内的氨基酸残基,
[0466]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0467]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基。
[0468]
根据本发明的突变可以是,但不限于,一个或更多个氨基酸的缺失、插入或取代。氨基酸的这种取代可以是利用任意天然存在的或非天然氨基酸的取代。因此,在一个实施
方案中,至少一个氨基酸残基的突变是缺失。在另一个实施方案中,至少一个氨基酸残基的突变是插入。在另一个实施方案中,至少一个氨基酸残基的突变是取代。
[0469]
在一个实施方案中,至少一个氨基酸残基的突变选自对应于以下的氨基酸残基:人igg1重链fc区的e345x、e430x、s440w/y、q386k、p247g、i253v、s254l、q311l/w、d/e356r、e382v、y436i和k447d/e/缺失,其中x是任意氨基酸,例如天然存在的氨基酸。
[0470]
在一个具体实施方案中,抗体变体包括选自下组的至少一个氨基酸残基的突变:人igg1重链fc区的e345、e430、s440和q386。
[0471]
在进一步的实施方案中,至少一个氨基酸残基的突变是选自对应于以下的氨基酸取代:人igg1重链fc区的e345x、e430x、s440w/y、q386k,其中x是任意氨基酸,例如天然存在的氨基酸。
[0472]
在优选的实施方案中,至少一个氨基酸残基的突变是选自对应于以下氨基酸取代的氨基酸取代:人igg1重链fc区的e345r,q,n,k,a,c,d,f,g,h,i,l,m,p,s,t,v,w,y;e430t,s,g,a,c,d,f,h,i,l,k,m,n,p,q,r,v,w,y;s440w,y和q386k。
[0473]
在进一步优选的实施方案中,至少一个氨基酸残基的突变是选自对应于以下氨基酸取代的氨基酸取代:人igg1重链fc区的e345r/q/n/k、e430t/s/g、s440y/w和q386k。
[0474]
可选的,至少一个氨基酸残基选自e382和h433。具体可选的氨基酸取代包括e345y,d,w;和e430f,h。可选的,e382d,q,k,r;和h433r。
[0475]
在一个具体实施方案中,氨基酸取代是e345r。在可选的实施方案中,突变选自i253到e,n,q,s或t;h310到n,q,w或y;q311到e或r;e382到d,h,k,r,n,q,s,t,w或y;g385到e,h,k,n,q,r,s,t,w或y;h433到r;n434到d,e,h,k,q,r,s,t,w或y;y436到a,e,f,h,i,k,l,m,n,q,r,s,t或v;q438到a,e,g,h,k,n,q,r,s,t,w或y;k439到d,h,q,r,w或y;和s440到d,e,h,f,n,q,w或y。
[0476]
在另一个可选的实施方案中,突变选自i253到n或q;h310到q;q311到e或r;e382到d,q,k,或r;g385到d,e,k或r;h433到r;n434到h,k,q或r;y436到n,q,s或t;q438到n,s或t;k439到q;和s440到d,e或q。
[0477]
在另一个可选实施方案中,突变选自e382到d,q,k或r;和h433到r。
[0478]
在一个实施方案中,变体包括e382r突变。
[0479]
在一个实施方案中,变体包括h433r突变。
[0480]
如实施例所示,包括上述氨基酸取代之一的cd38抗体humab-005和-003和/或cd20抗体humab-7d8和-11b8和利妥昔单抗和/或egfr抗体humab-2f8的变体分别具有比野生型humab 005和7d8更高的c1q结合,补体激活和/或cdc。应理解所述变体也可以只包括表1所列“示例性取代”中的一个突变。所述变体可以包括超过一个突变,例如表1所列任意突变中的两,三,四,五或六个。
[0481]
因此,本发明优选的实施方案提供在选自上述方面所列的氨基酸残基中包括一个突变的变体。具体的氨基酸突变可以是对应于以下氨基酸取代任一项的氨基酸取代:p247g,i253v,s254l,q311l,q311w,e345a,e345c,e345d,e345f,e345g,e345h,e345i,e345k,e345l,e345m,e345n,e345p,e345q,e345r,e345s,e345t,e345v,e345w,e345y,d/e356g,d/e356r,t359r,e382l,e382v,q386k,e430a,e430c,e430d,e430f,e430g,e430h,e430i,e430k,e430l,e430m,e430n,e430p,e430q,e430r,e430s,e430t,e430v,e430w,
e430y,y436i,s440y和s440w。如实施例19,表17所示,这些具有增加的细胞裂解(对于wien133细胞>39%)。
[0482]
在可选的实施方案中,所述变体包括在选自对应于e382r,h433r,h435r和h435a的氨基酸残基中的一个氨基酸残基的突变。
[0483]
除了所述指定突变外,变体可以具有针对亲本抗体描述的任意特征。具体来说,它可以是人抗体。除了突变外,变体可以进一步是任意igg1亚型。
[0484]
当与抗原表达细胞表面、细胞膜、病毒粒子或另一颗粒上的其抗原结合时,或抗原与病毒粒子有关时,任选的其中抗原包括在病毒粒子的蛋白衣壳或脂质包膜内,与亲本抗体相比,这类抗体变体可以具有以下至少一种的增加:(i)c1q结合,(ii)抗体介导的补体激活,(iii)抗体介导的cdc,(iv)寡聚体形成,(v)寡聚体稳定性,或(i)到(v)中任意的组合。在(iv)或(v)的一个实施方案中,寡聚体是六聚体。在一个实施方案中,变体还可以或者可选的具有保留的或改善的其他效应子作用,例如c1q结合,补体激活,补体依赖性细胞毒性(cdc),抗体依赖性细胞介导的细胞毒性(adcc),fcrn结合,fc受体结合包括fc-γ受体结合,蛋白a结合,蛋白g结合,抗体依赖性细胞吞噬(adcp),补体依赖性细胞的细胞毒性(cdcc),补体增强的细胞毒性,调理作用,包含fc的多肽内化,靶标下调,adc摄取,凋亡诱导,细胞死亡,细胞周期停滞,及其任意组合。
[0485]
不受限于任何具体理论,用表1的氨基酸残基在指定位置取代氨基酸引起的功效可以是,例如,引起自身的功效,参与直接接触另一分子的fc域,或可以被突变以与另一fc域直接相互作用或间接影响分子间fc:fc相互作用。因此,不受限于理论,取代被认为直接或间接增强寡聚形式的抗体分子间的结合强度,增强寡聚体结构(例如六聚体,五聚体,四聚体,三聚体或二聚体结构)的稳定性。例如,氨基酸取代可以是促进或增强新分子间fc:fc键(例如,但不限于,范德华相互作用,氢键,电荷-电荷相互作用)形成的氨基酸取代,或在fc:fc相互作用下通过水分子释放促进熵增加的氨基酸取代。就表1而言,可以基于参与或促进分子间fc:fc相互作用或分子内相互作用(异构突变)的大小和物化特性挑选“示例性取代”。可以基于最适合参与或激发分子间fc:fc相互作用或分子内相互作用(异构突变)的大小和物化特性挑选“示例性取代”。
[0486]
表1所列氨基酸的“示例性取代”包括用e残基替换r残基,和用h残基替换r残基。表1所列每个具体氨基酸残基中氨基酸的每个“示例性取代”是本发明单独和具体无限制的实施方案。此外,表1所列每个具体氨基酸残基中氨基酸的每个“优选的取代”是本发明单独和具体无限制的实施方案。
[0487]
在另一个方面,本发明涉及一种亲本多肽的变体,其包括免疫球蛋白的fc域和结合区,其中所述变体在选自下组的至少两个氨基酸残基中包含突变:
[0488]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0489]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0490]
(c)n端ch3螺旋内的氨基酸残基,
[0491]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0492]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,
[0493]
其中至少两个氨基酸突变是不同的。
[0494]
在一个实施方案中,亲本多肽及其变体可以是抗体。
[0495]
因此,本发明还涉及一种亲本抗体的变体,其包括抗原结合区和fc域,其中所述变体在选自下组的至少两个氨基酸残基中包含突变:
[0496]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0497]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0498]
(c)n端ch3螺旋内的氨基酸残基,
[0499]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0500]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,
[0501]
其中至少两个氨基酸突变是不同的。
[0502]
因此,上述实施方案的变体可以包括表1中至少两个,例如两、三、四、五或更多个氨基酸的突变。
[0503]
在变体包括至少两个氨基酸的这种突变的任意实施方案中,它可以存在于变体的每一重链中,或两个中的一个包括在一条重链内,其他的分别包括在其他重链内,或反之亦然。
[0504]
在一个实施方案中,变体包括在选自对应于以下氨基酸残基的至少两个氨基酸残基的突变:人igg1重链fc区中的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447,条件是s440的突变是s440y或s440w。可选的,变体进一步包括选自以下的至少一个残基的突变:h310、g385、h433、n434、q438和k439。
[0505]
在一个实施方案中,变体包括选自对应于以下氨基酸残基的至少两个氨基酸残基的突变:人igg1重链fc区的e345x、e430x、s440w/y、q386k、p247g、i253v、s254l、q311l/w、d/e356r、e382v和y436i,其中x是任意氨基酸,例如天然存在的氨基酸。
[0506]
例如,抗体变体可以包括以下至少一个突变:e345、e430、s440和q386,可选的e382和h433,例如e345、e430、s440和q386中的两个或全部,可选的e382和h433,任选的进一步包括表1所列一个或更多个其他氨基酸的突变。因此,在进一步的实施方案中,变体包括选自对应于以下的至少两个氨基酸残基的突变:人igg1重链fc区的e345x、e430x、s440w/y和q386k,其中x是任意氨基酸,例如天然存在的氨基酸。
[0507]
至少两个氨基酸残基的突变的示例性组合是e345x/e430x、e345x/s440y或w、e345x/q386k、e430x/s440y或w和e430x/q386k。
[0508]
在一个实施方案中,至少两个氨基酸残基的突变是缺失,插入或取代。氨基酸的这种取代可以是利用任意天然存在的或人工氨基酸的取代。
[0509]
在具体的实施方案中,至少两个氨基酸残基的突变可以是对应于以下任意的氨基酸取代:p247g、i253v、s254l、q311l、q311w、e345a、e345c、e345d、e345f、e345g、e345h、e345i、e345k、e345l、e345m、e345n、e345p、e345q、e345r、e345s、e345t、e345v、e345w、e345y、d/e356g、d/e356r、t359r、e382l、e382v、q386k、e430a、e430c、e430d、e430f、e430g、e430h、e430i、e430k、e430l、e430m、e430n、e430p、e430q、e430r、e430s、e430t、e430v、e430w、e430y、y436i、s440y和s440w。
[0510]
在优选的实施方案中,变体包括至少两个氨基酸残基的突变,所述突变是选自对应于以下氨基酸取代的氨基酸取代:人igg1重链fc区的e345r,q,n,k,a,c,d,f,g,h,i,l,m,
p,s,t,v,w,y;e430t,s,g,a,c,d,f,h,i,l,k,m,n,p,q,r,v,w,y;s440w,y和q386k。
[0511]
可选的进一步突变选自对应于以下的突变:i253e,n,q,s,t;h310n,q,w,y;q311e,r;e382d,h,k,r,n,q,s,t,w,y;g385e,h,k,n,q,r,s,t,w,y;h433r;n434d,e,h,k,q,r,s,t,w,y;y436,a,e,f,h,i,k,l,m,n,q,r,s,t,v;q438a,e,g,h,k,n,q,r,s,t,w,y;k439d,h,q,r,w,y;和s440d,e,h,f,n,q。
[0512]
在优选的实施方案中,至少两个氨基酸残基的突变是选自对应于以下氨基酸取代的氨基酸取代:人igg1重链fc区中的e345r/q/n/k,e430t/s/g,s440y/w和q386k。可选的进一步突变选自对应于以下的突变:i253n,q;h310q;q311e,r;e382d,q,k,r;g385d,e,k,r;h433r;n434h,k,q,r;y436n,q,s,t;q438n,s,t;k439q;和s440d,e,q。
[0513]
至少两个氨基酸残基突变的示例性具体组合是e345r/e430t,e345r/s440y,e345r/s440w,e345r/q386k,e345r/e430g,e345q/e430t,e345q/s440y,e345q/s440w,e430t/s440y,e430t/s440w,e430t/q386k,和s440y/q386k。
[0514]
在一个具体实施方案中,突变不是位于对应于i253、n434或q311的氨基酸残基。在另外的或可选的实施方案中,突变不是位于h433,或氨基酸取代不是h433a。
[0515]
在一个实施方案中,本发明涉及一种包括至少三个氨基酸残基的突变的变体,所述氨基酸残基选自下组:
[0516]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0517]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0518]
(c)n端ch3螺旋内的氨基酸残基,
[0519]
(d)c端ch3β链内的氨基酸残基,条件是它不包括对应于人igg1重链fc区中s440的突变,和
[0520]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基。
[0521]
其中至少三个氨基酸突变是不同的。
[0522]
在一个具体实施方案中,变体包括氨基酸残基的突变,其是对应于e345r、q396k和e430g的氨基酸取代,其可以位于变体的一条或两条重链。
[0523]
至少三个氨基酸残基的突变可以单独选自表1所列的取代。包括至少三个突变的变体的非限制性实例是:e345r/e430g/s440y,e345r/e430g/s440w,e345k/e430g/s440y,e345k/e430g/s440w,e345q/e430g/s440y,e345q/e430g/s440w,e345n/e430g/s440y,e345n/e430g/s440w,e345r/e430t/s440y,e345r/e430t/s440w,e345k/e430t/s440y,e345k/e430t/s440w,e345q/e430t/s440y,e345q/e430t/s440w,e345n/e430t/s440y,e345n/e430t/s440w,e345r/e430s/s440y,e345r/e430s/s440w,e345k/e430s/s440y,e345k/e430s/s440w,e345q/e430s/s440y,e345q/e430s/s440w,e345n/e430s/s440y,e345n/e430s/s440w,e345r/e430f/s440y,e345r/e430f/s440w,e345k/e430f/s440y,e345k/e430f/s440w,e345q/e430f/s440y,e345q/e430f/s440w,e345n/e430f/s440y,和e345n/e430f/s440w。
[0524]
除了表1或2a和b中一个或更多个氨基酸的突变,igg重链可以包括本领域已知的另外突变,例如,进一步改善效应子作用的突变。这类另外突变包括增强cdc,fc-受体结合或fcrn结合和/或改善fc-γ受体介导的效应子作用的已知突变。
[0525]
在一个实施方案中,本发明的变体还包括已知的cdc增强修饰,例如,igg同种型之
间的片段替换以产生嵌合igg分子(natsume等,2008cancer res68(10),3863-72);铰链区的一个或更多个氨基酸取代(dall'acqua等,2006j immunol 177,1129-1138),和/或围绕残基d270、k322、p329和p331,在ch2域的c1q结合位点中或附近的一个或更多个氨基酸取代(idusogie等,2001j immunol 166,2571-2575;michaelsen等,2009scand j immunol 70,553-564和wo 99/51642)。例如,在一个实施方案中,本发明的变体还包括氨基酸取代s267e,h268f,s324t,s239d,g236a和i332e中任一项的组合,通过cdc或adcc提供增强的效应子作用(moore等,2010mabs 2(2),181-189))。影响与fc受体结合(描述于wo 2006/105062,wo 00/42072,美国专利us6,737,056和美国专利us7,083,784)或抗体物理特性(描述于wo 2007/005612 a1)的其他fc突变也可用于本发明的变体。
[0526]
在一个实施方案中,本发明的变体还包括增强fc-γ受体结合和/或fc-γ受体介导的效应子作用的修饰。这种修饰包括(i)降低ch2相连的糖基化中岩藻糖的量(糖工程改造)(umana p,等,nat biotechnol 1999;17:176-80;niwa r,等,clin cancer res 2004;10:6248-55.)),和(ii)抗体铰链或ch2区域中氨基酸的定点诱变(蛋白工程改造)(lazar ga,等,proc natl acad sci u s a2006;103:4005-10)。
[0527]
在一个实施方案中,本发明的变体在fcrn结合位点被进一步改造,例如,以延长igg抗体的半衰期(t1/2)。这类修饰包括(i)n434a和t307a/e380a/n434a突变(petcova等int immunol.2006dec;18(12):1759);(ii)提高fcrn结合的将pro238,thr256,thr307,gln311,asp312,glu380,glu382和asn434中的一个或更多个取代为丙氨酸残基(shields rl,等j.biol.chem.2001;276:6591);和(iii)igg1的氨基酸取代或氨基酸取代的组合,组合选自:m252y/s254t/t256e,m252w,m252y,m252y/t256q,m252f/t256d,v308t/l309p/q311s,g385d/q386p/n389s,g385r/q386t/p387r/n389p,h433k/n434f/y436h,n434f/y436h,h433r/n434y/y436h,m252y/s254t/t256e-h433k/n434f/y436h或m252y/s254t/t256e-g385r/q386t/p387r/n389p,增加对fcrn的亲和力(dall'acqua等,上文)。
[0528]“双突变体”[0529]
应理解本发明中所有实施方案参考亲本抗体进行描述,还应将第一亲本抗体或第二亲本抗体理解为有关包括免疫球蛋白和结合区的亲本、第一亲本或第二亲本多肽的实施方案。
[0530]
如上下文所述,本发明还涉及“双突变体”方面,其中两个突变单独的话各自降低效应子作用但共同将效应子作用恢复到亲本抗体水平。当一起使用时,变体的特异性增加。根据“双突变体”方面的抗体变体包括两个突变,通常是特异性氨基酸残基相互作用对k439和s440的氨基酸取代。
[0531]
因此在一个方面,本发明涉及一种多肽的变体,其包括免疫球蛋白的fc域和结合区,其中所述变体在以下位置包含突变:
[0532]
(i)选自对应于人igg1重链fc区的k439和s440的氨基酸残基的至少一个氨基酸残基,条件是s440的突变不是s440y或s440w,例如其中在对应于人igg1重链fc区的k439位置的突变是k439d/e,和/或在对应于人igg1重链fc区的s440位置的突变是s440k/h/r;
[0533]
(ii)对应于k447d/e或对应于人igg1重链fc区的k447k/r/h和448p的至少一个氨基酸残基;或
[0534]
(iii)对应于k447d/e或对应于人igg1重链fc区的k447k/r/h和448k/r/h和449p的
至少一个氨基酸残基。
[0535]
在一个实施方案中,亲本多肽及其变体可以是抗体。
[0536]
因此,在一个方面,本发明涉及一种抗体的变体,其包括免疫球蛋白的抗原结合区和fc域,其中所述变体在选自对应于人igg1重链fc区中的k439和s440的氨基酸残基的至少一个氨基酸残基中包含突变,条件是s440的突变不是s440y或s440w,例如其中对应于人igg1重链fc区中的k439位置的突变是k439d/e,和/或对应于人igg1重链fc区k440位置的突变是s440k/h/r。表2a和b显示“双突变体”(表a)和“混合突变体”(表2b)方面的“示例性”和“优选的取代”。
[0537]
表2a“双突变体”方面的示例突变位点和氨基酸取代
[0538][0539]
表2b:“混合突变体”方面(抗体1+抗体2)的示例突变位点和氨基酸取代
[0540][0541]
在一个变体实施方案中,其中突变位于s440和k447之外的位置并且其中变体进一步包括在以下位置的突变:
[0542]
(i)对应于人igg1重链fc区的k439或s440的至少一个氨基酸残基,条件是s440的突变不是s440w或s440y;
[0543]
(ii)对应于人igg1重链fc区的k447d/e或对应于k447k/r/h和448p的至少一个氨基酸残基;或
[0544]
(iii)对应于人igg1重链fc区的k447d/e或对应于k447k/r/h和448k/r/h和449p的至少一个氨基酸残基。
[0545]
因此,本发明提供一种抗体变体,其包括对应于k439的人igg1重链ch2和/或ch3区域的残基的第一突变,和对应于s440的人igg1重链ch2和/或ch3区域的残基的第二突变。
[0546]
本发明预期变体也可以只包括一个氨基酸残基取代,例如k439e或s440k,例如所述变体在k439包含突变,任选的在s440没有突变。
[0547]
在一个实施方案中,本发明涉及变体,其中k439的突变是氨基酸取代为选自e和d的氨基酸,例如k439e。
[0548]
在另一个实施方案中,变体包括s440的突变,任选的在k439没有突变。
[0549]
在一个实施方案中,本发明涉及变体,其中s440的突变是氨基酸取代为选自e、r和h的氨基酸,例如s440k。
[0550]
在一个实施方案中,变体包括k439和s440两处的突变。
[0551]
在另一个实施方案中,k439的突变选自k439到d、e或r,s440的突变选自s440到d、e、k、h和r。
[0552]
在另一个实施方案中,k439的突变选自k439d和k439e,和s440的突变选自s440k、s440r和s440h。
[0553]
在另一个实施方案中,变体包括k439e和s440k突变。
[0554]
如实施例4-6所述,只包括k439e和s440k突变之一的抗体变体对c1q的k
d
剧烈增加,反映降低的补体激活和/或cdc能力。出人意料的,发现包括两个突变的humab 7d8或005的抗体变体具有恢复的或增加的c1q结合或cdc。不受任何具体理论限制,根本机制也许可以通过相应突变空间上彼此弥补来解释,如图4和5所示。
[0555]
本发明描述的任意“双突变体”也可以与自身能够增加效应子作用的突变组合使用。因此,“双突变体”方面可以与“单突变体”方面组合,例如变体可以进一步包括表1所列任意氨基酸位置或针对上述“单突变体”方面描述的任意其他实施方案的突变。因此,在一个实施方案中,突变位于s440之外的位置,和其中变体进一步包括对应于人igg1重链fc区的k439或s440的至少一个氨基酸残基的突变,条件是s440的突变不是s440w或s440y。
[0556]
在一个方面,本发明涉及一种多肽的变体,其包括免疫球蛋白的fc域和结合区,其中所述变体在选自下组的至少一个氨基酸残基中包含突变:
[0557]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0558]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0559]
(c)n端ch3螺旋内的氨基酸残基,
[0560]
(d)c端ch3β链内的氨基酸残基,条件是它不包括对应于人igg1重链fc区中s440的突变,和
[0561]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,和
[0562]
其中所述变体包括进一步突变
[0563]
(i)在对应于人igg1重链fc区的k439或s440的至少一个氨基酸残基,条件是s440的突变不是s440w或s440y;
[0564]
(ii)在对应于人igg1重链fc区的k447d/e或对应于k447k/r/h和448p的至少一个氨基酸残基;或
[0565]
(iii)在对应于人igg1重链fc区的k447d/e或对应于k447k/r/h和448k/r/h和449p的至少一个氨基酸残基。
[0566]
在一个实施方案中,亲本多肽及其变体可以是抗体。
[0567]
因此在一个方面,本发明涉及一种亲本抗体的变体,其包括免疫球蛋白的抗原结合区和fc域,其中所述变体包括选自下组的至少一个氨基酸残基的突变:
[0568]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0569]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0570]
(c)n端ch3螺旋内的氨基酸残基,
[0571]
(d)c端ch3β链内的氨基酸残基,条件是它不包括对应于人igg1重链fc区中s440的突变,和
[0572]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,和
[0573]
其中所述变体包括在如下位置的进一步突变
[0574]
(i)在对应于人igg1重链fc区的k439或s440的至少一个氨基酸残基,条件是s440的突变不是s440w或s440y;
[0575]
(ii)在对应于人igg1重链fc区的k447d/e或对应于k447k/r/h和448p的至少一个氨基酸残基;或
[0576]
(iii)在对应于k447d/e或对应于人igg1重链fc区的k447k/r/h和448k/r/h和449p
的至少一个氨基酸残基。
[0577]
在一个实施方案中,变体包括选自对应于以下氨基酸残基的至少一个氨基酸残基的突变:人igg1重链fc区中的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447,条件是s440的突变是s440y或s440w,并且变体包括对应于人igg1重链fc区中的k439或s440的至少一个氨基酸残基的进一步突变,条件是s440的突变不是s440w或s440y。
[0578]
在进一步的实施方案中,变体包括至少一个氨基酸残基的突变,其是选自对应于人igg1重链fc区的e345x,e430x,s440w/y,q386k的氨基酸取代的氨基酸取代,其中x是任意氨基酸,例如天然存在的氨基酸,和变体包括对应于人igg1重链fc区的k439或s440的至少一个氨基酸残基的进一步突变,条件是s440的突变不是s440w或s440y。
[0579]
在一个实施方案中,变体包括对应于igg1重链fc区的k439和s440两个位置的氨基酸突变,条件是s440的突变不是s440y或s440w。
[0580]
在进一步的实施方案中,对应于人igg1重链fc区的k439位置的突变是k439d/e,和/或对应于人igg1重链fc区的s440位置的突变是s440k/h/r。
[0581]
在进一步的实施方案中,第一突变位于选自对应于人igg1重链fc区的e345,e430,q386和s440的氨基酸残基,条件是s440的突变是s440y或s440w;以及第二和第三突变是位置k439e或s440k的氨基酸取代。
[0582]
在一个实施方案中,第一突变是缺失,插入或取代。这种取代可以是任意天然存在的或非天然的氨基酸。
[0583]
在进一步的实施方案中,第一突变选自e345r,q,n,k,a,f,g,h,i,l,m,p,s,t,v,w,y,c,d;e430t,s,g,a,f,h,l,p,r,v,c,d,i,k,m,n,q,w,y;和s440w,y,d;以及第二和第三突变是位置k439e或s440k的氨基酸取代。
[0584]
在优选的实施方案中,一个突变选自e345r,q,n,k,y;e430t,s,g,f,h;s440w,y;和q386k。
[0585]
另一个实施例,在本发明的一个实施方案中,变体包括e345r,k439e和s440k突变,从而提供增加的并且更特异性介导的cdc应答。
[0586]
在一个实施方案中,变体包括选自对应于以下氨基酸残基的至少两个氨基酸残基的突变:人igg1重链fc区中的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447,条件是s440的突变是s440y或s440w,并且变体包括对应于人igg1重链fc区中的k439或s440的至少一个氨基酸残基的进一步突变,条件是s440的突变不是s440w或s440y。
[0587]
在进一步的实施方案中,变体包括至少两个氨基酸残基的突变,其是选自对应于人igg1重链fc区的e345x,e430x,s440w/y,q386k的氨基酸取代,其中x是任意氨基酸,例如天然存在的氨基酸,和变体包括对应于人igg1重链fc区的k439或s440的至少一个人氨基酸残基的进一步突变,条件是s440的突变不是s440w或s440y。
[0588]
在一个实施方案中,变体包括对应于igg1重链fc区的k439和s440两个位置的氨基酸突变,条件是s440的突变不是s440y或s440w。
[0589]
在进一步的实施方案中,对应于人igg1重链fc区的k439位置的突变是k439d/e,和/或对应于人igg1重链fc区的s440位置的突变是s440k/h/r。
[0590]
在进一步的实施方案中,第一和第二突变位于选自对应于人igg1重链fc区的e345,e430,q386和s440的氨基酸残基的氨基酸残基上,条件是s440的突变是s440y或s440w;以及第三和第四突变是位置k439e或s440k的氨基酸取代。
[0591]
在另一个实施方案中,与亲本抗体或只包括k439和s440之一突变的抗体变体相比,如本发明所述包括位置k439和s440的突变的变体具有fc介导的效应子作用的增加,所述效应子作用选自c1q结合,补体激活,补体依赖性细胞毒性(cdc),抗体依赖性细胞介导得细胞毒性(adcc),fcrn结合,fc受体结合包括fc-γ受体结合,蛋白a结合,蛋白g结合,抗体依赖性细胞吞噬(adcp),补体依赖性细胞的细胞毒性(cdcc),补体增强的细胞毒性,调理作用,包含fc的多肽内化、靶标下调、adc摄取、凋亡诱导、细胞死亡、细胞周期停滞、及其任意组合。
[0592]
本发明还提供抗体中k439e和s440k突变在与亲本抗体(其可以是,例如,野生型抗体或只包括k439e或s440k突变之一的抗体变体)相比,恢复以下一种或更多种作用中的用途:(i)c1q结合亲和力,(ii)抗体介导的补体激活,(iii)抗体介导的cdc,(iv)寡聚体形成,(v)寡聚体稳定性,或(i)到(v)中任意的组合。在(iv)或(v)的一个实施方案中,寡聚体是六聚体。
[0593]
在一个实施方案中,变体选自单特异性抗体,双特异性抗体或多特异性抗体。
[0594]
混合突变体
[0595]
应理解本发明中所有实施方案参考亲本抗体进行描述,还应将第一亲本抗体或第二亲本抗体理解为有关包括免疫球蛋白和结合区的亲本、第一亲本或第二亲本多肽的实施方案。
[0596]
如上所述,本发明的本发明人还发现存在自身降低效应子作用但一起使用时恢复效应子作用的突变,例如人igg1重链fc区的位置k439和s440的突变。因此,通过在一个抗体中引入k439并在另一抗体中引入s440,这一原理也可用于确保两种不同抗体的配对。因此,根据“混合突变体”方面的抗体变体包含突变,但是通常导致相同fc分子间fc:fc相互作用降低或大量降低的一个突变。但是,作为“混合突变体”的本发明抗体变体能够彼此配对;提供与例如每种单独的突变体或亲本抗体的混合相比,特异性抗体变体对的恢复或甚至增加的c1q结合,补体激活,cdc,寡聚体形成,和/或寡聚体稳定性。在本发明的一个实施方案中,寡聚体是六聚体。在一个实施方案中,抗体变体对还可以或者可选的具有保留的或改善的其他效应子作用,例如c1q结合,补体激活,补体依赖性细胞毒性(cdc),抗体依赖性细胞介导的细胞毒性(adcc),fcrn结合,fc受体结合包括fc-γ受体结合,蛋白a结合,蛋白g结合,抗体依赖性细胞吞噬(adcp),补体依赖性细胞的细胞毒性(cdcc),补体增强的细胞毒性、调理作用、包含fc的多肽内化、靶标下调、adc摄取、凋亡诱导、细胞死亡、细胞周期停滞、及其任意组合。本发明的这种方面提供大量应用,其中不仅c1q结合、补体激活、cdc或其他效应子作用的强度并且其选择性均可以被调节。
[0597]“混合突变体”对中每种抗体变体的示例性突变位点显示于表2。具体来说,本发明提供一种包括免疫球蛋白抗原结合区和fc域的抗体变体,所述变体在对应于k439和s440之一的人igg1重链fc区的残基包含突变。在一个实施方案中,突变位于k439,并且是是氨基酸取代为选自e或d的氨基酸,例如k439e。在一个实施方案中,突变位于s440,并且是氨基酸取代为选自k、r或h的氨基酸,例如s440k。
[0598]
因此在一个实施方案中,本发明还涉及一种包括至少一个氨基酸残基的突变的变体,所述氨基酸残基选自:
[0599]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0600]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0601]
(c)n端ch3螺旋内的氨基酸残基,
[0602]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0603]
(e)对应于人igg1重链fc区的e345、e382或q386的氨基酸残基;
[0604]
和对应于人igg1重链fc区的k439的氨基酸残基。
[0605]
在另一个实施方案中,本发明还涉及一种包括至少一个氨基酸残基的突变的变体,所述氨基酸残基选自:
[0606]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0607]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0608]
(c)n端ch3螺旋内的氨基酸残基,
[0609]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0610]
(e)对应于人igg1重链fc区的e345、e382或q386的氨基酸残基;
[0611]
和对应于人igg1重链fc区的s440的氨基酸残基。
[0612]
在一个实施方案中,上述两种实施方案可以组合成本发明的“混合突变体”对。
[0613]“混合突变体”对的每个变体可以进一步包括表1所列的氨基酸突变。
[0614]
在本发明的一个实施方案中,“混合突变体”对包括亲本抗体的第一变体和亲本抗体的第二变体,其中第一变体包括免疫球蛋白的第一fc域和抗原结合区,其中所述第一变体包括(i)选自除了k439突变之外的以下氨基酸残基组的至少一个氨基酸残基的第一突变:
[0615]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0616]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0617]
(c)n端ch3螺旋内的氨基酸残基,
[0618]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0619]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,和
[0620]
(ii)在对应于人igg1重链fc区的k439位置的第二突变;和其中第二变体包括免疫球蛋白的第二fc域和抗原结合区,其中所述第二变体包括(i)选自除了s440突变之外的以下氨基酸残基的至少一个氨基酸残基的第一突变:
[0621]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0622]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0623]
(c)n端ch3螺旋内的氨基酸残基,
[0624]
(d)c端ch3β链内的氨基酸残基,和
[0625]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,
[0626]
和(ii)在对应于igg1重链fc区的s440位置的第二突变,条件是s440的突变不是
s440y或s440w。
[0627]
其他示例性“混合突变体”对可以进一步包括,和不限于,下列对的任意一项:包含突变k447e的第一变体和包含突变k447/p448的第二变体;包含突变k447e的第一变体和包含突变k447/k448/p449的第二变体。
[0628]
在一个实施方案中,第一突变是缺失、插入或取代。氨基酸的这种取代可以是利用任意天然存在的或非天然氨基酸的取代。在一个实施方案中,突变是缺失。在另一个实施方案中,突变是插入。在另一个实施方案中,突变是氨基酸的取代。
[0629]
在一个具体实施方案中,第一变体和/或第二变体包括选自对应于以下氨基酸残基的至少一个氨基酸残基的突变:人igg1重链fc区中的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447,条件是s440的突变是s440y或s440w。
[0630]
在一个具体实施方案中,第一变体和/或第二变体包括至少一个氨基酸残基的突变,所述突变可以是对应于以下任意的氨基酸取代:p247g,i253v,s254l,q311l,q311w,e345a,e345c,e345d,e345f,e345g,e345h,e345i,e345k,e345l,e345m,e345n,e345p,e345q,e345r,e345s,e345t,e345v,e345w,e345y,d/e356g,d/e356r,t359r,e382l,e382v,q386k,e430a,e430c,e430d,e430f,e430g,e430h,e430i,e430k,e430l,e430m,e430n,e430p,e430q,e430r,e430s,e430t,e430v,e430w,e430y,y436i,s440y和s440w,和第一变体包括对应于人igg1重链fc区的k439位置的第二突变;和第二变体包括对应于人igg1重链fc区的s440位置的第二突变。
[0631]
例如,在一个实施方案中,“混合突变体”对的一个变体包括e345r和k439e突变,而其他变体包括e345r和s440k突变,因此提供增加和更具特异性的c1q结合亲和力,补体激活,cdc,寡聚体形成,寡聚体稳定性,和/或其他效应相关功能例如fcrn结合,adcc,fc-γ受体结合,蛋白a结合,蛋白g结合,adcp,cdcc,补体增强的细胞毒性,抗体介导的吞噬,内化,凋亡,与受调理抗体的补体受体的结合,和/或其组合。
[0632]“混合突变体”方面还可以包括两种变体,每种变体在人igg1重链fc区包括表1所列的超过一种突变,例如第一变体包含突变s440k/k447e,和第二变体包含突变k439e/k447/p448;例如第一变体包含突变k439e/k447e,和第二变体包含突变s440k/k447/p448。
[0633]
本发明描述的“混合突变体”对中的变体可以衍生自相同或不同的亲本抗体。此外,“混合突变体”方面还可以用于双特异性或非对称抗体。此外,第一、第二和第三抗体可以结合相同或不同靶标上的不同表位。
[0634]
此外,“混合突变体”方面通过利用针对第一抗原的具有k439e突变的第一抗体和针对第二抗原的具有s440k或s440r突变的第二抗体,提供更特异性的针对表达两种特异性肿瘤抗原的肿瘤细胞的cdc或其他效应应答。通过利用包括三种变体的“混合突变体”方面,任选的是双特异性抗体,可以提供更特异性的针对表达至少两种,例如两、三、四、五或六种特异性肿瘤抗原的肿瘤细胞的cdc或其他效应应答。
[0635]
在“单突变体”,“双突变体”和“混合突变体”方面的任意一实施方案中,变体选自单特异性抗体,双特异性抗体或多特异性抗体。
[0636]
在“混合突变体”方面的任意实施方案中,第一,第二和/或第三变体可以包括表1所列的任意氨基酸取代的相同或不同突变。
[0637]
多特异性抗体
[0638]
应理解本发明中所有实施方案参考亲本抗体进行描述,还应将第一亲本抗体或第二亲本抗体理解为有关包括免疫球蛋白和结合区的亲本、第一亲本或第二亲本多肽的实施方案。
[0639]
应理解本发明描述的“单突变体”,“双突变体”和“混合突变体”方面的任意实施方案可用于如下所述的多特异性抗体方面。
[0640]
在一个主要方面,本发明涉及一种变体,其是包括第一多肽(包括免疫球蛋白的第一ch2-ch3区和第一抗原结合区)和第二多肽(包括免疫球蛋白的第二ch2-ch3区和第二抗原结合区)的双特异性抗体,其中第一和第二抗原结合区结合相同抗原或不同抗原上的不同表位,以及其中
[0641]
第一和第二ch2-ch3区各自包括选自对应于以下氨基酸残基的至少一个氨基酸残基第一突变:人igg1重链fc区的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447,条件是s440的突变是s440y或s440w。
[0642]
在一个实施方案中,第一突变是缺失,插入或取代。氨基酸的这种取代可以是利用任意天然存在的或非天然氨基酸的取代。
[0643]
本发明的双特异性抗体不限于具体形式,它可以是上文和此处描述的任意一项。
[0644]
在本发明的一个实施方案中,第一和第二多肽包括选自以下的氨基酸残基的氨基酸残基第一突变:人igg1重链fc区的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447。
[0645]
在一个具体的实施方案中,至少一个氨基酸残基的突变可以是对应于以下任意的氨基酸取代:p247g,i253v,s254l,q311l,q311w,e345a,e345c,e345d,e345f,e345g,e345h,e345i,e345k,e345l,e345m,e345n,e345p,e345q,e345r,e345s,e345t,e345v,e345w,e345y,d/e356g,d/e356r,t359r,e382l,e382v,q386k,e430a,e430c,e430d,e430f,e430g,e430h,e430i,e430k,e430l,e430m,e430n,e430p,e430q,e430r,e430s,e430t,e430v,e430w,e430y,y436i,s440y和s440w。
[0646]
在一个具体实施方案中,双特异性抗体具有wo 2011/131746描述的形式。因此,在一个实施方案中,变体是双特异性抗体,其中第一多肽包括选自对应于人igg1重链fc区的k409,t366,l368,k370,d399,f405和y407的氨基酸残基的进一步氨基酸残基突变;和第二多肽包括选自对应于人igg1重链fc区的f405,t366,l368,k370,d399,y407和k409的氨基酸残基的进一步氨基酸残基突变,和其中第一多肽的所述进一步突变不同于第二多肽的所述进一步突变。
[0647]
在一个具体实施方案中,双特异性抗体具有第一多肽(包括对应于人igg1重链fc区k409的氨基酸残基的进一步突变)和第二多肽(包括对应于人igg1重链fc区f405的氨基酸残基的进一步突变)。可以按照实施例22的描述产生本发明的这种双特异性抗体。此外,可以利用实施例23使用的测定法检测所产生异二聚体蛋白对cdc杀伤的作用。
[0648]
在一个具体实施方案中,双特异性抗体包括第一和第二多肽,其中第一多肽包括对应于人igg1重链fc区k409的氨基酸残基的突变;第二多肽包括对应于人igg1重链fc区f405的氨基酸残基的突变;以及第一和/或第二多肽包括对应于人igg1重链fc区的氨基酸取代e345r的氨基酸残基的进一步突变。
[0649]
在一个具体实施方案中,双特异性抗体包括第一和第二多肽,其中第一多肽包括
对应于人igg1重链fc区k409的氨基酸残基的突变;第二多肽包括对应于人igg1重链fc区f405的氨基酸残基的突变;以及第一和/或第二多肽各自包括对应于人igg1重链fc区的氨基酸取代e345r和q386k的氨基酸残基的进一步突变。所述进一步突变可位于第一和第二多肽两者,或e345r可以位于第一多肽和q386k位于第二多肽;或反之亦然。
[0650]
在一个具体实施方案中,双特异性抗体包括第一和第二多肽,其中第一多肽包括对应于人igg1重链fc区k409的氨基酸残基的突变;第二多肽包括对应于人igg1重链fc区f405的氨基酸残基的突变;以及第一和/或第二多肽各自包括对应于人igg1重链fc区的氨基酸取代e345r、q386k和e430g的氨基酸残基的进一步突变。所述突变可位于第一和第二多肽两者,或第一多肽可以包含突变e345r和e430g,和第二多肽可以包含突变q386k;或反之亦然。
[0651]
双特异性抗体可以,例如,包括cd20抗体的抗原结合区和cd38抗体的抗原结合区,以及表1和/或2所列一个或更多个氨基酸的氨基酸取代。示例性cd20结合区包括wo2004/035607(其因此完整引入作为参考)描述的奥法木单抗(2f2)、7d8和11b,和利妥昔单抗(wo 2005/103081)的cd20结合区。示例性cd38结合区包括wo2006/099875(其因此完整引入作为参考)描述的003和daratumumab(005)的那些cd38结合区。
[0652]
在一个实施方案中,双特异性抗体结合相同或不同的靶标上的不同表位。
[0653]
在另一个实施方案中,第一和第二多肽中的第一突变可以是相同或不同的。
[0654]
在“单突变体”、“双突变体”、“混合突变体”和多特异性抗体方面的一个实施方案中,变体是人igg1、igg2、igg3、igg4、iga1、iga2、igd或ige抗体,任选的人全长抗体,例如人全长igg1抗体。
[0655]
在任意“单突变体”、“双突变体”、“混合突变体”方面和多特异性抗体方面,根据实施例4描述的测定法确定抗体的c1q结合,根据实施例5、6或10描述的测定法确定cdc,突变不位于直接参与c1q结合的氨基酸残基,任选的通过将根据实施例3elisa测定法的c1q结合与根据实施例4基于细胞的测定法的c1q结合进行比较来确定,以及根据实施例12描述的测定法确定adcc。
[0656]
此外,本发明提供如上所述的任意“单突变体”、“双突变体”、“混合突变体”和多特异性抗体方面或实施方案的变体的制品。本发明还提供包括如上所述的任意“双突变体”方面和实施方案的变体的组合物,例如,药物组合物。本发明还提供任意这种变体、制品或组合物作为药物的用途。
[0657]
本发明的上述“单突变体”、“双突变体”、“混合突变体”和多特异性抗体方面尤其适用于具有igg1重链的人抗体分子,所述igg1重链包括对应于人igg1重链恒定区下划线残基130到330(uniprot登录号p01857;seq id no:1)的相关片段p247到k447:
[0658][0659]
本发明还可以用于具有人igg2重链部分的抗体分子。igg1重链的氨基酸残基p247
到k447对应于igg2重链恒定区的下划线残基126到326(登录号p01859;seq id no:2)
[0660][0661]
本发明还可以用于具有人igg3重链部分的抗体分子。igg1重链的氨基酸残基p247到k447对应于igg3重链恒定区的残基177到377(uniprot登录号p01860;seq id no:3),在下面用下划线表示:
[0662][0663]
本发明还可以用于具有人igg4重链部分的抗体分子。igg1重链的氨基酸残基p247到k447对应于igg4重链恒定区的下划线残基127到327(登录号p01859,seq id no:4)
[0664][0665]
本发明还可以用于具有人igg1m(f)同种异型重链部分的抗体。igg1m(f)同种异型的氨基酸序列(ch3序列用下划线表示)-seq id no:5
[0666][0667]
igg1,igg2,igg3,igg4和igg1m(f)恒定区相应片段的比对显示于图2。因此,表1或表2a和b描述的氨基酸的任意突变可以引入通过比对确定的igg2,igg3,igg4和/或igg1m(f)的等同位置,以获得本发明的变体。
[0668]
在一个实施方案中,本发明提供全长igg1,igg2,igg3或igg4抗体的变体,包括如上所述任意方面的一个或更多个氨基酸取代。
[0669]
在任意“单突变体”、“双突变体”、“混合突变体”方面和多特异性抗体中,igg1重链的fc区可以包括以下序列:seq id no:1的残基130到330,seq id no:2的残基126到326,
seq id no:3的残基177到377,或seq id no:4的残基127到327。
[0670]
在一个实施方案中,亲本抗体包括选自seq id no.:1-5的序列,例如seq id no.:1,seq id no.:2,seq id no.:3,seq id no.:4或seq id no.:5。
[0671]
在一个实施方案中,igg1重链的fc区包括seq id no:1的残基130到330的序列。
[0672]
亲本抗体可以是本发明描述的任意亲本抗体。在此上下文处亲本抗体还旨在是第一亲本和第二亲本抗体。
[0673]
在一个实施方案中,亲本抗体是人igg1,igg2,igg3或igg4,iga1,iga2,igd或ige抗体。
[0674]
在一个实施方案中,亲本抗体是人全长抗体,例如人全长igg1抗体。
[0675]
在一个实施方案中,亲本抗体、第一亲本抗体和第二亲本抗体是人igg1抗体,例如igg1m(za)或igg1m(f)同种异型,任选的包括包含seq id no:1或5的fc区。
[0676]
在一个实施方案中,亲本抗体是人igg2抗体,任选的包括包含seq id no:2的fc区。
[0677]
在一个实施方案中,亲本抗体是人igg3抗体,任选的包括包含seq id no:3的fc区。
[0678]
在一个实施方案中,亲本抗体是人igg4抗体,任选的包括包含seq id no:4的fc区。
[0679]
在“单突变体”、“双突变体”、“混合突变体”和多特异性抗体方面的任意一项的具体实施方案中,变体包括与seq id nos:1,2,3,4和5的氨基酸p247到k447具有至少70%,72%,74%,76%,78%,80%,81%,82%,83%,84%,85%,86%,87%,88%,89%,90%,91%,92%,93%,94%,95%,96%,97%,98%或至少约99%的同一性程度的氨基酸序列,除了根据本发明引入的突变之外。
[0680]
因此,除了本发明限定的任意突变之外,变体可以包括seq id no:1,seq id no:2,seq id no:3,seq id no:4或seq id no:5的序列。
[0681]
根据本发明的任意上述“单突变体”、“双突变体”、“混合突变体”和多特异性方面应理解为包括下列实施方案。
[0682]
在一个实施方案中,第一和/或第二亲本抗体是抗体片段,任选的选自单价抗体,重链抗体,链交换改造域(seed),triomab,双可变域免疫球蛋白(dvd-ig),钮-入-孔(knob-into-holes)抗体,小抗体,双亲和力再导向分子(fc-dart或ig-dart);luz-y抗体,biclonic抗体,双靶向(dt)-ig抗体,二合一(two-in-one)抗体,交联mab,mab
2
,covx体,igg样双特异性抗体,ts2ab,bsab,hercules抗体,tvab,scfv/fc融合抗体,scorpion,与fc域融合的scfv片段,和与fc域融合的双重scfv片段。
[0683]
在进一步的实施方案中,第一和第二亲本抗体均结合在人肿瘤细胞表面表达的抗原。
[0684]
在进一步的实施方案中,针对第一和第二亲本抗体的抗原单独选自erbb1(egfr),erbb2(her2),erbb3,erbb4,muc-1,cd4,cd19,cd20,cd38,cd138,cxcr5,c-met,herv-包膜蛋白,骨膜蛋白,bigh3,sparc,bcr,cd79,cd37,egfrviii,l1-cam,axl,组织因子(tf),cd74,epcam和mrp3。
[0685]
在进一步的实施方案中,第一和第二亲本抗体是完全人的。
[0686]
在进一步的实施方案中,针对第一和第二亲本抗体的抗原是,按照任意顺序,选自cd20和cd38,任选的其中第一和第二亲本抗体是,按照任意顺序,选自7d8和005。
[0687]
在进一步的实施方案中,第一抗体和第二抗体均结合在细菌细胞或病毒粒子表面表达的抗原。
[0688]
在另一个实施方案中,细菌细胞选自金黄色葡萄球菌(s.aureus),表皮葡萄球菌(s.epidermidis),葡萄球菌肺炎(s.pneumonia),炭疽杆菌(bacillus anthracis),绿针假单胞菌(pseudomonas aeruginosa),沙眼衣原体(chlamydia trachomatis),大肠杆菌(e.coli),沙门氏菌(salmonella),志贺氏菌(shigella),耶尔森氏菌(yersinia),鼠伤寒沙门氏菌(s.typhimurium),脑膜炎奈瑟菌(neisseria meningitides)和结核分枝杆菌(mycobacterium tuberculosis)。
[0689]
在进一步的实施方案中,第一和第二亲本抗体结合相同的抗原。
[0690]
在另一个实施方案中,第一和第二亲本抗体是相同的抗体。
[0691]
在另一个实施方案中,亲本抗体选自7d8和005。组合物
[0692]
应理解本发明中所有实施方案参考亲本抗体进行描述,还应将第一亲本抗体或第二亲本抗体理解为有关包括免疫球蛋白和结合区的亲本、第一亲本或第二亲本多肽的实施方案。
[0693]
本发明还涉及包括变体的组合物,并且亲本抗体可以是本发明描述的任意变体和亲本抗体。具体方面和实施方案将在下文描述。此外,可以根据本发明描述的任意方法获得这种变体。
[0694]
在一个方面,本发明涉及一种包括亲本多肽的第一变体和亲本多肽的第二变体的组合物,其中第一变体包括免疫球蛋白的第一fc域和结合区,其中第二变体包括免疫球蛋白的第二fc域和结合区,并且其中
[0695]
(i)所述第一变体包括对应于人igg1重链fc区的k439位置的突变,和所述第二变体包括对应于人igg1重链fc区的s440位置的突变,条件是s440的突变不是s440y或s440w,
[0696]
(ii)所述第一变体包括对应于人igg1重链fc区的k447d/e位置的突变;和所述第二变体包括对应于人igg1重链fc区的k447k/r/h和448p位置的突变,或
[0697]
(iii)所述第一变体包括对应于人igg1重链fc区的k447d/e位置的突变;和所述第二变体包括对应于人igg1重链fc区的k447k/r/h、448k/r/h和449p位置的突变。
[0698]
在一个实施方案中,亲本多肽的第一个或两个变体以及亲本多肽的第二变体可以是抗体。
[0699]
因此在一个方面,本发明涉及一种包括亲本抗体的第一变体和亲本抗体的第二变体的组合物,其中第一变体包括免疫球蛋白的第一fc域和抗原结合区,其中第二变体包括免疫球蛋白的第二fc域和抗原结合区,和其中
[0700]
(i)所述第一变体包括对应于人igg1重链fc区的k439位置的突变,和所述第二变体包括对应于人igg1重链fc区的s440位置的突变,条件是s440的突变不是s440y或s440w,
[0701]
(ii)所述第一变体包括对应于人igg1重链fc区的k447d/e位置的突变;和所述第二变体包括对应于人igg1重链fc区的k447k/r/h和448p位置的突变,或
[0702]
(iii)所述第一变体包括对应于人igg1重链fc区的k447d/e位置的突变;和所述第二变体包括对应于人igg1重链fc区的k447k/r/h、448k/r/h和449p位置的突变。
[0703]
在一个实施方案中,组合物包括亲本抗体的第一变体和亲本抗体的第二变体,其中第一变体包括免疫球蛋白的第一fc域和抗原结合区,其中第二变体包括免疫球蛋白的第二fc域和抗原结合区,以及其中所述第一变体包括对应于人igg1重链fc区的k439位置的突变,和其中第二变体包括对应于s440位置的突变,条件是s440的突变不是s440y或s440w。
[0704]
在一个实施方案中,组合物包括亲本抗体的第一变体和亲本抗体的第二变体,其中第一变体包括免疫球蛋白的第一fc域和抗原结合区,其中所述第一变体包括(i)选自下组的除了k439突变之外的至少一个氨基酸残基的第一突变:
[0705]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0706]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0707]
(c)n端ch3螺旋内的氨基酸残基,
[0708]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0709]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,和
[0710]
(ii)对应于人igg1重链fc区的k439位置的第二突变;和
[0711]
其中第二变体包括免疫球蛋白的第二fc域和抗原结合区,其中所述第二变体包括(i)选自下组的除了s440突变之外的至少一个氨基酸残基的第一突变:
[0712]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0713]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0714]
(c)n端ch3螺旋内的氨基酸残基,
[0715]
(d)c端ch3β链内的氨基酸残基,和
[0716]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,和
[0717]
(ii)在对应于igg1重链fc区的s440位置的第二突变,条件是s440的突变不是s440y或s440w。
[0718]
在另一个实施方案中,组合物包括抗体的第一变体和亲本抗体的第二变体,其中第一变体包括(i)选自下组的除了k439突变之外的至少一个氨基酸残基的第一突变:
[0719]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0720]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0721]
(c)n端ch3螺旋内的氨基酸残基,
[0722]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0723]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,和
[0724]
(ii)对应于人igg1重链fc区的k439位置的第二突变,和
[0725]
其中第二变体包括对应于人igg1重链fc区的s440位置的突变,其不是s440y或s440w。
[0726]
在另一个实施方案中,组合物包括亲本抗体的第一变体和亲本抗体的第二变体,其中第一变体包括对应于人igg1重链fc区的k439位置的突变;和
[0727]
其中第二变体包括免疫球蛋白的第二fc域和抗原结合区,其中所述第二变体包括(i)选自下组的除了s440突变之外的至少一个氨基酸残基的第一突变:
[0728]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0729]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0730]
(c)n端ch3螺旋内的氨基酸残基,
[0731]
(d)c端ch3β链内的氨基酸残基,和
[0732]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,
[0733]
和(ii)在对应于igg1重链fc区的s440位置的第二突变,条件是s440的突变不是s440y或s440w。
[0734]
在一个实施方案中,对应于人igg1重链fc区的k439位置的突变是k439d/e,和/或对应于人igg1重链fc区的s440位置的突变是s440k/h/r。
[0735]
在另一个方面,本发明涉及一种包括包含免疫球蛋白的fc域和结合区的亲本多肽第一变体以及包含免疫球蛋白的fc域和结合区的亲本多肽第二变体的组合物,其中
[0736]
第一变体包括免疫球蛋白的第一fc域和抗原结合区,其中第一变体包括选自以下氨基酸残基的至少一种氨基酸残基的第一突变:
[0737]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0738]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0739]
(c)n端ch3螺旋内的氨基酸残基,
[0740]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0741]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,和其中第二变体不包括选自以下的氨基酸残基的氨基酸残基突变:
[0742]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0743]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0744]
(c)n端ch3螺旋内的氨基酸残基,
[0745]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0746]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基。
[0747]
在一个实施方案中,第一和/或第二亲本多肽可以是抗体。
[0748]
本发明还涉及一种组合物的实施方案,其中第一变体包括(i)选自下组的除了k439突变之外的至少一个氨基酸残基的第一突变:
[0749]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0750]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0751]
(c)n端ch3螺旋内的氨基酸残基,
[0752]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0753]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,和
[0754]
(ii)对应于人igg1重链fc区的k439位置的第二突变,和
[0755]
其中第二变体包括对应于人igg1重链fc区的s440位置的突变,其不是s440y或s440w。
[0756]
本发明还涉及一种组合物的实施方案,其中第一变体包括对应于人igg1重链fc区的k439位置的突变;和
[0757]
其中第二变体包括免疫球蛋白的第二fc域和抗原结合区,其中所述第二变体包括(i)选自下组的除了s440突变之外的至少一个氨基酸残基的第一突变:
[0758]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0759]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0760]
(c)n端ch3螺旋内的氨基酸残基,
[0761]
(d)c端ch3β链内的氨基酸残基,和
[0762]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,和
[0763]
(ii)在对应于igg1重链fc区的s440位置的第二突变,条件是s440的突变不是s440y或s440w。
[0764]
在另一个方面,本发明涉及一种包括抗体的第一变体和亲本抗体的第二变体的组合物,其中第一变体包括免疫球蛋白的第一fc域和第一抗原结合区,其中第一变体包括选自下组的至少一种氨基酸残基的第一突变:
[0765]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0766]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0767]
(c)n端ch3螺旋内的氨基酸残基,
[0768]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0769]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,和
[0770]
其中第二变体包括免疫球蛋白的第二fc域和第二抗原结合区,其中所述第二变体不包括选自下组的氨基酸残基的突变:
[0771]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0772]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0773]
(c)n端ch3螺旋内的氨基酸残基,
[0774]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0775]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基。
[0776]
在其中第二变体不包括本发明所列举的任意突变的实施方案中,这种第二变体可包括涉及效应子作用的方法时上文列举的任意适合第二抗体实例。
[0777]
在一个实施方案中,第一变体和第二变体包括选自对应于以下氨基酸残基的至少一个氨基酸残基的第一突变:人igg1重链fc区中的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436,和k447。
[0778]
在一个实施方案中,第一变体包括选自对应于以下氨基酸残基的至少一个氨基酸残基的突变:人igg1重链fc区中的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447,条件是s440的突变是s440y或s440w。
[0779]
在一个具体实施方案中,第一变体包括对应于人igg1重链fc区的e345r和q386k的氨基酸残基的突变,并且第二变体不包括这种突变。
[0780]
在一个具体实施方案中,第一变体包括对应于人igg1重链fc区的e345r、q386k和e430g的氨基酸残基的突变,并且第二变体不包括这种突变。
[0781]
在一个实施方案中,第一和第二变体中至少一个第一突变是不同的。
[0782]
在一个实施方案中,第一变体和第二变体各自是人igg1,igg2,igg3,igg4,iga1,iga2,igd或ige抗体,任选的各自是人全长抗体,如各自是人全长igg1抗体。
[0783]
在一个实施方案中,第一变体和第二变体选自单特异性抗体,双特异性抗体或多特异性抗体。
[0784]
在进一步的实施方案中,第一和第二变体结合相同抗原或不同抗原上的不同表位。因此,在其中第一和第二抗体是双特异性抗体的实施方案中,它们可以各自结合两个不同表位。至少两种双特异性抗体可以是相同或不同的。如果双特异性抗体是不同的,则组合物包括针对相同或不同靶标上最多4种不同表位。
[0785]
在进一步的实施方案中,第一变体和第二变体中的一种或两种与药物、毒素或放射性标记缀合,例如其中第一变体和第二变体中的一种或两种与毒素通过接头缀合。
[0786]
在进一步的实施方案中,第一变体和第二变体中的一种或两种是融合蛋白的一部分。
[0787]
在另一个方面,本发明涉及一种包括本发明所述任意变体、任意双特异性抗体或任意组合物和药学上可接受的载体的组合物。
[0788]
预期根据“混合突变体”方面的任意实施方案也包括在任意组合物实施方案中。
[0789]
在一个实施方案中,第一和第二亲本抗体的变体结合在相同细胞上表达的抗原。
[0790]
在另一个实施方案中,第一亲本抗体的变体包括k439到选自e和d的氨基酸的氨基酸取代。
[0791]
在另一个实施方案中,第一亲本抗体的变体的氨基酸取代是k439e。
[0792]
在另一个实施方案中,第二亲本抗体的变体包括s440到选自k、r和h的氨基酸的氨基酸取代。
[0793]
在另一个实施方案中,第二亲本抗体的变体的氨基酸取代是s440k。
[0794]
在可选的实施方案中,第一和/或第二抗体的变体进一步包括选自h310,g385,h433,n434和q438的残基的突变。
[0795]
在进一步可选的实施方案中,第一和/或第二亲本抗体的变体进一步包括选自e345到d,k,n,q,r或w;e382到d,q,k或r;和h433到r的突变。
[0796]
在进一步的实施方案中,第一和第二亲本抗体的变体进一步包括选自e345r、e382r和h433r的突变,例如e345r。
[0797]
在另一个方面,本发明涉及一种药物组合物,其包括上文所列任一实施方案的第一亲本抗体的变体和第二亲本抗体的变体。
[0798]
药物组合物可以根据常规技术进行配制,例如公开于remington:the science and practice of pharmacy,第19版,gennaro,ed.,mack publishing co.,easton,pa,1995。本发明的药物组合物可以例如包括稀释剂,填充剂,盐,缓冲剂,去污剂(例如,非离子型洗涤剂,如tween-20或tween-80),稳定剂(例如,糖或无蛋白氨基酸),防腐剂,等渗剂,抗氧化剂,组织固定剂,增溶剂,和/或适于包括在药物组合物中的其他材料。可用于本发明药物组合物的合适水性和非水性载体的实例包括水,盐水,磷酸缓冲盐水,乙醇,葡萄糖,多元醇(例如甘油,丙二醇,聚乙二醇)。
[0799]
可以通过任意合适的途径和方式施用药物组合物。在一个实施方案中,肠胃外施用本发明的药物组合物。本发明使用的术语“肠胃外施用”表示除了肠和局部施用以外的施
用方式,通常通过注射,并且包括表皮,静脉内,肌内,动脉内,鞘内,囊内,眶内,心内,皮内,腹腔内,腱内,经气管,皮下,表皮下,关节内,包囊下,蛛网膜下,脊柱内,颅内,胸内,硬膜外和胸骨内的注射和输注。
[0800]
套组
[0801]
应理解本发明中所有实施方案参考亲本抗体进行描述,还应将第一亲本抗体或第二亲本抗体理解为有关包括免疫球蛋白和结合区的亲本、第一亲本或第二亲本多肽的实施方案。
[0802]
本发明还涉及用于治疗时同时、分开或连续使用的包括变体和亲本抗体的套组,其中任意变体和亲本抗体可以如本发明所述。具体方面和实施方案将在下文描述。此外,可以根据本发明描述的任意方法获得这种变体。
[0803]
本发明还涉及用于治疗时同时、分开或连续使用的包括变体和亲本抗体的套组,它们可以是如本发明所述的任意变体和亲本抗体。具体方面和实施方案将在下文描述。此外,可以根据本发明描述的任意方法获得这种变体。
[0804]
在一个方面,本发明涉及一种用于治疗时同时、分开或连续使用的包括亲本多肽的第一变体和亲本多肽的第二变体的套组,其中第一变体包括免疫球蛋白的第一fc域和结合区,其中第二变体包括免疫球蛋白的第二fc域和结合区,和其中
[0805]
(i)所述第一变体包括对应于人igg1重链fc区的k439位置的突变,和所述第二变体包括对应于人igg1重链fc区的s440位置的突变,条件是s440的突变不是s440y或s440w,
[0806]
(ii)所述第一变体包括对应于人igg1重链fc区的k447d/e位置的突变;和所述第二变体包括对应于人igg1重链fc区的k447k/r/h和448p位置的突变,或
[0807]
(iii)所述第一变体包括对应于人igg1重链fc区的k447d/e位置的突变;和所述第二变体包括对应于人igg1重链fc区的k447k/r/h、448k/r/h和449p位置的突变。
[0808]
在一个实施方案中,亲本多肽的第一个或两个变体以及亲本多肽的第二变体可以是抗体。
[0809]
因此在一个方面,本发明涉及一种用于治疗时同时、分开或连续使用的包括亲本抗体的第一变体和亲本抗体的第二变体的套组,其中第一变体包括免疫球蛋白的第一fc域和抗原结合区,其中第二变体包括免疫球蛋白的第二fc域和抗原结合区,和其中
[0810]
(i)所述第一变体包括对应于人igg1重链fc区的k439位置的突变,和所述第二变体包括对应于人igg1重链fc区的s440位置的突变,条件是s440的突变不是s440y或s440w,
[0811]
(ii)所述第一变体包括对应于人igg1重链fc区的k447d/e位置的突变;和所述第二变体包括对应于人igg1重链fc区的k447k/r/h和448p位置的突变,或
[0812]
(iii)所述第一变体包括对应于人igg1重链fc区的k447d/e位置的突变;和所述第二变体包括对应于人igg1重链fc区的k447k/r/h、448k/r/h和449p位置的突变。
[0813]
在一个实施方案中,用于治疗时同时、分开或连续使用的套组,包括亲本抗体的第一变体和亲本抗体的第二变体,其中第一变体包括免疫球蛋白的第一fc域和抗原结合区,其中第二变体包括免疫球蛋白的第二fc域和抗原结合区,和其中所述第一变体包括对应于人igg1重链fc区的k439位置的突变,和其中第二变体包括对应于s440位置的突变,条件是s440的突变不是s440y或s440w。
[0814]
在一个实施方案中,用于治疗时同时、分开或连续使用的套组,包括亲本抗体的第
一变体和亲本抗体的第二变体,
[0815]
其中第一变体包括免疫球蛋白的第一fc域和抗原结合区,其中所述第一变体包括(i)选自下组的除了k439突变之外的至少一个氨基酸残基的第一突变:
[0816]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0817]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0818]
(c)n端ch3螺旋内的氨基酸残基,
[0819]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0820]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,和
[0821]
(ii)对应于人igg1重链fc区的k439位置的第二突变;和
[0822]
其中第二变体包括免疫球蛋白的第二fc域和抗原结合区,其中所述第二变体包括(i)选自下组的除了s440突变之外的至少一个氨基酸残基的第一突变:
[0823]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0824]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0825]
(c)n端ch3螺旋内的氨基酸残基,
[0826]
(d)c端ch3β链内的氨基酸残基,和
[0827]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,和
[0828]
(ii)在对应于igg1重链fc区的s440位置的第二突变,条件是s440的突变不是s440y或s440w。
[0829]
在另一个实施方案中,用于治疗时同时、分开或连续使用的套组,包括抗体的第一变体和亲本抗体的第二变体,其中第一变体包括(i)选自下组的除了k439突变之外的至少一个氨基酸残基的第一突变:
[0830]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0831]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0832]
(c)n端ch3螺旋内的氨基酸残基,
[0833]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0834]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,和
[0835]
(ii)对应于人igg1重链fc区的k439位置的第二突变,和
[0836]
其中第二变体包括对应于人igg1重链fc区的s440位置的突变,其不是s440y或s440w。
[0837]
在另一个实施方案中,用于治疗时同时、分开或连续使用的套组,包括亲本抗体的第一变体和亲本抗体的第二变体,
[0838]
其中第一变体包括对应于人igg1重链fc区的k439位置的突变;和
[0839]
其中第二变体包括免疫球蛋白的第二fc域和抗原结合区,其中所述第二变体包括(i)选自下组的除了s440突变之外的至少一个氨基酸残基的第一突变:
[0840]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0841]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0842]
(c)n端ch3螺旋内的氨基酸残基,
[0843]
(d)c端ch3β链内的氨基酸残基,和
[0844]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,
[0845]
和(ii)在对应于igg1重链fc区的s440位置的第二突变,条件是s440的突变不是s440y或s440w。
[0846]
在一个实施方案中,对应于人igg1重链fc区的k439位置的突变是k439d/e,和/或对应于人igg1重链fc区的s440位置的突变是s440k/h/r。
[0847]
在另一个方面,本发明涉及一种用于治疗时同时、分开或连续使用的包括亲本多肽的第一变体和亲本多肽的第二变体的套组,其中第一变体包括免疫球蛋白的fc域和结合区,和第二变体包括免疫球蛋白的fc域和结合区,其中
[0848]
第一变体包括免疫球蛋白的第一fc域和抗原结合区,其中第一变体包括选自下组的至少一种氨基酸残基的第一突变:
[0849]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0850]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0851]
(c)n端ch3螺旋内的氨基酸残基,
[0852]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0853]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,和其中
[0854]
第二变体不包括选自以下的氨基酸残基组的突变:
[0855]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0856]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0857]
(c)n端ch3螺旋内的氨基酸残基,
[0858]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0859]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基。
[0860]
在一个实施方案中,第一和/或第二亲本多肽可以是抗体。
[0861]
本发明还涉及一个用于治疗时同时、分开或连续使用的套组的实施方案,其中第一变体包括(i)选自下组的除了k439突变之外的至少一个氨基酸残基的第一突变:
[0862]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0863]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0864]
(c)n端ch3螺旋内的氨基酸残基,
[0865]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0866]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,和
[0867]
(ii)对应于人igg1重链fc区的k439位置的第二突变,和
[0868]
其中第二变体包括对应于人igg1重链fc区的s440位置的突变,其不是s440y或s440w。
[0869]
本发明还涉及一个用于治疗时同时、分开或连续使用的套组的实施方案,其中第一变体包括对应于人igg1重链fc区的k439位置的突变;和
[0870]
其中第二变体包括免疫球蛋白的第二fc域和抗原结合区,其中所述第二变体包括
(i)选自下组的除了s440突变之外的至少一个氨基酸残基的第一突变:
[0871]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0872]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0873]
(c)n端ch3螺旋内的氨基酸残基,
[0874]
(d)c端ch3β链内的氨基酸残基,和
[0875]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,
[0876]
和(ii)在对应于igg1重链fc区的s440位置的第二突变,条件是s440的突变不是s440y或s440w。
[0877]
在另一个方面,本发明涉及一种用于治疗时同时、分开或连续使用的套组,包括抗体的第一变体和亲本抗体的第二变体,其中
[0878]
第一变体包括免疫球蛋白的第一fc域和抗原结合区,其中第一变体包括选自下组的至少一种氨基酸残基的第一突变:
[0879]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0880]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0881]
(c)n端ch3螺旋内的氨基酸残基,
[0882]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0883]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基,和
[0884]
其中第二变体包括免疫球蛋白的第二fc域和第二抗原结合区,其中所述第二变体不包括选自下组的氨基酸残基的突变:
[0885]
(a)在ch2-ch3区内提供异构突变的氨基酸残基,
[0886]
(b)在ch2-ch3区的疏水钮内的氨基酸残基,
[0887]
(c)n端ch3螺旋内的氨基酸残基,
[0888]
(d)c端ch3β链内的氨基酸残基,条件是在对应于人igg1重链fc区中s440的突变情况下,所述突变是s440y或s440w,和
[0889]
(e)对应于人igg1重链fc区中e345、e382或q386的氨基酸残基。
[0890]
在其中第二变体不包括本发明所列举的任意突变的实施方案中,这种第二变体可能包括涉及效应子作用的方法时上文列举的任意适合第二抗体实例。
[0891]
在一个实施方案中,第一变体和第二变体包括选自对应于以下氨基酸残基的至少一个氨基酸残基的第一突变:人igg1重链fc区中的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447。
[0892]
在一个实施方案中,第一变体包括选自对应于以下氨基酸残基的至少一个氨基酸残基的突变:人igg1重链fc区中的e345、e430、s440、q386、p247、i253、s254、q311、d/e356、t359、e382、y436和k447,条件是s440的突变是s440y或s440w。
[0893]
在一个具体实施方案中,第一变体包括对应于人igg1重链fc区的e345r和q386k的氨基酸残基的突变,并且第二变体不包括这种突变。
[0894]
在一个具体实施方案中,第一变体包括对应于人igg1重链fc区的e345r、q386k和e430g的氨基酸残基的突变,并且第二变体不包括这种突变。
[0895]
在一个实施方案中,第一和第二变体中至少一个第一突变是不同的。
[0896]
在一个实施方案中,第一变体和第二变体各自是人igg1,igg2,igg3,igg4,iga1,iga2,igd或ige抗体,任选的各自是人全长抗体,例如各自是人全长igg1抗体。
[0897]
在一个实施方案中,第一变体和第二变体选自单特异性抗体,双特异性抗体或多特异性抗体。
[0898]
在进一步的实施方案中,第一和第二变体结合相同抗原或不同抗原上的不同表位。因此,在其中第一和第二抗体是双特异性抗体的实施方案中,它们可以各自结合两个不同表位。至少两种双特异性抗体可以是相同或不同的。如果双特异性抗体是不同的,则用于治疗时同时、分开或连续使用的套组包括靶向相同或不同靶标上的最多4个不同表位。
[0899]
在进一步的实施方案中,第一变体和第二变体中的一种或两种与药物、毒素或放射性标记缀合,例如其中第一变体和第二变体中的一种或两种与毒素通过接头缀合。
[0900]
在进一步的实施方案中,第一变体和第二变体中的一种或两种是融合蛋白的一部分。
[0901]
预期根据“混合突变体”方面的任意实施方案可以包括在用于治疗时同时、分开或连续使用的任意套组实施方案中。
[0902]
在一个实施方案中,第一和第二亲本抗体的变体结合在相同细胞上表达的抗原。
[0903]
在另一个实施方案中,第一亲本抗体的变体包括k439到选自e和d的氨基酸的氨基酸取代。
[0904]
在另一个实施方案中,第一亲本抗体的变体的氨基酸取代是k439e。
[0905]
在另一个实施方案中,第二亲本抗体的变体包括s440到选自k、r和h的氨基酸的氨基酸取代。
[0906]
在另一个实施方案中,第二亲本抗体的变体的氨基酸取代是s440k。
[0907]
在可选的实施方案中,第一和/或第二抗体的变体进一步包括选自h310,g385,h433,n434和q438的残基的突变。
[0908]
在进一步可选的实施方案中,第一和/或第二亲本抗体的变体进一步包括选自e345到d,k,n,q,r或w;e382到d,q,k或r;和h433到r的突变。
[0909]
在进一步的实施方案中,第一和第二亲本抗体的变体进一步包括选自e345r、e382r和h433r的突变,例如e345r。
[0910]
在另一个方面,本发明涉及一种用于治疗时同时、分开或连续使用的药物套组,包括上文所列任意实施方案的第一亲本抗体的变体和第二亲本抗体的变体。
[0911]
用于治疗时同时、分开或连续使用的药物套组可以通过任意合适的途径和方式进行施用。在一个实施方案中,本发明用于治疗时同时、分开或连续使用的药物套组是肠胃外施用的。本发明使用的术语“肠胃外施用”表示除了肠和局部施用以外的施用方式,通常通过注射,并且包括表皮,静脉内,肌内,动脉内,鞘内,囊内,眶内,心内,皮内,腹腔内,腱内,经气管,皮下,表皮下,关节内,包囊下,蛛网膜下,脊柱内,颅内,胸内,硬膜外和胸骨内的注射和输注。
[0912]
组合
[0913]
应理解本发明中所有实施方案参考亲本抗体进行描述,还应将第一亲本抗体或第二亲本抗体理解为有关包括免疫球蛋白和结合区的亲本、第一亲本或第二亲本多肽的实施方案。
[0914]
此外,本发明提供一种如上所述的任意“单突变体”方面或实施方案的变体的制剂,即,包括多拷贝变体的制剂。本发明还提供包括如上所述的任意“单突变体”方面和实施方案的组合物,例如,药物组合物。本发明还提供任意这种“单突变体”变体、制品或组合物作为药物的用途。
[0915]
本发明还提供变体的组合,其中一种变体包括独立的选自表1所列的至少一种突变,和一种变体包括独立的选自表1所列的至少一种其他突变,以及这种变体组合的制品和药物组合物以及它们作为药物的用途。优选的,两种变体结合通常在相同细胞、细胞膜、病毒粒子和/或其他颗粒的表面上表达的相同抗原或不同抗原。
[0916]
缀合物
[0917]
应理解本发明中所有实施方案参考亲本抗体进行描述,还应将第一亲本抗体或第二亲本抗体理解为有关包括免疫球蛋白和结合区的亲本、第一亲本或第二亲本多肽的实施方案。
[0918]
在一个方面,本发明涉及一种变体,其中所述变体与药物,毒素或放射性标记缀合,例如其中变体与毒素通过接头缀合。
[0919]
在一个实施方案中,所述变体是融合蛋白的一部分。
[0920]
在另一个方面,本发明的变体不在c端与另一分子(例如毒素或标记)缀合。在一个实施方案中,变体与另一分子在另一位点(通常在不干扰寡聚体形成的位点)缀合。例如,抗体变体可以在其他位点与选自毒素(包括放射性同位素)、前药或药物的化合物连接。这种化合物可以使靶细胞的杀伤更有效,例如在癌症治疗中。因此所获变体是免疫缀合物。
[0921]
因此,在进一步的方面,本发明提供一种与一种或更多种治疗部分(例如细胞毒素,化疗药物,细胞因子,免疫抑制剂和/或放射性同位素)连接或缀合的抗体。这种缀合物在本发明中被称为“免疫缀合物”或“药物缀合物”。包括一种或更多种细胞毒素的免疫缀合物被称作“免疫毒素”。
[0922]
细胞毒素或细胞毒剂包括对细胞有害的(例如,杀伤)的任意药剂。用于形成本发明免疫缀合物的合适治疗剂包括红豆杉醇,细胞松弛素b,短杆菌肽d,溴乙锭,吐根碱,丝裂霉素,鬼臼亚乙苷,替尼泊甙,长春新碱,长春碱,秋水仙碱,多柔比星,道诺红菌素,二羟基炭疽菌素二酮,美登素或其类似物或衍生物,enediyene抗肿瘤抗生素包括新制癌菌素在内,刺孢霉素,埃斯波霉素,蒽环类抗生素,lidamycin,kedarcidin或其类似物或衍生物,anthracyclins,米托蒽醌,光辉霉素,放线菌素d,1-去氢睾酮,糖皮质激素,普鲁卡因,丁卡因,利多卡因,普萘洛尔,和嘌呤霉素,抗代谢物(例如氨甲蝶呤,6-巯基嘌呤,6-硫代鸟嘌呤,阿糖胞苷,fludarabin,5-氟尿嘧啶,氨烯咪胺,羟基脲,天冬酰胺酶,吉西他滨,克拉屈滨),烷化剂(例如氮芥,thioepa,苯丁酸氮芥,苯丙氨酸氮芥,亚硝脲氮芥(bsnu),环己亚硝脲(ccnu),环磷酰胺,白消安,二溴甘露醇,链脲霉素,氮烯唑胺(dtic),甲基苄肼,丝裂霉素c,顺铂和其他铂衍生物,例如卡铂;以及多卡米星a,多卡米星sa,cc-1065(亦称为rachelmycin),或cc-1065的类似物或衍生物,多拉司他汀,吡咯并[2,1-c][1,4]benzodiazepins(pdbs)或其类似物,抗生素(例如更生霉素(从前的放线菌素),博来霉素,道诺红菌素(从前的柔红霉素),多柔比星,伊达比星,光辉霉素,丝裂霉素,米托蒽醌,普卡霉素,氨茴霉素(amc)),抗有丝分裂的药剂(例如,微管蛋白抑制剂)例如一甲基auristatin e,一甲基auristatinf,或多拉司他汀10的其他类似物或衍生物;组蛋白去乙酰化酶抑制剂
例如异羟肟酸曲古抑菌素a,伏力诺他(saha),贝林司他,laq824,和帕比司他以及苯甲酰胺,恩替司他,ci994,莫西司他和脂肪族酸化合物例如丁酸苯酯和丙戊酸,蛋白酶体抑制剂例如danoprevir,硼替佐米,毒伞肽例如α-amantin,白喉毒素和相关分子(例如白喉a链及其活性片段和杂交分子);蓖麻毒毒素(例如蓖麻毒a或去糖基化蓖麻毒a链毒素),霍乱毒素,志贺样毒素(slt-i,slt-ii,slt-iiv),lt毒素,c3毒素,志贺毒素,百日咳毒素,破伤风毒素,大豆bowman-birk蛋白酶抑制剂,假单胞菌属外毒素,alorin,皂草素,蒴莲根毒素,gelanin,相思豆毒素a链,蒴莲根毒素a链,α-sarcin,油桐蛋白,石竹素蛋白,美国商陆蛋白(papi,papii,和pap-s),苦瓜抑制剂,泻果素,巴豆毒素,sapaonaria officinalis抑制剂,白树毒素,mitogellin,局限曲菌素,酚霉素,和伊诺霉素毒素。其他合适的缀合分子包括抗菌/裂解肽例如clip,爪蟾抗菌肽2,蜂毒素,杀菌肽,和p18;核糖核酸酶(rnase),dnase i,葡萄球菌肠毒素-a种,美洲商陆抗病毒蛋白,白喉毒素,和假单胞菌属内毒素。参见,例如,pastan等,cell 47,641(1986)和goldenberg,calif.a cancer journal for clinicians 44,43(1994)。可以与本发明别处描述的本发明抗体组合施用的治疗剂(如,例如,抗癌细胞因子或趋化因子)也可以是可用于缀合本发明抗体的治疗性部分的候选物。
[0923]
在一个实施方案中,本发明的药物缀合物包括本发明公开的与auristatins或auristatin肽类似物和衍生物缀合的抗体(us5635483;us5780588)。auristatins已经显示干扰微管动力学,gtp水解以及核和细胞分裂(woyke等(2001)antimicrob.agents和chemother.45(12):3580-3584)并具有抗癌(us5663149)和抗霉菌活性(pettit等,(1998)antimicrob.agents和chemother.42:2961-2965。auristatin药物部分可以利用接头,通过肽药物部分的n(氨基)端或c(端)粘附到抗体。
[0924]
示例性auristatin实施方案包括n端连接的一甲基auristatin药物部分de和df,公开于senter等.,proceedings of the american association for cancer research.45卷,摘要编号623,2004年3月28出版并描述于us 2005/0238649。
[0925]
示例性的auristatin实施方案是mmae(一甲基auristatin e)。另一个示例性的auristatin实施方案是mmaf(一甲基auristatin f)。
[0926]
在一个实施方案中,本发明的抗体包括缀合的核酸或核酸结合分子。在这种实施方案中,缀合的核酸是细胞毒核糖核酸酶,反义核酸,抑制性rna分子(例如,sirnas分子)或免疫刺激性核酸(例如,包含免疫刺激性cpg模体的dna分子)。在另一个实施方案中,本发明的抗体与适体或核酶缀合。
[0927]
在一个实施方案中,提供包括一种或更多种放射性标记的氨基酸的抗体。放射性标记的变体可用于诊断和治疗两种目的(与放射性标记分子的缀合是另一个可能的特征)。用于多肽的标记的非限制性实例包括3h,14c,15n,35s,90y,99tc和125i,131i,和186re。用于制备放射性标记的氨基酸和相关肽衍生物的方法是本领域已知的(参见,例如,junghans等,cancer chemotherapy and biotherapy 655-686(第二版.,chafner和longo,eds.,lippincott raven(1996))和u.s.4,681,581,u.s.4,735,210,u.s.5,101,827,u.s.5,102,990(us re35,500),u.s.5,648,471和u.s.5,697,902。例如,可以通过氯胺t法缀合放射性同位素。
[0928]
在一个实施方案中,本发明的变体与放射性同位素或包含放射性同位素的螯合物缀合。例如,变体可以与螯合剂接头例如dota、dtpa或tiuxetan缀合,这其允许抗体与放射
性同位素形成复合物。变体也可以或可选的包括一种或更多种放射性标记的氨基酸或其他放射性标记的分子,或与一种或更多种放射性标记的氨基酸或其他放射性标记的分子缀合。放射性标记的变体可用于诊断和治疗两种目的。在一个实施方案中,本发明的变体与α发射体缀合。放射性同位素的非限制性实例包括
3
h,
14
c,
15
n,
35
s,
90
y,
99
tc,
125
i,
111
in,
131
i,
186
re,
213
bs,
225
ac和
227
th。
[0929]
在一个实施方案中,本发明的变体可以与选自以下组的细胞因子缀合:il-2,il-4,il-6,il-7,il-10,il-12,il-13,il-15,il-18,il-23,il-24,il-27,il-28a,il-28b,il-29,kgf,ifna,ifn,ifny,gm-csf,cd40l,flt3配体,干细胞因子,安塞司亭和tnfα。
[0930]
本发明的变体也可以通过共价缀合到多聚体进行化学修饰以例如增加它们的循环半衰期。示例性的多聚体,以及将它们粘附到肽的方法,显示于例如us 4,766,106,us 4,179,337,us 4,495,285和us 4,609,546。其他的多聚体包括聚氧乙烯多元醇和聚乙二醇(peg)(例如,分子量为约1,000-约40,000,例如约2,000-约20,000的peg)。
[0931]
可以使用将本发明的变体缀合到缀合分子的本领域已知的任何方法,例如如上所述的那些,包括以下描述的方法:hunter等,nature 144,945(1962),david等,biochemistry 13,1014(1974),pain等,j.immunol.meth.40,219(1981)和nygren,j.histochem.and cytochem.30,407(1982)。可以通过将其他部分化学缀合到变体或其片段的n端侧面或c端侧面(例如,抗体h或l链)来产生这种变体(参见,例如,antibody engineering handbook,osamu kanemitsu编辑,chijinshokan出版(1994))。合适情况下,通过在内部残基或糖进行缀合也可以产生这种缀合的变体衍生物。
[0932]
药剂可以直接或间接缀合到本发明的变体。第二药剂的间接缀合的实例是通过间隔物或接头部分缀合到双特异性抗体的半胱氨酸或赖氨酸残基。在一个实施方案中,将变体通过间隔物或接头缀合到前药分子,所述前药分子可以在体内被活化为治疗药物。在一些实施方案中,接头在胞内条件下是可切割的,这样的话接头的切割在胞内环境中从抗体释放药物单位。在一些实施方案中,接头是通过可切割药剂来切割的,所述可切割药剂存在于胞内环境(例如,在溶酶体或内体或小窝内)。例如,间隔物或接头是通过肿瘤细胞相关酶或其他肿瘤特定条件来切割的,借此形成活性药物。这种前药技术和接头的实例描述于syntarga bv等的wo02083180,wo2004043493,wo2007018431,wo2007089149,wo2009017394和wo201062171。合适的抗体前药技术和多卡米星类似物还可以见于美国专利号6,989,452(medarex),此处引入作为参考。接头还可以或可选的是,例如肽基接头,其被胞内肽酶或蛋白酶切割,包括但不限于,溶酶体或胞内体蛋白酶。在一些实施方案中,肽基接头的长度为至少两个氨基酸或至少三个氨基酸。切割药剂可以包括组织蛋白酶b和d以及胞浆素,已知其都能水解二肽药物衍生物,导致靶细胞内活性药物的释放(参见例如dubowchik和walker,1999,pharm.therapeutics 83:67-123)。在一个具体实施方案中,可被胞内蛋白酶切割的肽基接头是val-cit(缬氨酸-瓜氨酸)接头或phe-lys(苯丙氨酸-赖氨酸)接头(参见例如us6214345,其描述了具有val-cit接头的多柔比星的合成以及phe-lys接头的不同实例)。val-cit和phe-lys接头结构的实例包括但不限于下文描述的mc-vc-pab,mc-vc-gaba,mc-phe-lys-pab或mc-phe-lys-gaba,其中mc是马来酰亚胺基己酰基的缩写,vc是val-cit的缩写,pab是p-氨基苄基氨基甲酸叔丁酯和gaba是γ-氨基丁酸的缩写。使用胞内蛋白水解释放治疗剂的优点是,药剂在缀合时通常是毒性降低的并且缀合物的血清稳定性通常较
高。
[0933]
仍在另一个实施方案中,接头单位是不可切割的,并且药物通过抗体降解来释放(参见us 2005/0238649)。通常,这种接头实质上不对胞外环境敏感。在接头的背景下,如本发明使用的“实质上不对胞外环境敏感”表示当变体抗体药物缀合化合物存在于胞外环境(例如血浆)时,变体抗体药物缀合化合物的样品中不超过20%,通常不超过约15%,更通常不超过约10%,和甚至更通常不超过约5%,不超过约3%,或不超过约1%的接头被切割。通过将变体抗体药物缀合化合物与血浆温育预定时间段(例如2,4,8,16或24小时)然后定量血浆中存在的游离药物量,可以确定接头是否实质上对胞外环境敏感。包括mmae或mmaf和各种接头组分的示例性实施方案具有下列结构(其中ab表示抗体,和p表示药物载量(或每个抗体分子的细胞生长抑制或细胞毒药物的平均数)是1到约8,例如p可为4-6,如3-5,或p可为1,2,3,4,5,6,7或8)。
[0934]
其中可切割接头与auristatin组合的实例包括mc-vc-pab-mmaf(也称为vcmmaf)和mc-vc-pab-mmaf(也称为vcmmae),其中mc是马来酰亚胺基己酰基的缩写,vc是基于val-cit(缬氨酸-瓜氨酸)接头的缩写,和pab是对-氨基苄基氨基甲酸叔丁酯的缩写。
[0935]
其他实例包括与不可切割接头组合的auristatins,例如mcmmaf(mc(mc与本上下文的mc相同)是马来酰亚胺基己酰基的缩写。
[0936]
在一个实施方案中,药物接头部分是vcmmae。vcmmae药物接头部分和缀合方法公开于wo2004010957,us7659241,us7829531,us7851437和us11/833,028(seattle genetics,inc.),(其在此引入作为参考),和使用类似于其中公开的方法的方法将vcmmae药物接头部分与抗体在半胱氨酸处结合。
[0937]
在一个实施方案中,药物接头部分是mcmmaf。mcmmaf药物接头部分和缀合方法公开于us7498298,us 11/833,954和wo2005081711(seattle genetics,inc.),(其在此引入作为参考),和使用类似于其中公开的方法将mcmmaf药物接头部分与变体在半胱氨酸处结合。
[0938]
在一个实施方案中,本发明的变体连接到螯合剂接头,例如tiuxetan,其允许双特异性抗体缀合到放射性同位素。
[0939]
在一个实施方案中,变体的每个臂(或fab-臂)与相同的一种或更多种治疗部分直接或间接偶联。
[0940]
在一个实施方案中,只有变体的一个臂与一种或更多种治疗部分直接或间接偶联。
[0941]
在一个实施方案中,变体的每个臂与不同的治疗部分直接或间接偶联。例如,在变体是双特异性抗体并通过两种不同单特异性抗体(例如本发明描述的第一和第二抗体)的受控fab-臂交换来制备的实施方案中,可以通过使用与不同治疗部分缀合或结合的单特异性抗体来获得这种双特异性抗体。
[0942]
进一步的用途
[0943]
应理解本发明中所有实施方案参考亲本抗体进行描述,还应将第一亲本抗体或第二亲本抗体理解为有关包括免疫球蛋白和结合区的亲本、第一亲本或第二亲本多肽的实施方案。在进一步的方面,本发明涉及如上所述的本发明变体作为药物的用途,具体来说作为治疗疾病或病症的药物的用途,其中需要对靶细胞(例如,肿瘤,细菌或真菌细胞)或靶标生
物体(例如,病毒)或细菌或病毒感染细胞的cdc介导杀伤。这种疾病和病症的实例包括,但不限于,癌症和细菌、病毒或真菌感染。
[0944]
在另一个方面,本发明涉及用于治疗疾病例如癌症的本发明所述变体,双特异性抗体,组合物和套组。
[0945]
在另一个方面,本发明涉及一种人的治疗方法,包括施用本发明描述的变体,组合物或套组。
[0946]
在另一个方面,本发明涉及一种治疗人类癌症的方法,包括施用本发明描述的变体,组合物或套组。
[0947]“治疗”是指施用有效量的本发明治疗活性化合物,目的是减轻,改善,阻滞或清除(治愈)症状或疾病状态。
[0948]“有效量”或“治疗有效量”是指能够有效的在剂量方面和一段时间内实现所需治疗结果必需的量。抗体的治疗有效量将根据各种因素而不同,例如个体的疾病状况,年龄,性别和体重,及所述抗体在个体中引发所需应答的能力。治疗有效量还是抗体或抗体部分的治疗有益作用超过任何毒性或不利作用的量。
[0949]
不受理论限制,当将本发明的任意“单突变体”方面或实施方案引入这种治疗活性化合物时,可以降低治疗活性化合物的有效量。
[0950]
用于癌症抗体的合适抗原可以与本发明描述的相同。实施例15到18描述了提供增强和/或更特异性的肿瘤细胞的补体激活或cdc的具体应用。例如,根据“单突变体方面”的包括例如e345r突变的抗肿瘤抗体,可以提供增强的肿瘤细胞的cdc或adcc、adcp应答。此外,在这种方法的变体中,根据“单突变体”方面的突变,例如,例如,e345r,e430,s440或q386,可选的e382或h433r,或表1所列的任意其他突变,可以添加到每种抗体中,从而提供特异性针对表达至少两种抗原的肿瘤细胞的增强cdc和/或adcc应答。
[0951]
用于细菌感染的合适抗体包括,但不限于,靶向金黄色葡萄球菌的那些抗体,例如嵌合单克隆igg1帕昔单抗(bsyx-a110;biosynexus),靶向脂胞壁酸(lta)的那些抗体,所述脂胞壁酸嵌入葡萄球菌属的细胞壁并描述于baker(natbiotechnol.2006dec;24(12):1491-3)和weisman等(int immunopharmacol.2009may;9(5):639-44),这两篇文献均在此完整引入作为参考。实施例14描述了使用包括e345r突变的金黄色葡萄球菌抗体变体的具体实施方案。但是,表1的其他突变,包括但不限于e430g和s440w,可选的e382r和h433r,可以按照类似方式应用以增强抗体抗细菌抗原的cdc介导能力。
[0952]
病毒或真菌感染的合适抗原可以是本发明描述的任一项。
[0953]
在一个实施方案中,变体结合的抗原不是人epha2。在另一个实施方案中,变体不是来源于人epha2 mab 12g3h11(描述于dall'acqua等,上文,其在此完整引入作为参考)。在另一个实施方案中,变体结合的抗原不是il-9。在另一个实施方案中,变体不是来源于wo2007005612(在此完整引入作为参考)描述的fa-hg1或fa-hg4抗体,或其任意变体。在一个实施方案中,变体结合的抗原不是hiv-1gp120。在另一个实施方案中,变体不是来源于针对gp120的b12人igg1κ抗体。
[0954]
在一个具体实施方案中,变体来源于双特异性亲本抗体。双特异性抗体可以具有任意同种型,例如,例如,igg1,igg2,igg3或igg4,并且可以是全长抗体或其包含fc的片段。用于制备双特异性抗体的示例性方法描述于wo2008/119353(genmab)。
[0955]
剂量
[0956]
应理解本发明中所有实施方案参考亲本抗体进行描述,还应将第一亲本抗体或第二亲本抗体理解为有关包括免疫球蛋白和结合区的亲本、第一亲本或第二亲本多肽的实施方案。
[0957]
用于抗体的有效剂量和给药方案取决于待治疗的疾病或病征,并且可以由本领域技术人员来确定。本发明抗体的治疗有效量的示例性、非限制性范围是约0.1到100mg/kg,例如约0.1到50mg/kg,例如约0.1到20mg/kg,例如约0.1到10mg/kg,例如约0.5,约如0.3,约1,约3,约5,或约8mg/kg。
[0958]
本发明的抗体变体还可以与一种或更多种补体因子或相关组分组合施用以增强变体的治疗功效和/或弥补补体消耗。这种补体因子和相关组分包括,但不限于,c1q,c4,c2,c3,c5,c6,c7,c8,c9,mbl和因子b。组合施用可以是同时、分开或连续的。在一个具体实施方案中,本发明提供一种试剂盒,其包括包含本发明变体的药物组合物,以及在相同或不同的药物组合物中的至少一种补体因子或相关组分,以及使用说明书。
[0959]
本发明的抗体变体也可以在组合疗法中施用,即,与待治疗疾病或病征相关的其他治疗剂组合。因此,在一个实施方案中,包含抗体的药物用于与一种或更多种其他治疗剂(例如细胞毒剂、化疗剂或抗血管生成剂)组合。这种组合施用可以是同时、分开或连续的。
[0960]
在进一步的实施方案中,本发明提供一种用于治疗或预防疾病例如癌症的方法,所述方法包括结合放疗和/或外科手术,给需要其的受试者施用治疗有效量的本发明变体或药物组合物。
[0961]
制备方法
[0962]
应理解本发明中所有实施方案参考亲本抗体进行描述,还应将第一亲本抗体或第二亲本抗体理解为有关包括免疫球蛋白和结合区的亲本、第一亲本或第二亲本多肽的实施方案。
[0963]
本发明还提供编码上述任一方面的变体的分离的核酸和载体,以及编码所述变体的载体和表达系统。用于抗体及其变体的合适核酸构建体、载体和表达系统是本领域已知的,并在实施例中描述。在其中变体不仅包括重链(或其包含fc的片段)而且包括轻链的实施方案中,编码重链和轻链部分的核苷酸序列可以存在于相同或不同的核酸或载体。
[0964]
本发明还提供一种在宿主细胞中产生上述任一方面的抗体变体的方法,其中所述变体至少包括重链的fc区,所述方法包括下列步骤∶
[0965]
a)提供编码所述变体的所述fc区的核苷酸构建体,
[0966]
b)在宿主细胞中表达所述核苷酸构建体,
[0967]
和
[0968]
c)从所述宿主细胞的细胞培养物回收所述抗体变体。
[0969]
在一些实施方案中,抗体是重链抗体。但是,在大部分实施方案中,抗体将还将包含轻链,因此所述宿主细胞还表达在相同或不同载体上编码轻链的构建体。
[0970]
适于抗体重组表达的宿主细胞是本领域公知的,包括cho,hek-293,per-c6,ns/0和sp2/0细胞。在一个实施方案中,所述宿主细胞是能够进行蛋白asn-连接糖基化的细胞,例如真核细胞,例如哺乳动物细胞,例如人细胞。在进一步的实施方案中,所述宿主细胞是非人细胞,其被遗传改造以产生具有似人糖基化或人糖基化的糖蛋白。这种细胞的实例是
遗传改造的毕赤酵母(hamilton等,science 301(2003)1244-1246;potgieter等,j.biotechnology 139(2009)318-325)和遗传改造的浮萍(lemna minor)(cox等,nature biotechnology 12(2006)1591-1597)。
[0971]
在一个实施方案中,所述宿主细胞是不能从抗体重链有效去除c端赖氨酸k447残基的宿主细胞。例如,liu等(2008)j pharm sci 97:2426(在此引入作为参考)的表2列出了大量这种抗体生产系统,例如sp2/0、ns/0或转基因乳腺(山羊),其中只获得c端赖氨酸的部分去除。在一个实施方案中,宿主细胞是具有改变的糖基化机制的宿主细胞。本领域已经描述了这种细胞并将其用做宿主细胞,在所述宿主细胞中表达本发明的变体从而产生糖基化改变的抗体。参见,例如,shields,r.l.等(2002)j.biol.chem.277:26733-26740;umana等(1999)nat.biotech.17:176-1,以及ep1176195;wo03/035835;和w099/54342。用于产生改造糖型的其他方法是本领域已知的,并且包括但不限于描述于以下的那些:davies等,2001,biotechnol bioeng 74:288-294;shields等,2002,j biol chem 277:26733-26740;shinkawa等,2003,j biol chem 278:3466-3473),us6602684,wo00/61739a1;wo01/292246a1;wo02/311140a1;wo 02/30954a1;potelligent
tm technology(biowa,inc.princeton,n.j.);glycomab
tm
糖基化工程技术(glycart biotechnology ag,zurich,switzerland);us 20030115614;okazaki等,2004,jmb,336:1239-49。
[0972]
本发明还涉及通过如上所述的本发明方法获得的或可获得的抗体。
[0973]
在进一步的方面,本发明涉及一种能够产生本发明抗体变体的宿主细胞。在一个实施方案中,已经用本发明的核苷酸构建体转化或转染宿主细胞。
[0974]
通过下列实施例进一步说明本发明,其不应被理解为进一步的限制。
实施例
[0975]
实施例1
[0976]
7d8突变体的设计和产生
[0977]
将人单克隆抗体humab-7d8(描述于wo2004/035607中)用作模式抗体。其属于人抗-cd20 igg1抗体组,包括奥法木单抗(humax-cd20,2f2)。这些抗体靶向cd20分子上独特的近膜抗原表位并显示出强烈的cdc。
[0978]
为了测试寡聚fc-fc相互作用在补体激活和cdc中的功能相关性,对位于fc:fc界面的疏水片中的氨基酸进行突变,以潜在地破坏7d8的fc-fc侧向相互作用和cdc效力。在第一组突变体中(表3),在基于1hzh晶体结构而被选择以及被发现暴露在ch2-ch3域疏水片中的位置上引入突变以改变电荷(burton mol immunol 1985 3月;22(3):161-206)。
[0979]
从第一组突变体中发现,i253d和h433a对7d8的cdc丢失产生了最为强烈的影响(例如,实施例5)。1hzh晶体结构显示,i253和h433与配对抗体相对fc位置上的两个不同的凹陷(pocket)相结合。基于这些数据,在晶体结构中的i253和h433位点周围合成了第二组突变以进一步研究位于fc:fc侧向界面的残基对于cdc的重要性。可潜在地破坏fc:fc界面稳定性并进而破坏cdc稳定性的位于i253和h433位点周围的第二组突变列于表4中。
[0980]
为了排除所观察到的对于cdc的影响是由于对c1q直接结合位点的破坏的这种可能性,基于两个显示出cdc丢失的单独突变体产生了双突变体,以测试其恢复由单突变所造成的cdc丢失的能力。该原理图示于图1d中。双突变列于表5中,结构示意图展示在图4和图5
中。
[0981]
施用quikchange定点诱变试剂盒(stratagene,美国)来制备突变体。简言之,使用编码所期望突变的正向引物和反向引物来复制编码igg1m(f)同种异型的7d8重链的全长质粒dna模板。使用dpni消化所得到的dna混合物以除去原料质粒dna,并将其用来转化大肠杆菌(e.coli)。通过dna测序来验证从所得到的菌落中分离出来的突变质粒dna(agowa,德国)。使用293fectin(invitrogen,美国)基本上如制造商所述将编码抗体重链和轻链两者的质粒dna混合物瞬时转染至freestyle hek293f细胞中(invitrogen,美国)。
[0982]
表3:导入7d8的ch2-ch3域的组1突变
[0983][0984]
(=)无电荷
[0985]
(-)负电荷
[0986]
(+)正电荷
[0987]
(δ+)部分正电荷
[0988]
表4:导入7d8的ch2-ch3域的组2突变
[0989][0990]
(=)无电荷
[0991]
(-)负电荷
[0992]
(+)正电荷
[0993]
(δ+)部分正电荷
[0994]
表5:双突变导入7d8的ch2-ch3域来组合各自显示cdc
[0995]
缺失的两个单突变
[0996][0997]
(=)无电荷
[0998]
(-)负电荷
[0999]
(+)正电荷
[1000]
实施例2
[1001]
7d8突变体在细胞上对cd20的结合
[1002]
通过facs分析来分析纯化的抗体样本与cd20阳性细胞的结合。在daudi细胞上测试第一组突变(表3),在raji细胞上测试第二组突变(表4)。在聚苯乙烯96孔圆底平板中(greiner bio-one 650101)将10
5
个细胞温育于具有系列稀释的抗体制剂(对于daudi上的第一组,范围从0.04至10μg/ml以3倍稀释,对于raji上的第二组,范围从0.003至10μg/ml以3倍稀释)的50μl rpm1640/0.1%bsa中,在4℃30分钟。在rpm1640/0.1%bsa中洗涤两次之后,将细胞温育于具有二抗的100μl中,在4℃30分钟。对于daudi细胞上的所有实验以及
raji细胞上使用7d8抗体的实验,都使用异硫氰酸荧光素(fitc)连接的兔-抗-人igg(f0056,dako,glostrup,丹麦;1/100)作为二抗。对于raji细胞上使用纯化的7d8进行的实验,则使用r-藻红蛋白(r-pe)-连接的山羊f(ab
’
)2抗-人κ轻链(2062-09,southernbiotech;1/500)作为二抗。接着,在pbs/0.1%bsa/0.02%叠氮物中洗涤细胞两次,重悬于100μl pbs/0.1%bsa/0.02%叠氮物中,并在facs cantoll(bd biosciences)上进行分析。使用graphpad prism v5.01软件(graphpad software,san diego,ca,美国)利用非线性回归来分析结合曲线。
[1003]
7d8抗体对daudi细胞的结合并没有受到在ch2-ch3域引入点突变的影响,对于所有测试的突变体和野生型7d8都是相同的。另外,相比起野生型7d8,7d8抗体对raji细胞的结合没有明显地受到在ch2-ch3域引入点突变的影响,除了e345r。在cd20-阳性raji细胞上检测到igg1-7d8-e345r在0.3μg/ml以上的浓度时结合减少。也检测到h433d和h433r在所测试的最高抗体浓度时(10μg/ml)的结合也减少。igg1-7d8-e345r、h433d和h433r的结合减少可以被二抗表位的遮掩所解释,这是由于e345r和h433r的直接标记导致相似或者甚至提高的对daudi细胞的结合。这种亲合力的提高可以通过相比起野生型igg1-7d8 fc-fc侧向结合被e345r和h433r所提高来解释。
[1004]
组合k439e和s440k突变并没有影响7d8抗体对raji细胞的结合,其与单突变和野生型7d8的结合是相同的。
[1005]
实施例3
[1006]
7d8突变体对c1q的结合elisa
[1007]
在elisa中测试7d8突变体结合c1q,其中经纯化的抗体涂覆在塑料表面上,使得随机的抗体多聚化。汇集的人血清用作c1q的来源。
[1008]
用系列稀释于pbs中的抗体(范围0.58-10.0μg/ml以1.5-倍稀释)涂覆96孔microlon elisa板(greiner,德国),在4℃过夜。用200μl/孔的补充有0.025%吐温20和0.1%白明胶的0.5
×
pbs洗涤和封闭平板。平板依次用3%汇集的人血清(sanquin,货号#m0008)在37℃温育1小时,用100μl/孔的兔抗人c1q(dako,货号#a0136,1/4.000)在室温下温育1小时,以及用100μl/孔的猪抗兔igg-hrp(dako,p0399,1∶10000)作为检测抗体在室温下1小时,并在温育之间进行洗涤。用1mg/ml的2,2
’-
二氮-双(3-乙基苯并噻唑-6-磺酸)(abts;roche,mannheim,德国)显影约30分钟。通过加入100μl 2%草酸终止反应。在微板读数器(biotek,winooski,vt)中测量405nm处的吸光度。通过使用graphpad prism软件利用可变斜率拟合s型剂量响应曲线来分析对数转换数据。突变体的ec
50
值对于每个平板针对野生型igg1-7d8进行归一化并将其乘以所有野生型igg1-7d8数据的平均值。
[1009]
如图6和表6中所示,如elisa中测量的所测试的点突变对于c1q结合具有最小的影响。对于igg1-7d8-i253d突变体,在elisa中测量到了稍低效率的c1q结合(更高的ec
50
值)。测量了所有抗体的涂覆效能,并发现所有抗体都是相似的。
[1010]
表6:elisa中c1q结合的ec
50
[1011][1012]
1
从至少3个实验计算平均和sd
[1013]
2
统计:对log转换的数据使用dunnett
’
s multiple comparison test(graphpad prism5.01)单路anova。与野生型igg1-7d8比较计算显著性:(na)不适用,(ns)不显著(*)p=0.01到0.05(**)p=0.001到0.01(***)p<0.001
[1014]
实施例4
[1015]
7d8突变体在细胞上对c1q的结合
[1016]
将抗体涂覆在塑料表面上产生抗体结合和fc-尾部呈递的人造静态系统。因此,在基于细胞的测定中也对补体结合进行测试,其中通过facs分析来测量c1q与抗体调理的cd20-阳性b细胞的结合。在使用组1突变体的实验中,在冰上将daudi或raji细胞重悬于90μl含有10%fbs的rpmi 1640培养基中(2
×
10
6
个细胞/ml)。加入10μl系列浓度的c1q(complement technologies,tyler,tx)(终浓度范围在0-60μg/ml和0-140μg/ml之间变化,取决于最大结合)。然后,加入10μl纯化的抗体(10μg/ml的终浓度,即饱和浓度),并立即将反应混合物转移至37℃水浴中,温育1小时。在使用组2突变体的实验中,将测试mab批量加入到daudi细胞中,然后再向等分试样中加入各种浓度的c1q,如上温育混合物。用pbs/1%bsa洗涤细胞三次,与fitc标记的兔抗c1q抗体(dakocytomation,10ug/ml)室温下温育30分钟。用pbs/1%bsa洗涤细胞并重悬于pbs中或者在溶于pbs2%的甲醛中固定。在facscalibur流式细胞仪(bd biosciences)上进行流式细胞术,使用标准化微球(spherotech)将平均荧光强度转化为可溶性荧光素当量分子(mesf)。使用sigmaplot软件(systat software inc.,washington)来计算c1q与经7d8抗体调理的cd20阳性细胞结合的解离常数(k
d
值)。从重复结合实验(在daudi细胞上进行了4次,在raji细胞中进行了3次)中计算平均k
d
值,并将其与c1q和经野生型7d8抗体调理的细胞结合的k
d
值进行对比(表7和表8)。
[1017]
在daudi和raji细胞上都对组1突变体进行了测试并给出了相同的结果。与c1q elisa结果相反,大多数测试的突变体在抗体调理的daudi(表7a)和raji(表8)细胞上都显示出降低的c1q结合活性(k
d
增加)。与野生型7d8相比,在c1q与经调理的daudi或raji细胞的结合活性上igg1-7d8-q311a与h435a几乎没有显示出降低,i253a、i253y和n434a显示出更为显著地降低,i253d和h433a则是急剧降低。在两种细胞类型上,igg1-7d8-h435r显示出稍微高于野生型7d8的c1q结合亲合力(较低k
d
),然而,这并不显著。
[1018]
在daudi细胞上测试组2突变体。与野生型7d8相比,igg1-7d8-e345r、e382r和h433r在c1q与经调理的daudi细胞的结合活性上显示出提高的结合亲合力,通过更低的k
d
值反应出来(表7b)。与野生型7d8相比,其他所有的组2突变体都显示出降低的结合亲合力,
g385d、y436d、q438d、k439e和s440k显示出急剧增加的k
d
值(表7b)以及h433d和y436c显示出急剧降低的结合以至于没有测量到可信的k
d
值。
[1019]
双突变体igg1-7d8-k439e/s440k在抗体调理的daudi细胞上显示出恢复c1q结合,而与野生型7d8相比,两个单突变体都显示出降低的c1q结合。与野生型7d8相比,k439e/s440k双突变体的结合亲合力甚至略为提高(表7c)。单突变体igg1-7d8-k439e和igg1-7d8-k440e的混合物能够完全恢复c1q结合,其可与野生型7d8的c1q结合相比较(表7c)。
[1020]
elisa中c1q结合没有变化(实施例3)与基于细胞的测定中igg1-7d8突变体影响c1q结合之间相互矛盾,显示所测试的参与抗体分子之间fc:fc相互作用的ch3位点,并没有直接影响c1q结合,但是当结合细胞时,却是影响抗体fc尾部动力学定位并因此也是影响c1q结合强度的重要决定因素。
[1021]
表7a:c1q与抗体调理的daudi细胞结合的k
d
值(突变体组1)
[1022][1023]
*与野生型7d8比较(t-检验)
[1024]
**(na)不适用
[1025]
表7b:c1q与抗体调理的daudi细胞结合的k
d
值(突变体组2)
[1026][1027]
*与野生型7d8比较(t-检验)
[1028]
**(na)不适用
[1029]
***7d8的平均k
d
从实验1,2,3,4,10和11计算
[1030]
****由于这些突变体的结合太弱不能测量可靠的拟合曲线和k
d
值
[1031]
表7c:c1q与抗体调理的daudi细胞结合的k
d
值(双突变体)
[1032][1033]
*与野生型7d8比较(t-检验)
[1034]
**(na)不适用
[1035]
***7d8的平均k
d
从实验1,2,3,4,10和11计算
[1036]
表8:c1q与抗体调理的raji细胞结合的k
d
值(突变体组1)
[1037][1038]
*与野生型7d8比较(t-检验)
[1039]
**(na)不适用
[1040]
实施例5
[1041]
在cd20阳性raji细胞上进行cdc测定中7d8突变体的c1q效力
[1042]
在cdc测定中使用经igg1-7d8突变体调理的细胞测试c1q效力,以研究所观察到的c1q结合亲合力变化对于cdc活性的影响。因此,使用补充有确定浓度系列的c1q的c1q缺乏正常人血清来进行cdc测定。在圆底96孔板中(nunc,rochester,ny),将0.1
×
10
6
个raji细胞与10μg/ml纯化的抗体以及系列浓度的人c1q(0.005、0.025、0.1、0.3、1.0、5.0、30.0μg/ml)在总体积为100μl的补充有0.1%bsa的rpmi1640培养基中室温下预温育15分钟。接着,加入25μl c1q缺乏的血清(quidel,sandiego,ca),在37℃的水浴中温育30分钟或培养箱中温育45分钟。温育之后,将样品置于冰上终止反应。利用碘化丙啶染色活细胞排斥(pi,sigma aldrich,zwijndrecht,荷兰)测定在facs上确定细胞的裂解。裂解百分比(%)如下确定:%裂解=(pi阳性细胞数量/细胞总数)
×
100%。
[1043]
将存在30μg/ml c1q时由野生型7d8进行的裂解减去不加入c1q时的裂解设定为100%。利用graphpad prism软件通过在对数转换数据上拟合s型剂量响应曲线来计算ch
50
值(导致50%裂解时的c1q浓度)。将突变体的ch
50
值针对野生型7d8进行标准化(表9)。
[1044]
表9中的数据显示,与c1q结合亲合力的测量值一致,igg1-7d8-q311a、e382r和h435a在c1q效力上没有显示出降低;i253a、i253y、g385d、n434a和y436c的c1q效力显著降低;i253d、h310k、k322a、h433a、h433d、y436d、q438d、k439e和s440k在所有所测试的c1q浓度下几乎完全丧失诱导cdc的能力。
[1045]
igg1-7d8-h435r和h433r能略微高效地使用c1q,这导致比起野生型7d8更为高效
的cdc。igg1-7d8-e345r在c1q效力上显示出急剧提高,这导致了显著高于野生型7d8的cdc裂解(表9)。
[1046]
图7显示,组合k439e和s440k突变当两个突变都被组合在一个分子中时或当两个单突变体被组合时(k439e+s440k混合)(其作为单突变体都能导致cdc丢失)在c1q效力测定中恢复了cdc。
[1047]
表9:对raji细胞的cdc检测中c1q效能的ch
50
[1048][1049]
(1)
实验数
[1050]
(2)
平均和sd从所有进行的实验计算
[1051]
(3)
统计:对log转换的数据使用dunnett
’
s multiple comparison test(graphpad prism 5.01)单路anova。与野生型igg1-7d8比较计算显著性:(na)不适用,(nd)不确定(ns)不显著(*)p=0.01到0.05(**)p=0.001到0.01(***)p<0.001
[1052]
(4)
当裂解未达到50%时,ch
50
设置到>30ug/ml
[1053]
(5)
对没有达到50%裂解的突变体不能确定p值。然而,假设这些与igg1-7d8-wt显著不同
[1054]
实施例6
[1055]
在cd20阳性细胞上进行cdc测定中7d8突变体的cdc
[1056]
在圆底96孔板中(nunc,rochester,ny),将0.1
×
10
6
个raji细胞与10μg/ml纯化的抗体以及系列浓度的抗体(0.01、0.03、0.1、0.3、1.0、3.0、10.0、30.0μg/ml)在总体积80μl中室温下在摇床中预温育15分钟。接着,加入20μl正常人血清作为c1q来源(20%终浓度),在37℃的培养箱中温育45分钟。加入30μl补充有0.1%bsa的冰冷的rpmi培养基终止反应。通过利用碘化丙啶在facs上确定细胞的裂解。
prism 5.01)单路anova。与野生型-7d8比较计算显著性:(na)不适用,(nd)不确定(ns)不显著(*)p=0.01到0.05(**)p=0.001到0.01(***)p<0.001
[1065]
(4)
当裂解未达到50%时,ec
50
设置到>30ug/ml
[1066]
(5)
对没有达到ec
50
裂解的突变体不能确定p值。然而,假设这些与野生型7d8-wt显著不同
[1067]
表11从对raji细胞的cdc检测计算的ed
50
[1068][1069]
(1)
实验数
[1070]
(2)
平均和sd从所有进行的实验计算
[1071]
(3)
统计:对log转换的数据使用dunnett
’
s multiple comparison test(graphpad prism 5.01)单路anova。与野生型7d8比较计算显著性:(na)不适用,(nd)不确定(ns)不显著(*)p=0.01到0.05(**)p=0.001到0.01(***)p<0.001
[1072]
(4)
当裂解未达到ch
50
时,ch50设置到>30ug/ml
[1073]
(5)
对没有达到ec
50
裂解的突变体不能确定p值。然而,假设这些与野生型7d8-wt显著不同
[1074]
实施例7
[1075]
根据其诱导cdc的能力对7d8突变体的排序
[1076]
对于所测试的7d8突变体,发现daudi细胞上的c1q结合(实施例4中所述)与raji细
胞上的c1q效力测定(实施例5中所述)之间,以及daudi细胞上的c1q结合与daudi和raji细胞上的cdc测定(实施例6中所述)之间存在相关性(相关性数据表13)。因此,使用在daudi细胞上进行的c1q结合测定的k
d
值根据其诱导cdc的能力对所有测试的7d8突变体进行排序,如表12所述。
[1077]
表12:根据c1q与daidi细胞结合的k
d
值的下降,这作为其诱导cdc能力的代表,来排列所有检测的7d8突变体
[1078][1079]
*无可靠的拟合曲线,由于这些突变体结合太弱不能测量斜体的k
d
值
[1080]
表13:c1q与daidi细胞结合(实施例4)和对daudi细胞的c1q效能检测(实施例5)之间,c1q与daidi细胞结合和对daudi及raji细胞的cdc检测(实施例06)之间的关联。关联分析之前数据被log转化
[1081]
[1082]
实施例8
[1083]
cd38抗体005突变体的设计和产生
[1084]
人单克隆抗体humab 005是全人igg1κ抗体,描述于wo2006/099875中。在本文中,将其用作验证所确定的fc突变增强cdc活性的模式抗体。所测试的突变列于表14中。
[1085]
使用具有igg1m(f)同种异型的humab 005的重链作为诱变反应的模板,制备不同突变体的dna构建体,并如实施例1所述进行瞬时转染。
[1086]
表14:导入005的ch2-ch3域的突变组(humax-cd38)
[1087][1088]
(=)无电荷
[1089]
(-)负电荷
[1090]
(+)正电荷
[1091]
(δ+)部分正电荷
[1092]
实施例9
[1093]
humab-005突变体在细胞上对cd38的结合
[1094]
通过facs分析来分析未纯化的抗体样本与cd38阳性daudi和raji细胞的结合。在聚苯乙烯96孔圆底平板中(greiner bio-one 650101)将10
5
个细胞温育于具有系列稀释的抗体制剂(0.01、0.03、0.1、0.3、1.0、3.0、10.0、30.0μg/ml)的100μl rpm1640/0.1%bsa中,在4℃30分钟。在rpm1640/0.1%bsa中洗涤两次之后,将细胞与fitc-连接的兔f(ab
’
)2抗人igg(货号f0056;dako;1∶150)温育于50μl中,在4℃30分钟。接着,在pbs/0.1%bsa/0.02%叠氮物中洗涤细胞两次,重悬于100μl pbs/0.1%bsa/0.02%叠氮物中,并在facscantoll(bd biosciences)上进行分析。使用graphpad prism v5.01软件分析结合曲线。使用模拟转染细胞的悬浮液作为阴性对照。
[1095]
humab 005对daudi细胞的结合并没有太多地受到在ch2-ch3域中引入点突变的影响。所有测试的抗体以剂量-依赖性的方式与daudi细胞结合。所有测试的抗体的结合与野生型humab-005相似,除了005-e345r,其显示出稍微降低的结合。然而,不受任何理论束缚,这种较低的结合可能是由于二抗结合的降低所造成的,与实施例2中的igg1-7d8-e345类似。005-e345的实际结合亲合力可能与005-wt相似或者甚至比005-wt还有所提高,然而由于缺少直接标记的抗体,我们无法对此进行证实。
[1096]
humab-005对raji细胞的结合也没有太多地受到在ch2-ch3域中引入点突变的影响。所有测试的抗体以剂量-依赖性的方式与raji细胞结合。005-i253d和h433a的最大结合与野生型005相似,而005-e435r、k439e、s440k突变体以及005-k439e+005-s440k的组合却比野生型005低。然而,不受任何理论束缚,这种较低的结合可能是由于二抗结合的降低所造成的,与实施例2中的igg1-7d8-e345类似(表位被遮蔽)。
[1097]
实施例10
[1098]
cd38抗体005突变体在cd38阳性细胞上的cdc测定
[1099]
在圆底96孔板中(nunc,rochester,ny),将0.1
×
10
6
个raji细胞与系列浓度的未
纯化抗体(0.01、0.03、0.1、0.3、1.0、3.0、10.0、30.0μg/ml)在总体积100μl中室温下在振荡器中预温育15分钟。接着,加入25μl正常人血清作为c1q来源(20%终浓度),在37℃的培养箱中温育45分钟。将平板置于冰上以终止反应。加入10μl碘化丙啶并通过facs确定细胞的裂解。
[1100]
在wien133细胞上使用不同浓度的正常人血清(nhs)来进一步分析e435r突变增强cdc的能力,据显示该突变能增强7d8和005抗体两者在daudi和raji细胞上的cdc活性。在圆底96孔板中(nunc,rochester,ny),将0.1
×
10
6
个raji细胞与系列浓度的未纯化抗体(0.001、0.003、0.01、0.03、0.1、0.3、1.0、3.0、10.0、30.0μg/ml)在总体积50μl中室温下在振荡器中预温育15分钟。接着,加入nhs作为c1q的来源使其在100μl体积中的终浓度达到20%或50%nhs。将反应混合物在37℃的培养箱中温育45分钟。将平板置于冰上以终止反应。加入10μl碘化丙啶并通过facs确定细胞的裂解。
[1101]
据发现,在ch2-ch3域中被确认的导致cd20抗体7d8丢失或增加cdc活性的突变对识别cd38的005抗体具有相同的作用。图9显示,005-i253d、h443a、k439e和s440k在daudi(图9a)和raji(图9b)细胞上都显示出cdc活性完全丢失,而005-e345r突变则在两个细胞系上均显示出cdc活性强烈增强。005-k439e+005-s440k(作为单突变体这两者都能导致cdc丢失)导致cdc的恢复,比得上7d8的数据。令人吃惊的是,005-e435r甚至能强有力地诱导对wien133细胞的cdc,而野生型005却不能够诱导cdc杀死(图9c)。在20%和50%血清浓度下均观察到005-e345r对wien133细胞的cdc杀死(图9c)。在50%血清中,7d8-e345r和005-e345r在体外对raji细胞也显示增强的cdc,这与在20%血清中的效力相似(图9d)。
[1102]
由于所测试的cd20抗体7d8和cd38抗体005两者的ch2-ch3区中的e345r突变都导致增强的cdc活性,因此e345r突变可被认为是能够用于诱导或增强cdc的普遍性抗体修饰。
[1103]
实施例11
[1104]
含有cdc增强型突变e345r的igg1抗体对于通过fc结合肽dcawhlgelvwct抑制cdc比野生型抗体的敏感性低
[1105]
通过对igg fc:fc界面的疏水片中的氨基酸位点进行突变,发现cdc效力要么被破坏,要么被增强。进一步探究了fc-fc界面处的相互作用,以及因此形成如b12晶体结构中所观察到的寡聚结构(例如,六聚环)参与了cdc效力。因此,使用了靶向在野生型igg fc表面的疏水片区域中的共识别结合位点的13残基肽(dcawhlgelvwct(seq id no:7))(delano等,science2000feb 18;287(5456):1279-83)。确实,igg fc表面上的共识别结合位点作为准备与大量不同分子相互作用的适应性区域的身份(delano等,science 2000feb 18;287(5456):1279-83),与igg1 b12晶体结构中参与fc-fc相互作用的疏水片中的核心氨基酸的身份相一致(saphire等,science 2001aug 10;293(5532):1155-9)。在所有结合界面存在的相互作用受到6个氨基酸的共享组(met-252、ile-253、ser-254、asn-434、his-435和tyr-436)以及共享骨架触点(delano等,science 2000feb 18;287(5456):1279-83)的介导。因此,可认为fc结合肽能影响fc-fc相互作用并从而影响cdc效力。
[1106]
在圆底96孔平板中,将0.1
×
10
6
daudi细胞与1.0μg/ml未纯化抗体于75μl中在摇床中室温预温育10分钟。将25μl浓度系列(范围从0.06-60μg/ml终浓度)的fc结合肽dcawhlgelvwct加入到经调理的细胞中,在摇床中室温温育10分钟。接着,加入25μl nhs作为补体来源(20%终浓度),在培养箱中37℃温育45分钟。加入25μl冰冷rpmi培养基(补充有
ꢀꢀꢀꢀꢀꢀ
平均8,32,5ng/ml sem4,11,9 t检验2-尾p=0,04 因子增强
ꢀꢀ
3,3时间
[1117]
图11显示,相比起野生型抗体humab-005,突变体igg1-005-e345r证明了adcc能力增强的效力,能够在较低浓度时诱导adcc。
[1118]
实施例13
[1119]
与野生型7d8相比7d8突变体的fcrn结合以及药物代谢动力学分析
[1120]
新生fc受体(fcrn)通过保护igg不被降解而成为igg长血浆半衰期的原因。在抗体内在化之后,fcrn在核内体中与抗体fc区结合,而核内体中在略微酸性的环境下(ph6.0)相互作用是稳定的。当回收至质膜时,这里的环境是中性的(ph7.4),相互作用丢失,抗体被释放回血液循环中。这影响了igg的血浆半衰期。
[1121]
在elisa中测试7d8突变体igg1-7d8-e354r与来自于小鼠、食蟹猴和人的fcrn相互作用的能力。所有温育在室温下完成。将5μg/ml重组生产的生物素化的fcrn胞外域(小鼠、人或食蟹猴)(fcrnecdhis-b2m-bio)涂覆在96孔板中(100μl/孔),稀释于pbst加0.2%bsa中;1小时。平板用pbst洗涤3次,加入3-倍系列稀释(于pbst/0.2%bsa,ph6.0)的野生型igg1-7d8或igg1-7d8-e354r,并将平板温育1小时。用ph6.0的pbst/0.2%bsa洗涤平板。加入稀释于ph6.0的pbst/0.2%bsa中的山羊-抗-人igg(fab
’
2)-hrp(jackson immuno research,货号:109-035-097),并将平板温育1小时。洗涤之后,加入abts作为底物,并将平板在黑暗中温育30分钟。使用el808 elisa酶标仪读取405处的吸光度。
[1122]
本研究中的小鼠安置在中央实验动物设施(utrecht,荷兰)的屏障单元中,并以随意提供的水和食物饲养在滤盖饲养笼中。所有的实验都由乌德勒支大学动物伦理委员会批准。
[1123]
为了分析7d8突变体在体内的药物代谢动力学,用100μg(5mg/kg)野生型7d8、igg1-7d8-e354r、-s440k或k322a静脉注射scid小鼠(c.b-17/icrcrl-scid-br,charles-river);每组3只小鼠。
[1124]
在施用抗体后10分钟、4小时、24小时、2天、7天、14天和21天从隐静脉血管中收集50μl血样。将血液收集进含有肝素的小瓶中,并以10,000g离心5分钟。将血浆储藏于-20℃直至确定mab浓度。
[1125]
使用夹心式elisa来确定人igg浓度。将以2μg/ml的浓度涂覆在96孔microlon elisa平板(greiner,德国)上的小鼠mab抗-人igg-κ克隆mh16(#m1268,clb sanquin,荷兰)用作捕获抗体。将平板用补充有2%鸡血清的pbs封闭之后,加入样本,系列稀释于elisa缓冲液中(补充有0.05%吐温20和2%鸡血清的pbs),室温下(rt)温育于平板摇床中1小时。随后用山羊抗-人igg免疫球蛋白(#109-035-098,jackson,west grace,pa)温育平板,并用2,2
’-
连氮-双-(3-乙基苯并噻唑啉-6-磺酸)(abts,roche,mannheim,德国)显影。在微板酶标仪上(biotek,winooski,vt)测量405nm处的吸光度。
[1126]
选择scid小鼠是因为它们的血浆igg浓度低,并因此具有相对低的igg清除。这提供了对检测清除中的变化(由于fcγ-部分与新生fc受体(fcrn)结合的减少)非常敏感的pk
模型。
[1127]
使用graphpad prism版本4(graphpad software)来进行统计学测试。
[1128]
图12显示野生型humab-7d8和igg1-7d8-e345r两者都与小鼠、人和食蟹猴fcrn结合良好。igg1-7d8-e345r的结合稍好于野生型7d8的结合。
[1129]
图13显示血浆浓度随时间的变化。野生型humab-7d8相比起igg1-7d8-e345r、-s440k或k322a中任何一个在血浆浓度随时间变化(清除)方面并没有差异。
[1130]
实施例14
[1131]
利用稳定fc-fc的突变e345r来提高igg抗体针对表达fc结合表面蛋白的细菌的杀菌活性
[1132]
补体级联系统是重要的针对病原体的宿主防御机制,其可以被分为三种不同的的激活途径以识别病原体:i)抗体介导的经典途径,通过c1q结合至与病原体结合的抗体来激活,ii)凝集素和iii)替代途径,其中在没有抗体存在的情况下补体系统直接识别病原体并由病原体激发。这三条途径汇聚于c3切割和c3b沉积的步骤。微生物已经进化出多种补体逃避机制,其中一种是由蛋白a介导的(joiner ann.rev.microbiol.(1988)42:201-30;foster nat rev microbiol(2005)dec;3(12):948-58)。蛋白a是在金黄色葡萄球菌(staphylococcus aureus)的细胞壁中被首次鉴定出来,并因其与igg的fc区结合而被人所熟知(deisenhofer等.,biochem(1981)20,2361-70;uhlen等.,j.biol.chem(1984)259,1695-1702)。迄今为止,蛋白a的抗噬菌功能及其在金黄色葡萄球菌发病机制中的作用可由蛋白a和igg之间的相互作用来解释,其导致不正确的抗体方向将被嗜中性fc受体所识别(foster nat rev microbiol(2005)dec;3(12):948-58)。
[1133]
实施例11显示,由b细胞特异性igg1抗体所介导的cdc被竞争的fc-结合肽dcawhlgelvwct所抑制。该肽靶向igg fc上与蛋白a、蛋白g和类风湿因子的结合位点相一致的共有结合位点(delano等,science 2000feb 18;287(5456):1279-83)。基于这些数据,相信蛋白质a介导的细菌补体逃逸机制可通过与fc结合竞争来起作用,导致微生物特异性抗体的fc-fc相互作用不稳定,并因此抑制抗体介导的补体激活。此外,实施例11还显示含有cdc-增强的e345r突变的b细胞特异性igg1抗体对于通过竞争性fc-结合肽dcawhlgelvwct抑制cdc要比亲本野生型抗体更不敏感。通过将这些结果外推至微生物表达的fc结合蛋白,由e345r突变而提高的igg1 fc-fc相互作用稳定性使得微生物特异性抗体更不易于借助微生物表面蛋白(如蛋白a)通过fc结合竞争的病原体逃逸策略来抑制补体。因此,与亲本野生型抗体相比,将e345r突变引入到定向针对细菌的igg抗体将导致c3b在细菌上的沉积增加以及杀菌活性的提高。
[1134]
作为补体介导的细菌杀死的体外测量,嗜中性粒细胞的细胞吞噬以及血浆中c3a的生成两者(其与c3b在细菌上的沉积一致)可如下所述确定。实际上,已经描述了c3b在金黄色葡萄球菌上的沉积导致增强的细胞吞噬作用并与细胞杀死有关(rooijakkers等,nature immunology 2005:6,920-927)。
[1135]
通过将指数生长的细菌培养物与100μg/ml fitc在0.1m碳酸盐缓冲液(ph 9.6)中在37℃温育1小时来用fitc标记金黄色葡萄球菌。利用ficoll梯度来分离人多形核细胞(pmn)。用系列浓度的具有或不具有e345r的特异性抗体来调理fitc标记的细菌。通过使1
×
10
8
个经过调理的fitc标记细菌与人pmn在存在25%igg缺失血清作为补体来源的情况下在
总体积200μl中在37℃在强烈振荡的条件下温育25分钟来体外进行细胞吞噬作用。固定细胞并通过在室温与bd facs裂解溶液温育15分钟来进行红细胞溶解。洗涤之后,通过facs来测量细胞吞噬。通过正向和侧向散射门限来选择嗜中性粒细胞群体,而细胞吞噬则以嗜中性粒细胞群体中的平均荧光表示。或者,通过elisa作为补体激活和c3b沉积的测量来对样品中的c3a生成进行测量。
[1136]
据估计含有e345r突变的金黄色葡萄球菌特异性抗体比起亲本野生型抗体将诱导更多的补体激活以及嗜中性粒细胞的细胞吞噬。可用于此类实验的抗体的示例为嵌合单克隆igg1帕吉昔单抗(bsyx-a110;biosynexus),其靶向嵌入(embed)在葡萄球菌细胞壁之中的脂磷壁酸(lta)(baker,nat biotechnol.2006dec;24(12):1491-3;weisman等,int immunopharmacol.2009may;9(5):639-44)。
[1137]
实施例15
[1138]
限制对于同时被两种不同治疗性单克隆抗体的混合物所结合的靶细胞的cdc激活的cdc-抑制性突变的用途
[1139]
如实施例6所述,cd20抗体7d8突变k439e和s440k作为单克隆抗体降低了cdc效力。含有这些突变的混合7d8抗体恢复cdc。因此有效cdc受限于同时结合有两个突变体抗体的细胞。如实施例10所述,cd38抗体005突变k439e和s440k作为单克隆抗体降低了cdc效力。含有这些突变的混合005抗体恢复cdc。因此有效cdc受限于同时结合有两个突变体抗体的细胞。
[1140]
限制针对同时表达两种特异性抗原的靶细胞的有效cdc的诱导是有利的,这利用了它们的组合表达来改善cdc诱导的选择性。为了限制针对同时结合有cd20和cd38抗体的细胞的cdc诱导,在cdc实验中如下单独加入或以1:1混合7d8-k439e和005-s440k对或者7d8-s440k和005-k439e对。在圆底96孔板中,将0.1
×
10
6
个daudi或raji细胞与系列浓度的未纯化抗体或抗体混合物(0.01、0.03、0.1、0.3、1.0、3.0、10.0、30.0μg/ml)在总体积100μl中在室温在摇床中预温育15分钟。接着,加入25μl正常人血清作为补体来源(20%终浓度),在37℃的培养箱中温育45分钟。将平板置于冰上以终止反应。加入10μl碘化丙啶并在facs上确定细胞的裂解。据估计,7d8-k439e、005-s440k、7d8-s440k和005-k439e将表现出有限的cdc效力。据估计,同时加入7d8-k439e和005-s440k将恢复特异性针对表达cd20和cd38两者的细胞的有效cdc。同样,据估计7d8-s440k和005-k439e的混合物将恢复特异性针对表达cd20和cd38两者的细胞的有效cdc。
[1141]
实施例16
[1142]
通过将e345r与互补的抑制性突变k439e和s440k组合在两种不同单克隆抗体的混合物中来提高增强型cdc的特异性
[1143]
如实施例6所述,cd20抗体7d8突变k439e和s440k作为单克隆抗体降低了cdc效力。含有这些突变的混合7d8抗体恢复cdc。因此有效cdc受限于针对同时结合有两个突变体抗体的细胞。如实施例10所述,cd38抗体005突变k439e和s440k作为单克隆抗体降低了cdc效力。含有这些突变的混合005抗体恢复cdc。因此有效cdc受限于同时结合有两个突变体抗体的细胞。
[1144]
限制针对同时表达两种特异性抗原的细胞的cdc诱导的增强是有利的,这利用了它们的组合表达来改善增强的cdc诱导的可选择性。限制针对同时结合至少两种不同抗体
的混合物的靶细胞的cdc诱导的增强同样是有利的,所述抗体在两个不同的表位上同时,或在两个交叉竞争、相似或相同的表位上结合相同的细胞表面抗原。
[1145]
因此,为了限制针对同时结合有cd20和cd38抗体两者的细胞的增强性cdc诱导,将cdc增强突变e345r与cdc抑制性突变组合在抗体7d8-e345r/k439e、7d8-e345r/s440k、005-e345r/s440k和005-e345r/k439e中。在cdc实验中如下单独加入或以1:1混合这些抗体。在圆底96孔平板中,将0.1
×
10
6
个wien133细胞(也可以使用其它的细胞类型,例如daudi或raji细胞)与系列浓度的未纯化抗体(终浓度0.056-10,000ng/ml以3倍稀释的7d8
--
e345r/k439e、7d8-e345r/s440k、005-e345r/s440k和005-e345r/k439e)或抗体混合物(终浓度为0.01μg/ml cd20抗体与0-333ng/ml以3倍稀释的cd38抗体混合;或3.3μg/ml的cd38抗体与0.0056-1,000ng/ml以3倍稀释的cd20抗体混合)在总体积100μl中在室温在摇床中预温育15分钟。接着,加入25μl正常人血清作为补体来源(20%终浓度),在37℃的培养箱中温育45分钟。将平板置于冰上以终止反应。加入10μl碘化丙啶并通过facs确定细胞的裂解。
[1146]
将系列浓度的005-e345r/k439e或005-e345r/s440k抗体与固定浓度为0.01μg/ml的7d8双突变体抗体混合(如从图14a中所确定的作为单剂对wien133细胞具有最小cdc的最大浓度)以制备互补性组合005-e345r/k439e+7d8-e345r/s440k或005-e345r/s440k+7d8-e345r/k439e。图14c显示,在分别存在固定浓度的互补性7d8-e345r/k439e或7d8-e345r/s440k cd20抗体时,005双突变cd38抗体剂量依赖性地诱导cdc。这些互补性组合的cdc效力(图14c)与005-e345r单突变体(增强物)抗体作为单剂相当(图14b)。相反,在存在无关抗体b12时,005-e345r/k439e或005-e345r/s440k在测试的系列浓度中几乎没有显示出任何cdc(与005-e345r/k439e或005-e345r/s440k作为单剂相当,图14b中所示)。
[1147]
将系列浓度的7d8-e345r/k439e或7d8-e345r/s440k抗体与固定浓度为3.3μg/ml的005双突变体抗体混合(如从图14b中所确定的作为单剂对wien133细胞显示出少许但有限的cdc)以制备互补性组合7d8-e345r/k439e+005-e345r/s440k或7d8-e345r/s440k+005-e345r/k439e。图14d显示在分别存在互补性005-e345r/k439e或005-e345r/s440k cd38抗体的情况下,7d8双突变体cd20抗体非常有效地诱导了cdc,甚至是在测试的最低浓度时,就像每个细胞只有一些7d8双突变体抗体分子。为了评估细胞膜上fc尾部密度的提高对于所观察到的由具有互补性k439e和s440k突变的7d8和005抗体混合物增强的cdc的贡献,还测试了具有非互补性突变的抗体组合。图14d显示,非互补性组合显示出远低于互补性组合的cdc效力,这是由于比互补性组合效率更低的fc-fc相互作用。
[1148]
这些数据表明,由治疗性抗体的(增强的)cdc诱导可受限于同时结合两个互补性抗体混合物的细胞,在这种情况下具有不同的抗原特异性,从而通过需要这些抗原共表达来提高靶细胞特异性。
[1149]
如图14a和14b中所见,与单独的7d8-e345r相比,7d8-e345r/k439e、005-e345r/s440k、7d8-e345r/s440k和005-e345r/k439e展现出有限的cdc效力。可进一步见到的是,7d8-e345r/k439e和7d8-e345r/s440k与作为单剂的野生型7d8抗体相比使得cdc具有增强的效力。同样地,也观察到005-e345r/k439e和005-e345r/s440k的混合物与作为单剂的野生型005抗体相比使得cdc具有增强的效力(数据未显示)。
[1150]
实施例17
[1151]
将有效cdc激活限制于专门由治疗性施用的抗体组成的抗体复合物的cdc抑制性
突变的用途
[1152]
如实施例6中所述,cd20抗体7d8双突变体k439e/s440k恢复了由k439e或s440k单点突变体所减少的cdc效力。如实施例10中所述,cd38抗体005双突变体k439e/s440k恢复了由k439e或s440k单点突变体所抑制的cdc效力。正如所观察到的,单点突变破坏了在fc:fc界面面向侧(facing side)上具有未突变氨基酸的fc:fc相互作用。在fc:fc界面的面向侧引入补偿性突变恢复了cdc效力。因此有效的cdc显然受限于专门由含有这两个突变的抗体组成的抗体复合物。
[1153]
在另外的实施例中,cdc的诱导受限于专门由治疗性施用的抗体组成的抗体复合物。为了专门限制针对结合有cd20或cd38抗体的细胞的cdc诱导,将cdc抑制性突变k439e和s440k组合到抗体7d8-k439e/s440k或005-k439e/s440k中。这些抗体在存在或不存在非靶标特异性的igg时如下单独加入到cdc实验中。在圆底96孔板中,将0.1
×
10
6
个daudi或raji细胞与未纯化的抗体或抗体混合物(0.01、0.03、0.1、0.3、1.0、3.0、10.0、30.0μg/ml)在总体积100μl中在室温在摇床中预温育15分钟。接着,加入25μl正常人血清作为补体来源(20%终浓度),在37℃的培养箱中温育45分钟。将平板置于冰上终止反应。加入10μl碘化丙啶并通过facs确定细胞的裂解。
[1154]
预计7d8-k439e/s440k将诱导具有与野生型7d8抗体相似效力的cdc。预计向7d8-k439e/s440k加入非特异性igg不会影响该抗体的cdc诱导的效力。同样地,预计005-k439e/s440k将使cdc具有与野生型humab 005相似的效力。预计向005-k439e/s440k加入非特异性igg不会影响该抗体的cdc诱导的效力。
[1155]
实施例18
[1156]
将增强的cdc激活限制于专门由治疗性施用的抗体组成的抗体复合物的cdc抑制性突变的用途
[1157]
如实施例6中所述,cd20抗体7d8双突变体k439e/s440k恢复了由k439e或s440k单点突变体所减少的cdc效力。如实施例10中所述,cd38抗体humab 005双突变体k439e/s440k恢复了由k439e或s440k单点突变体所抑制的cdc效力。正如所观察到的,单点突变破坏了在fc:fc界面面向侧(facing side)上具有未突变氨基酸的fc:fc相互作用。在fc:fc界面的面向侧引入补偿性突变恢复了cdc效力。因此有效的cdc显然受限于专门由含有这两个突变的抗体组成的抗体复合物。
[1158]
在另外的实施例中,cdc诱导的增强限制于专门由治疗性施用的抗体组成的抗体复合物。通过筛选和选择激发用于cdc激发的fc:fc相互作用的突变,人们可以利用对于感兴趣抗原靶标非特异性的血清抗体鉴定能够形成cdc诱导抗体复合物的突变。为了专门限制针对结合有cd20或cd38抗体的细胞的增强型cdc诱导,将cdc增强性突变e345r与cdc抑制性突变组合到抗体7d8-e345r/k439e/s440k或005-e345r/k439/s440k中。这些抗体在存在或不存在非靶标特异性的igg时如下单独加入到cdc实验中。在圆底96孔板中,将0.1
×
10
6
个daudi或raji细胞与未纯化的抗体或抗体混合物(0.01、0.03、0.1、0.3、1.0、3.0、10.0、30.0μg/ml)在总体积100μl中在室温在摇床中预温育15分钟。接着,加入25μl正常人血清作为补体来源(20%终浓度),在37℃的培养箱中温育45分钟。将平板置于冰上终止反应。加入10μl碘化丙啶并通过facs确定细胞的裂解。
[1159]
预计与野生型humab 7d8相比,7d8-e345r/k439e/s440k会诱导具有增强效力的
cdc。向7d8-e345r/k439e/s440k加入非特异性igg预计不会影响与野生型7d8抗体相比的cdc诱导的效力。同样地,预计与野生型005抗体相比,005-e345r/k439e/s440k将使cdc具有增强的效力。向005-e345r/k439e/s440k加入非特异性igg预计不会影响与野生型005抗体相比的cdc诱导的效力。
[1160]
实施例19
[1161]
使用突变体筛选方法鉴定能激发通过cdc测定法检测的fc:fc相互作用介导的抗体寡聚化的突变
[1162]
如实施例6和10所述,对于在表达各种水平的所述抗原的多个细胞系上识别两种不同靶标抗原(cd20和cd38)的抗体,鉴定了激发cdc的氨基酸突变。令人惊奇的是,单点突变e345r证实足以将wien133细胞的cdc依赖性细胞裂解赋予抗cd38抗体005,但该抗体以野生型igg1形式不能通过cdc来裂解这些细胞。
[1163]
位于或在fc:fc界面周围的其他突变能够以类似的方式激发寡聚化和cdc。或者,突变能够间接激发寡聚化,例如通过变构性诱导fc:fc相互作用。
[1164]
为了确定其他氨基酸突变是否能够激发fc介导的抗体寡聚化,使用cdc测定法对抗cd38 igg1-005突变体文库进行筛选,单独地及以成对的方式混合以选择例如在fc:fc界面两侧相互作用的氨基酸对。然而,可以将相同的策略用于其他的抗体,例如另外的igg1或igg3抗体。
[1165]
产生了位于表15中所示位置的突变的集中文库。使用quikchange定点诱变试剂盒(stratagene,美国)将突变引入至igg1-005 fc区中。简而言之,对于每个所期望的突变位置,使用在所期望位置上编码简并密码子的正向和反向引物来复制具有igg1m(f)同种异型的005重链的全长质粒dna模板。用dpni消化所得到的dna混合物以移除来源质粒dna并用于转化大肠杆菌。汇集并培养所得到的菌落,从这些汇集物中分离质粒dna并转化入大肠杆菌中以获得克隆菌落。通过dna测序(lgc genomics,berlin,德国)来验证从所得到的菌落中分离出来的突变体质粒dna。通过pcr从质粒dna中扩增表达盒,并使用293fectin(invitrogen,美国)基本上如制造商所述将同时含有igg1-005突变体重链和野生型轻链两者的dna混合物瞬时转染至freestyle hek293f细胞(invitrogen,美国)。收集含有抗体突变体的转染细胞的上清液。在cdc测定法中单独地以及配对混合地如下筛选突变体抗体上清液。
[1166]
在圆底96孔板中,将0.1
×
10
6
个daudi或wien-133细胞(也可使用其它的细胞类型,例如raji细胞)与1.0μg/ml未纯化抗体在总体积100μl中在室温在摇床中预温育15分钟。接着,加入30μl正常人血清作为补体来源(30%终浓度)并在37℃的培养箱中温育45分钟。将平板置于冰上以终止反应。加入10μl碘化丙啶并通过facs确定细胞的裂解。
[1167]
针对其增强寡聚化的能力(如通过cdc效力检测的),无论是作为单个突变体还是与其他突变体混合(例如面向fc:fc界面对面的突变),对描述于表16、表17和表18中的突变进行选择。针对其不会损坏fcrn、蛋白-a或蛋白-g结合、adcc、adcp或其他由fc域介导的效应子功能的能力,可选择进一步对突变进行筛选。将此类激发性点突变组合到一个fc域中甚至能够更进一步地激发寡聚化和cdc效力。
[1168]
针对其抑制寡聚化的能力(如对于daudi细胞通过cdc确定的),对并入cd38抗体005中的ch2-ch3区中的突变进行测试。突变体抗体的裂解与野生型005(其裂解被设置为
100%)相比较。抑制的截断值被设定为≤66%裂解。用这种方式进行测量,大部分所测试的突变都抑制cdc(见表16)。
[1169]
针对其抑制寡聚化的能力(如对于wien133细胞通过cdc确定的),对并入cd38抗体005中的ch2-ch3区中的突变进行测试(表17)。野生型cd38抗体005不能够诱导针对wien133细胞的cdc。将显示出≥39%细胞裂解的突变体评价为增强。完全预料不到的是,几乎所有获得的氨基酸e345和e430取代都激发通过cdc的细胞裂解。为了证实这个结果,通过定点诱变用每种可能的突变对氨基酸e345、e430和s440进行取代,并测试它们增强寡聚化的能力(如使用新鲜人血清批次通过wien133细胞的cdc确定的),产生略微更有效的裂解(表18)。再一次,所有的e345和e430取代都诱导了wien133细胞的有效cdc。
[1170]
下述优选的突变引起了wien133细胞≥39%的细胞裂解:p247g、i253v、s254l、q311l、q311w、e345a、e345c、e345d、e345f、e345g、e345h、e345i、e345k、e345l、e345m、e345n、e345p、e345q、e345r、e345s、e345t、e345v、e345w、e345y、d/e356g、d/e356r、t359r、e382l、e382v、q386k、e430a、e430c、e430d、e430f、e430g、e430h、e430i、e430l、e430m、e430n、e430p、e430q、e430r、e430s、e430t、e430v、e430w、e430y、y436i、s440y和s440w。
[1171]
[1172][1173]
实施例20
[1174]
皮下b细胞淋巴瘤移植模型中igg1-7d8-e345r的体内效力
005-e345r抗体处理的小鼠中的肿瘤形成与用阴性对照抗体igg1-b12处理的小鼠(mantel-cox分析,p<0.001)或者用野生型igg1-005处理的小鼠(p<0.05)相比被显著延迟。
[1186]
这些数据显示,在cd38抗体005中引入e345r突变导致增强的体内抗肿瘤活性。
[1187]
表19治疗组和剂量
[1188]
组抗体剂量1野生型igg1-005-wt500μg(=25mg/kg)2cdc-增强突变体igg1-005-e345r500μg(=25mg/kg)3无关抗体对照igg1-b12500μg(=25mg/kg)
[1189]
实施例22
[1190]
单价靶标结合进一步增强e345r抗体的cdc效力
[1191]
在b12晶体结构中观察到的igg1六聚环分子表面表明,对于六聚环中的每个igg,两个c1q结合位点中其中一个面朝环结构的上方而另外一个则面朝环结构的下方,并且每个抗体的fab-臂也是一个方向朝上,一个方向朝下,这导致每个抗体只有一个fab-臂参与到抗原结合中,这暗示了六聚抗体环中每个抗体分子的单价结合。单价性可能会使抗体的抗原结合以六聚化相容性方向进行。为了测试这个设想,在cd38阳性、egfr-阴性的wien133细胞上(对于该细胞,这个双特异性抗体仅能通过cd38而进行单价结合)测试具有e345r突变的双特异性cd38/egrf抗体的cdc效力,并与同样具有e345r突变双价结合cd38的抗体的cdc效力进行比较。将人单克隆抗体humax-egfr(2f8,描述于wo2004/056847中)用作本实施例中所述egfr抗体的基础。
[1192]
根据duobody
tm
平台,即如wo2011/147986中所述的2-mea-诱导的fab-臂交换,体外产生双特异性抗体。该方法的基础是互补性ch3域(其在特定的测定条件下促进杂二聚体的形成)的使用。为了通过这个方法来进行双特异性抗体的生产,产生了在ch3域中携带特定突变的igg1分子:在亲本igg1抗体的其中之一上是f405l突变,而在另一个亲本igg1抗体上是k409r突变。为了产生双特异性抗体,将这两个亲本抗体(每个抗体的终浓度为0.5mg/ml)与25mm的2-巯基乙胺-hcl(2-mea)在总体积100μl的te中于37℃温育90分钟。当根据制造商规程使用离心柱(microcon离心过滤器,30k,millipore)移除还原剂2-mea时,还原反应终止。
[1193]
对于cdc测定,在圆底96孔板中,将0.1
×
10
6
个wien133细胞与系列浓度的抗体(0.01至10.0μg/ml)在总体积100μl中在室温在摇床中预温育15分钟。接着,加入25μl正常人血清作为补体来源(20%终浓度),在37℃的培养箱中温育45分钟。将平板置于冰上终止反应。加入10μl碘化丙啶并通过facs确定细胞的裂解。
[1194]
图17显示,如预期的,不具有e345r突变的cd38抗体(野生型igg1-005和igg-b12-k409r
×
igg1-005-f405l)并没有诱导wien133细胞的杀死。如预期的,egfr抗体igg1-2f8-e345r/f405l(其并不结合egfr-阴性wien133细胞(数据未显示))也没有诱导cdc。k408r突变的引入并没有影响igg1-005-e345r抗体在wien133细胞上诱导~60%杀死的能力(描述于实施例10中)。有趣的是,双特异性cd38/egfr抗体igg1-005-e345r/k409r
×
igg1-2f8-e345r/f405l(其只能单价地结合cd38阳性、egfr-阴性wien133细胞)显示出提高的最大cdc杀死(从~60%至~100%杀死)。
[1195]
这些数据显示,单价靶向能够进一步增强含有cdc增强型e345r突变的抗体的最大
杀死能力。此外,这些数据显示,e345r寡聚化增强突变(如通过增强cdc活性来测量)能够应用于其他抗体形式中,如duobody。
[1196]
实施例23
[1197]
寡聚化增强e345r突变能够应用于其他抗体形式如duobody
tm
[1198]
在duobody形式的双特异性抗体中测试e345r突变的效果。利用cd20/cd38双特异性抗体在cd20-阳性、cd38-阳性wien133和raji细胞上实施cdc测定。
[1199]
如实施例22中所述产生双特异性抗体。对于cdc测定,在圆底96孔板中,将0.1
×
10
6
个wien133或raji细胞与系列浓度的抗体(0.01至30.0μg/ml)在总体积100μl中在室温在摇床中预温育15分钟。接着,加入25μl正常人血清作为补体来源(20%终浓度),在37℃的培养箱中温育45分钟。将平板置于冰上终止反应。加入10μl碘化丙啶并通过facs确定细胞的裂解。
[1200]
图18显示引入e345r突变增强了igg1-005-f405l
×
igg1-7d8-k409r抗体对wien133(图18a)和raji(图18b)细胞的cdc。这些数据显示,e345r寡聚化增强型突变能够应用于其他抗体形式以增强cdc活性。
[1201]
实施例24
[1202]
e345r拯救egfr抗体2f8的cdc,其能够通过单价靶标结合进一步增强
[1203]
如实施例6、10和26中所述,e345r增强或拯救了识别不同血液肿瘤靶标(cd20和cd38)的抗体的cdc。为了将该分析延伸至实体瘤抗原,在a431表皮癌细胞上测试e345r对egfr抗体2f8的cdc能力的影响。此外,利用双特异性egfr
×
cd20抗体(igg1-2f8-e345r/f405l
×
igg1-7d8-e345r/k409r)在egfr-阳性、cd20阴性的a431细胞上测试单价性egfr靶向对于e345r介导的cdc诱导的影响。
[1204]
如实施例22所述产生双特异性抗体。对于cdc测定法,用100μci
51
cr对5
×
10
6
个a431细胞/ml在37℃标记1小时。用pbs洗涤细胞三次,并以1
×
10
5
个细胞/ml的浓度重悬于培养基中。在圆底96孔板中,将25,000个标记细胞与系列浓度的未纯化抗体(0-30μg/ml以3倍稀释)在总体积100μl中在室温温育15分钟。接着,加入50μl正常人血清稀释物作为补体来源(25%终浓度),在37℃的培养箱中温育1小时。将细胞旋转沉淀下来(300
×
g,3分钟),并在96孔optiplate(perkinelmer)中向100μl microscint加入25μl上清液以在摇床(750rpm)上温育15分钟。在闪烁计数器上以每分钟计数(cpm)来确定
51
cr释放。通过在经triton x-100处理细胞上清液中测量的
51
cr水平来确定最大裂解(100%)。通过在未经抗体温育的细胞上清液中测量的
51
cr水平来确定自发裂解。根据公式:特异性裂解=100
×
(cpm样本-cpm自发)/(cpm最大-cpm自发)来计算特异性的细胞裂解。
[1205]
图19显示igg1-2f8-e345r/f405l能够通过cdc裂解a431细胞,而野生型2f8却不能杀死a431细胞。这些数据显示,通过引入e345r突变能够拯救egfr抗体2f8的cdc活性。这可能将cdc增强型e345r突变的应用扩展至靶向实体瘤抗原的抗体。
[1206]
双特异性egfr
×
cd20抗体igg-2f8-e345r/f405l
×
igg1-7d8-e345r/k409r,在egfr-阳性、cd20阴性的a431细胞上显示cdc的进一步增强。
[1207]
这些数据进一步支持了该设想,即单价性促进fc-fc相互作用的形成及随后的cdc诱导,如对于实施例22中所述的cd38结合抗体所提出的。
[1208]
实施例25
[1209]
e345r增强或拯救cd38抗体003以及cd20抗体11b8和利妥昔单抗的cdc
[1210]
正如实施例6、10和24所述,e345r增强或诱导了具有不同靶标特异性(cd20、cd38和egfr)的几个抗体的cdc活性,如在表达多种水平的所述抗原的细胞系上所测试的。因此,e345r突变的引入认为是对目前抗体进行增强或拯救cdc的普遍性机制。为了进一步支持这个,对更多的对daudi和wien133细胞具有不同的固有cdc效力的抗体测试e345r突变对cdc的影响:cd38抗体003,描述于wo 2006/099875中以及cd20抗体利妥昔单抗(i型)和11b8(ii型),描述于wo 2005/103081中。cd20抗体可被分为两个亚群(beers等seminars in hematology 47,(2)2010,107-114)。i型cd20抗体展示出卓越的通过将质膜中的cd20分子重新分布于脂筏上(其使得抗体fc区聚集并能够改进c1q结合)从而激活补体以及激发cdc的能力。ii型cd20抗体并没有可感知地改变cd20的分布,并且也没有伴随的聚集,它们在cdc方面是相对无效的。
[1211]
在圆底96孔板中,将0.1
×
10
6
个daudi或raji细胞与系列浓度的未纯化抗体(0.001、0.003、0.01、0.03、0.1、0.3、1.0、3.0、10.0μg/ml)在总体积70μl中在室温在摇床中预温育15分钟。接着,加入30μl正常人血清作为c1q来源(30%终浓度),并在37℃的培养箱中温育45分钟。将平板置于冰上以终止反应。加入10μl碘化丙啶并通过facs确定细胞的裂解。
[1212]
图20显示,e345r突变增强了所有测试的抗体对daudi(a)和wien133(b)细胞两者的cdc。有趣的是,在所使用的浓度,以野生型形式存在时没有诱导cdc的所有抗体,在引入e345r突变之后都有效地诱导了cdc:cd38 mab 003和cd20 ii型mab 11b8对daudi细胞,以及cd38 mabs 005和003以及cd20 ii型mab11b8对wien133细胞。这些数据表明抗体寡聚化的增强,更具体地通过e345r突变的引入而增强,是增强或拯救目前抗体的cdc的普遍性机制。
[1213]
实施例26
[1214]
e345r增强了组织因子抗体的内在化
[1215]
为了测试增强的寡聚化能否诱导提高的抗体内在化,利用溶酶体标记物lamp1通过共聚焦显微镜来进行野生型和e345r突变型组织因子(tf)抗体的共定位研究。
[1216]
sk-ov-3细胞在玻璃盖玻片上(厚度1.5微米,thermo fisher scientific,braunschweig,德国)于标准组织培养基中在37℃生长1天。将细胞与50μg/ml亮肽酶素(leupeptin)(sigma)预温育1小时以封闭溶酶体活性,其后加入10μg/ml的组织因子(tf)抗体(wo2010/066803)。在37℃将细胞再温育1、3或16个小时。之后,用pbs洗涤细胞,并用4%甲醛(klinipath)在室温下(rt)温育30分钟。用封闭缓冲液(补充有0.1%的皂苷[roche]和2%bsa[roche]的pbs)洗涤盖玻片,并与含有20mm nh
4
cl的封闭缓冲液温育20分钟以淬灭甲醛。用封闭缓冲液再次洗涤盖玻片,将其与用于鉴定溶酶体lamp1的鼠-抗-人cd107a-apc(bd pharmingen)和用于鉴定tf抗体的山羊-抗-人igg-fitc(jackson)的混合物在室温下温育45分钟。用封闭缓冲液再次洗涤盖玻片并使用20μl封固培养基(将6克glycerol[sigma]和2.4克mowiol 4-88[omnilabo]溶解于6ml蒸馏水中,向其中加入12ml 0.2m tris[sigma]ph8.5,随后在50-60℃温育10分钟;将封固培养基分装并储存于-20℃)封固在显微镜载物片上过夜。用配备63
×
1.32-0.6油浸物镜和las-af软件的leica spe-ii共聚焦显微镜(leica microsystems)对玻片成像。
[1217]
使用metamorph软件(版本meta series 6.1,molecular devices inc,sunnyvale california,美国)对12-位灰度级的tiff图像分析共定位。以层叠输入图像并扣除背景。对于所有的fitc图像和所有的apc图像都使用相同的阈值设定(手动设置)。将共定位描绘为感兴趣区域中(roi)的fitc的像素强度,而roi由所有的apc阳性区组成。为了对用不同tf抗体染色的不同玻片进行比较,用apc的像素强度对图像标准化。使用鼠抗人cd107a-apc对溶酶体标记lamp1(cd107a)进行染色。lamp1的像素强度在所成像的不同tf抗体之间不应当有不同。
[1218]
fitc和apc共定位的标准化数值根据公式[(tpi fitc
×
共定位百分比)/100]
×
[1/tpi apc]表示为任意单位。
[1219]
共定位百分比=与apc像素共定位的tpi fitc/tpi apc
[1220]
tpi:总像素强度
[1221]
图21描绘了与apc标记的溶酶体标记物重叠的野生型和e345r突变型tf抗体的fitc像素强度的量。对于每个抗体或所测试的条件,分析来自含有~1、3或>5个细胞的一个玻片的三张不同图像。在每个玻片中的不同图像之间观察到了差异。然而,明显的是当与野生型011和098相比时,抗体011和098的e345r突变在1小时的温育之后导致溶酶体共定位的提高。这些结果指示突变e345r诱导了更为迅速的内在化和溶酶体共定位,并因此能够增强抗体药物缀合。
[1222]
实施例27
[1223]
在具有相似cd20表达但具有不同水平的膜结合补体调控蛋白的不同b细胞系中利妥昔单抗中的e345r突变增强了cdc
[1224]
实施例25和28显示,野生型利妥昔单抗对于daudi和wien133细胞的cdc效力通过引入e345r突变而被增强。这种增强的cdc效力是由于e345r介导的fc-fc相互作用稳定化造成的。在靶细胞膜上伴随形成的六聚抗体环结构通过促进靠近细胞膜的激活的补体组分的捕获和浓缩能够促进膜攻击复合体的有效产生。由于这种有效的补体激活,能够部分地克服膜结合补体调控蛋白(mcrp)的抑制效果。mcrps(如cd55、cd46和cd59)的过表达认为是利用单克隆抗肿瘤抗体成功进行免疫治疗的障碍(jurianz等,mol immunol 1999 36:929-39;fishelson等,mol immunol 2003 40:109-23,gorter等,immunol today1999 20:576-82,zell等,clin exp immunol.2007dec 150(3):576-84)。因此,比较利妥昔单抗-e345r与野生型利妥昔单抗对一系列具有不同水平的mcrps cd46、cd55和cd59但是具有相当水平的cd20靶标表达的b细胞系的效力。
[1225]
b细胞系daudi、wil2-s、wsu-nhl、mec-2和arh-77表达了相同量的cd20分子(~250.000特异性抗体结合能力-sabc),如通过qifikit分析确定的(数据未显示)。为了比较这些细胞系之间补体调控蛋白的表达水平,实施qifikit分析以确定cd46(小鼠抗人cd46、cbl488、克隆j4.48 chemicon)、cd55(小鼠抗人cd55、cbl511、克隆bric216、chemicon)和cd59(小鼠抗人cd59、mca1054x、克隆mem-43、serotec)的水平。
[1226]
对于cdc测定,在圆底96孔板中,将0.1
×
10
6
个细胞与饱和抗体系列浓度(0.002-40.0μg/ml以4倍稀释)在总体积100μl中室温下在摇床中预温育15分钟。接着,加入25μl正常人血清作为补体来源(20%终浓度),在37℃的培养箱中温育45分钟。将平板置于冰上以终止反应。加入10μl碘化丙啶并通过facs确定细胞的裂解。在graphpad prism 5中利用非
线性拟合的最佳拟合值的顶点从两个独立实验中计算最大cdc介导杀死。
[1227]
图22a-d显示在野生型利妥昔单抗中引入e345r导致对所有测试的b细胞系的cdc效力增强,如通过提高的最大裂解和降低的ec
50
所观察到。
[1228]
图22e显示,由利妥昔单抗-e345r突变体诱导的最大cdc-介导杀死总是比野生型利妥昔单抗要高,而独立于膜结合补体调控蛋白的表达水平。这些数据指示e345r的引入增强了单克隆抗体的治疗潜能,这是由于肿瘤细胞在规避由含有e345r抗体所导致的抗体介导的补体攻击方面较不有效。
[1229]
实施例28
[1230]
野生型和e345r抗体的cdc动力学的比较
[1231]
已经显示引入稳定fc:fc相互作用的e345r突变增强或拯救cdc,如通过ec
50
值降低以及实施例6(cd20抗体7d8对daudi和raji)、实施例10(cd38抗体005对daudi、raji和wien133)和实施例25中(cd38抗体003和cd20抗体利妥西单抗和11b8对daudi和wien133)所述的不同抗体对不同细胞系的最大裂解提高所观察到的。接着,对cdc反应的动力学进行分析以进一步揭示野生型和e345r抗体的cdc效力之间的差别。
[1232]
在圆底96孔板中,将0.1
×
10
6
个daudi或raji细胞与饱和浓度的抗体(10.0μg/ml)在总体积100μl中室温下在摇床中预温育15分钟。接着,加入25μl正常人血清作为补体来源(20%终浓度),在37℃的培养箱中温育不同的时间,0-60分钟变化。将平板置于冰上以终止反应。加入10μl碘化丙啶并通过facs确定细胞的裂解。
[1233]
图23a显示野生型cd20抗体igg1-7d8显示cdc介导最大杀死为80%的raji细胞,这在测试的条件下5分钟后已经达到。然而,对于igg-7d8-e345r,甚至更快(3分钟后)观察到raji细胞的80%杀死。igg-7d8-e345r的最大裂解(95%)也在5分钟之后达到。
[1234]
图23b显示对于野生型cd20抗体利妥西单抗也同样(该抗体针对所使用的raji细胞诱导cdc的能力比7d8弱),e345r突变的引入导致靶细胞的更快杀死。野生型利妥西单抗显示出cdc介导的最大杀死为32%,这是在20分钟之后达到的。而利妥西单抗-e345r则在大约3分钟之后就达到32%杀死并且显著地,在20分钟后也达到了利妥西单抗的最大裂解(85%)。
[1235]
图23c+d显示能够通过引入e345r突变更快地杀死所使用的raji细胞(该细胞对于野生型cd38抗体igg1-003和igg1-005的cdc介导的杀死具有抗性)。igg1-003-e345r和igg1-005-e345r在5分钟后已经显示出最大cdc(分别是50%和60%)。
[1236]
总而言之,e345r抗体要比它们的野生型相对物更加有效,这是由于更高效力(更低的ec
50
)、增加的最大裂解以及更快的cdc反应动力学的组合造成的。
[1237]
实施例29
[1238]
具有或不具有e345r突变的双特异性抗体的cdc动力学的比较
[1239]
在实施例23中描述了可将e345r突变应用于由duobody平台所产生的cd38
×
cd20双特异性抗体igg1-005-f405l
×
igg1-7d8-k409r,导致增强的杀死能力,如通过在cdc测定法中对于raji和wien133细胞的降低的ec
50
所观察到的。接着,对cdc反应的动力学进行分析以进一步揭示具有或不具有e345r突变的cd38
×
cd20双特异性抗体之间cdc效力的差异。
[1240]
在圆底96孔板中,将0.1
×
10
6
个raji细胞与饱和浓度的抗体(10.0μg/ml)在总体积100μl中室温下在摇床中预温育15分钟。接着,加入25μl正常人血清作为补体来源(20%
终浓度),在37℃的培养箱中温育不同的时间,0-60分钟变化。将平板置于冰上以终止反应。加入10μl碘化丙啶并通过facs确定细胞的裂解。
[1241]
图24显示,双特异性抗体igg1-005-f405l
×
igg1-7d8-k409r诱导了cdc介导的最大杀死为83%,这在10分钟之后达到。e345r突变的引入导致igg1-005-f405l
×
igg1-7d8-k409r所诱导的最大杀死提高(98%),这在2分钟之后就已经达到。这些数据指示在双特异性抗体中引入使fc-fc相互作用稳定的e345r突变导致cdc介导的靶细胞杀死加速。
[1242]
实施例30
[1243]
具有或不具有e345r的单价结合抗体的cdc动力学的比较
[1244]
实施例22显示,单价靶标结合进一步增强e345r抗体的cdc效力,如通过cd38
×
egfr双特异性抗体对cd38阳性、egfr阴性的wien133细胞的最大裂解提高所观察到的。接着,对cdc反应的动力学进行分析以进一步揭示具有或不具有e345r突变的单价结合抗体之间cdc介导的杀死能力的差异。
[1245]
根据如实施例22所述的duobody平台体外产生具有或不具有e345r突变的双特异性cd38
×
egfr和cd20
×
egfr抗体。测试cd38
×
egfr双特异性抗体针对cd38阳性、egfr阴性raji细胞(双特异性抗体只能通过cd38单价地结合该细胞)的cdc效力。在圆底96孔板中,将0.1
×
10
6
个raji细胞与饱和浓度的抗体(10.0μg/ml)在总体积100μl中室温下在摇床中预温育15分钟。接着,加入25μl正常人血清作为补体来源(20%终浓度),在37℃的培养箱中温育不同的时间,0-60分钟变化。将平板置于冰上以终止反应。加入10μl碘化丙啶并通过facs确定细胞的裂解。
[1246]
图25显示,双特异性抗体cd38
×
egfr(igg1-005-k409r
×
igg1-2f8-f405l)诱导了cdc介导的最大杀死为55%,这在大约10分钟后达到。e345r的引入导致最大杀死提高(96%),这在5分钟之内就已经达到。
[1247]
图25显示,双特异性抗体cd20
×
egfr(igg1-7d8-k409r
×
igg1-2f8-f405l)诱导了cdc介导的最大杀死为85%,这在大约5分钟后达到。然而,使用具有e345r引入的cd20
×
egfr抗体,观察到更快的85%裂解(2分钟后)。e345r cd20
×
egfr抗体的最大裂解(97%)同样在5分钟后达到。
[1248]
总而言之,在这些单价结合抗体中引入e345r突变导致更为有力的抗体,这是由于提高的最大裂解以及更快的cdc反应动力学所造成的。
[1249]
实施例31
[1250]
治疗性和e345r/q386k抗体组合的cdc
[1251]
如实施例19中所述,衍生自igg1-005的突变体cd38抗体当野生型抗体的e345位置被除谷氨酸(e)之外的任何其他氨基酸取代时能针对wien133细胞诱导有效的cdc。这意味着,寡聚化(作为cdc的先决条件)由于抗体345位置处存在谷氨酸侧链而被阻碍。由于在六聚抗体环结构中一条fc上的e345紧邻着其对面第二条fc部分的q386,因此有可能通过将第二个抗体的q386位置进行取代而移除第一个抗体中e345-介导的寡聚化阻碍。在两个抗体组合的情况下这将使得第一个抗体的e345能够与第二个抗体的突变386位置更好地相互作用。为了测试这个假设,在wien133上进行cdc测定法,其中将野生型抗体(igg1-003、igg1-005或igg1-11b8)与igg1-005-e345r/q386k或igg1-005-e345r/q386k/e430g混合作为实例。
[1252]
在圆底96孔板中,将0.1
×
10
6
个wien133细胞与系列浓度的未纯化的igg1-005-e345r/q386k、igg1-005-e345r/q386k/e430g或对照抗体(0.0001-20.0μg/ml以3.33倍稀释)在存在或不存在1.0或10.0μg/ml野生型igg1-003、igg1-005或igg1-11b8的情况下在总体积100μl中室温下在摇床中预温育15分钟。接着,加入25μl正常人血清作为补体来源(20%终浓度),在37℃的培养箱中温育45分钟。将平板置于冰上以终止反应。加入10μl碘化丙啶并通过facs确定细胞的裂解。
[1253]
图26a/b/c显示cd38抗体igg1-005-e345r/q386k以剂量依赖的方式诱导wien133细胞的cdc介导的裂解(虚线)。将igg1-005-e345r/q386k与1或10μg/ml野生型cd38抗体igg1-003(图26a)或野生型cd20抗体igg1-11b8(图26b)组合导致提高的最大细胞裂解。将igg1-005-e345r/q386k与野生型igg1-005组合以剂量依赖的方式抑制cdc,可能是通过竞争结合位点实现(图26c)。
[1254]
图26d/e/f显示了cd38抗体igg1-005-e345r/q386k/e430g的相似的结果。
[1255]
这些数据指示,野生型抗体igg1-003和igg1-11b8当与igg1-005-e345r/q386k或igg1-005-e345r/q386k/e430g组合时参与了抗体的寡聚化和cdc激活。在这些组合中,存在于野生型抗体中的由e345-位置造成的寡聚化阻碍能够至少部分被突变抗体中的q386k取代所除去。本申请对改善使用e345位置为野生型的抗体(例如利妥西单抗、奥法木单抗、daratumumab或曲妥单抗)的治疗特别感兴趣。此外,这种寡聚化诱导抗体可能能够促进与直接针对靶细胞(像肿瘤细胞或细菌)的患者自身抗体形成细胞结合复合体。
[1256]
实施例19描述了除e345之外的多个氨基酸(其突变后增强cdc),例如e430和s440,当将它们的特异性突变导入cd38抗体igg1-005时诱导了针对wien133细胞的有效cdc。除了i253和y436突变体,在六聚环结构中所鉴定的寡聚化增强突变与对面第二fc部分的未突变氨基酸相接触。因此,所鉴定的寡聚化增强突变,无论是单独还是组合,预计都能够促进与未突变抗体的寡聚化,并且可通过与实施例19中所应用的相似的选择策略来完成这种突变体的进一步优化。
[1257]
实施例32
[1258]
igg2、igg3和igg4抗体同种型中e345r诱导的cdc
[1259]
为了测试引入促进寡聚化的突变是否能够激发非igg1抗体同种型的cdc活性,通过本领域已知方法来产生具有人igg2、igg3或igg4恒定区的cd38抗体igg1-005同种型变体,生成igg2-005、igg3-005和igg4-005。此外,将增强寡聚化的e345r突变引入所有这些抗体中,生成igg2-005-e345r、igg3-005-e345r和igg4-005-e345r。从cd38抗体igg1-003以类似的方式产生igg2-003和igg2-003-e345r。在体外cdc测定中比较不同同种型的cdc效力。
[1260]
在圆底96孔板中,将0.1
×
10
6
个wien133细胞与10μg/ml的未纯化抗体在总体积100μl中室温下在摇床中预温育15分钟。加入igg1-005-e345r至浓度为3.0μg/ml。接着,加入25μl正常人血清作为补体来源(20%终浓度),在37℃的培养箱中温育45分钟。将平板置于冰上以终止反应。加入10μl碘化丙啶并通过facs确定细胞的裂解。
[1261]
图27显示,igg2-005、igg2-003、igg3-005和igg4-005在所测试的条件下不能裂解(a)daudi或(b)wien133细胞(所观察到~20%的裂解被认为是背景)。e345r突变的引入使得所有所测试的igg同种型的针对daudi细胞的有力cdc。使用针对wien133细胞的cdc证实了这些结果,尽管相对于其他的同种型变体igg3-005-e345r展示出有限的cdc活性。这些数
据指示除了igg1,增强寡聚化的突变,如e345也能用于促进igg2、igg3和igg4抗体的cdc活性。
[1262]
实施例33
[1263]
在离体cdc测定中igg1-005和igg1-005-e345r的针对患者来源cd38阳性b细胞慢性淋巴细胞白血病(cll)细胞的cdc
[1264]
来自于cll患者样本的冷藏保存的原代细胞获自cdb-idibaps-临床医院(dr.elias campo,血液科,病理部,临床医院,institutd
’
investigacions biomediques august pi i sunyer(idibaps),巴塞罗那大学,巴塞罗那,西班牙)的血液病理生物库,或国家心、肺和血液研究所(nhlbi)的临床研究(dr.adrian wiestner,nhlbi,国家健康研究所的血液学分支机构(hematology branch of the naional insitutes of health(nih)),bethesda)。根据临床医院的机构伦理委员会(巴塞罗那,西班牙)或nih的机构审查委员会和赫尔辛基公告,从所有患者处获得知情同意书。所有样本都进行遗传性和免疫表型表征。
[1265]
将cll样本根据如facs所确定的它们的cd38表达分类成两组:5个样品包括于cd38高表达组(daudi细胞上具有50%-98%的cd38表达),4个样品包括于cd38低表达组(daudi细胞上具有0.5%-3%的cd38表达)。
[1266]
将荧光标记的cll细胞(用5μm钙黄绿素(calcein)am标记)与系列浓度的抗体(0.01-10μg/ml以10倍稀释)一起温育。接着,将正常人类血清加入至抗体调理的细胞中(100,000细胞/孔)以作为补体来源(10%终浓度)并在37℃温育45分钟。回收上清液,在synergy
tm ht荧光计数器上读取荧光作为细胞裂解的测量。如下计算细胞杀死:特异性裂解=100
×
(样品自发裂解)/(最大裂解-自发裂解),其中最大裂解由用1%triton处理的细胞样品确定,而自发裂解则由其中将细胞温育于不含抗体的10%nhs中的样品所确定。
[1267]
图28显示,相比起野生型igg1-005,igg1-005-e345r强烈地增强了针对具有高cd38表达的cll原代细胞以及具有低cd38表达的cll原代细胞两者的cdc效力。
[1268]
等同形式
[1269]
只是利用常规的实验本领域技术人员将了解,或者能够确定本文所描述的特定实施方式的许多等同形式。此类等同形式旨在涵盖于所附权利要求中。从属权利要求中所公开的实施方案的任意和全部组合同样意欲在发明的范围之内。
起点商标作为专业知识产权交易平台,可以帮助大家解决很多问题,如果大家想要了解更多知产交易信息请点击 【在线咨询】或添加微信 【19522093243】 与客服一对一沟通,为大家解决相关问题。
与客服一对一沟通,为大家解决相关问题。
此文章来源于网络,如有侵权,请联系删除
相关标签:

 热门咨询
热门咨询




tips