
 商标分类
商标分类  商标转让
商标转让 
抗Aβ治疗性疫苗的制作方法
 2021-01-08 13:01:37|
2021-01-08 13:01:37| 390|
390| 起点商标网
起点商标网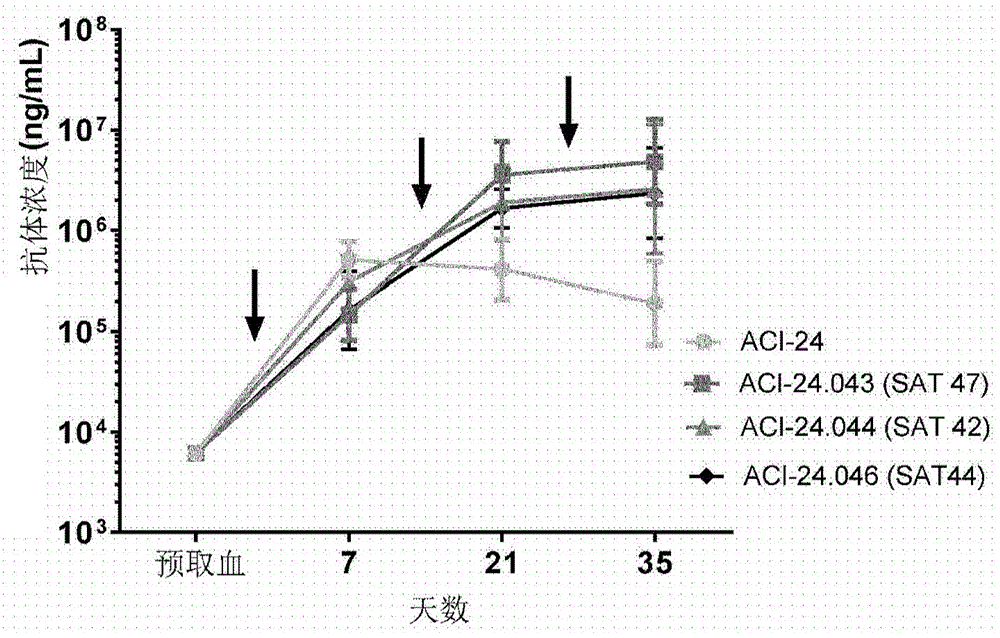
发明涉及抗aβ治疗性疫苗及其在治疗和预防疾病中的用途。疫苗并入了aβ来源的肽b细胞抗原和t细胞表位。
描述
阿尔茨海默病(alzheimer’sdisease,ad)是以认知功能(包括记忆)受损以及进行常规日常活动之能力受损为特征的破坏性、进行性的退行性疾病。ad影响全世界约4,000万患者,随着人口老龄化,该数字迅速提高。ad患者脑中的主要神经病理学变化是神经元死亡,主要发生在记忆和认知相关区域(soto,1999)。ad最显著的病理学特征之一是患病个体的脑中淀粉样蛋白β(amyloidbeta,aβ)斑块的大量存在(soto,1999)。aβ斑块由39至43个氨基酸长的aβ肽形成,该肽在其天然非病理形式中呈无规卷曲构象。在向病理状态转变期间,其主要转变为β-片层二级结构,自发地聚集成不溶性沉积物。
用于ad的少量目前可用的治疗被认为其作用主要是对症的。尽管多年来在开发治疗方面付出了巨大努力,但迄今为止尚未批准用于ad的疾病改善治疗(diseasemodifyingtreatment)。已经进行了尝试以开发可长期中和患病脑中病理性aβ的免疫治疗剂(winblad,2014)。疫苗呈现出刺激免疫系统产生一组略有不同但非常特异性的抗体的优点,而如果需要的话,可通过另外的疫苗接种进一步记起(recall)该应答。然而,针对aβ的主动免疫接种(疫苗接种)方法具有数个主要挑战。淀粉样蛋白β是人体不断暴露于它的所谓的自身抗原。因此,很难打破免疫耐受并诱导针对它的抗体应答。此外,由于免疫系统减弱和免疫细胞数量降低,很难在老年人和病人(例如ad患者)中诱导针对疫苗的强烈免疫应答。
尽管存在这些挑战,但在最初的研究中,全长aβ1-42疫苗(an1792)诱导了抗体应答和有希望的疗效,接受疫苗接种的患者认知下降的速率低于安慰剂治疗的患者(gilman,2005)。然而,6%的被治疗患者发生了脑膜脑炎,这是被认为由t细胞介导的针对全长aβ1-42的应答引起的炎性反应(orgogozo,2003)。
另一种已知的抗aβ疫苗aci-24包含15个氨基酸的序列,其与人aβ序列1-15完全一致(wo2007/068411)。该肽抗原与脂质体载体连接,目的是刺激针对aβ的抗体,同时避免脑膜脑炎和出血(muhs,2007,pihlgren,2013)。选择aβ1-15肽作为抗原是基于该序列含有b细胞表位但缺少全长aβ1-42的强t细胞反应性位点的原理(monsonego,2003),后者被认为是不需要的炎性反应的原因。已显示aci-24通过同时活化对aβ1-15具有特异性的b细胞受体和通过存在于aci-24疫苗中的佐剂单磷酰脂质a(monophosphoryllipida,mpla)活化的toll样受体4(toll-likereceptor4,tlr4)来发挥作用(pihlgren,2013)。通过交联b细胞表面ig受体,b细胞被活化以增殖并产生免疫球蛋白(immunoglobulin,ig)。为了提高抗体产生,可通过由t细胞表位活化的t辅助细胞提供第二信号。由抗原呈递细胞(antigen-presentingcell,apc)表面上的主要组织相容性复合体(majorhistocompatibilitycomplex,mhc)分子(在人中称为人白细胞抗原(humanleucocyteantigen,hla))呈递的t细胞表位促进能够产生ifnγ和il-4的同源t辅助细胞的分化。活化的t细胞与b细胞之间的细胞因子释放和共刺激信号提高抗体应答和类别转换。初次疫苗接种之后,幼稚t细胞增殖并分化成效应细胞。这些细胞中的一小部分将形成长寿记忆t细胞库,在疫苗加强之后重新遇到同源肽时能够快速增殖(sallusto,2010)。所谓的“通用”t细胞表位对绝大多数人群中存在的t细胞是特异性的。它们通常源自人在其一生中通常暴露的抗原(例如破伤风、流感等)。t细胞表位活化t细胞的能力是至少两种互补性质的结果:i)与hla沟(groove)结合的亲和力,其意味着结合的强度,以及ii)其以混杂方式(promiscuousmanner)结合不同hla单倍型的能力,其意味着关于hla分子表达的差异,覆盖非常多样的人群的能力。
需要开发具有高度免疫原性同时保持良好安全特性的抗aβ疫苗。这种需要已通过在脂质体aci-24疫苗内并入通用t细胞表位(universalt-cellepitope)而满足。由于aci-24疫苗在脂质体表面上展示aβ1-15,因此发明人认为在脂质体表面上包含通用t细胞表位是改善疫苗效力的首选途径。然而,出人意料的是,在脂质体表面上包含通用t细胞表位不能提高(或显著提高)疫苗的效力。因此,如本文中所说明的,采用了包封方法,显示其提供了改善的效力。本文中已显示在脂质体疫苗内并入通用t细胞表位提高(或显著提高)了疫苗的效力,同时通过不针对aβ的t细胞活化保持了良好的安全特性。然而,开发这样的方法存在数个挑战。首先,本文中开发的通用t细胞表位倾向于是疏水性的,这使得在脂质体中的包封变得困难。其次,为了改善免疫原性,经常将多种通用t细胞表位组合。然而,随着肽长度的提高,肽合成产率和成功率降低。第三,由于带负电荷的脂质体膜,所选择的通用t细胞表位的电荷影响包封的效率和确保包封所需的实验条件。
因此,本发明提供了脂质体疫苗组合物,其包含:
a.展示在脂质体的表面上的β-淀粉样蛋白(aβ)来源的肽b细胞抗原;以及
b.包封在脂质体内的含有通用t细胞表位的肽。
特别优选的疫苗组合物包含aci-24疫苗,其被修饰以包含包封在脂质体内的含有通用t细胞表位的肽。脂质体是载体的一个实例。因此,载体通常是脂质体,但可以是适合于以与脂质体所实现的相同方式在表面上呈现aβ来源的肽抗原(其中aβ来源的肽抗原采用主要是β-片层的构象)并且还包封含有通用t细胞表位的肽的任何载体。一些实例包括囊泡和颗粒体。
“通用t细胞表位”意指对存在于大多数人群中的t细胞具有特异性的表位。它们通常源自人在其一生中通常暴露的抗原。一些实例包括并入常规施用的疫苗中的抗原。一些具体实例是破伤风、流感和白喉,以及匙孔
包含在本发明的疫苗组合物中的通用t细胞表位通常通过固相合成来合成。因此,在一些实施方案中,通用t细胞表位通过固相合成来合成。包封的这个和其他实际挑战意味着,在一些非限制性实施方案中,含有通用t细胞表位的肽的长度不超过85、80、75或70个氨基酸。确保足够免疫原性的t细胞表位肽的最小长度通常为约10个氨基酸。因此,肽的最小长度通常为约10个氨基酸,以确保产生具有足够免疫原性的t细胞表位。在一些实施方案中,肽的长度为至少20个氨基酸。在另一些实施方案中,肽的长度为30至60个氨基酸;这基于每个通用t细胞表位的优选最小长度和对包含至少两个、三个或四个(连接的)通用t细胞表位的肽的偏好。
还发现,根据本发明实用的通用t细胞表位通常是疏水性的。由于其与脂质的相互作用,这为其合成、纯化和包封在脂质体内提供了进一步的挑战。在包含通用t细胞表位的整个肽中(当考虑整个肽时)或在单独的t细胞表位中(当单独考虑每种通用t细胞表位时),通过用疏水性氨基酸(phe、ile、leu、met、val、trp、ala和pro)的总数除以氨基酸总数并乘以100来计算疏水性百分比。用于本发明目的的疏水性氨基酸定义为亮氨酸(leu)、异亮氨酸(ile)、苯丙氨酸(phe)、色氨酸(trp)、缬氨酸(val)、甲硫氨酸(met)、脯氨酸(pro)和丙氨酸(ala)。
因此,通常来说,含有通用t细胞表位的肽包含至少30%的疏水性氨基酸。这意味着含有通用t细胞表位的整个肽中至少30%的氨基酸是疏水性氨基酸。大多数含有通用t细胞表位的受试肽包含多至50%的疏水性氨基酸。在一些情况下,肽可包含至少35%、40%、45%或50%的疏水性氨基酸。
为了提高免疫原性水平,优选疫苗组合物包含包封在脂质体内的至少两种不同的通用t细胞表位。由于脂质体容量,以及肽的疏水性和合成限制,理想地,每种通用t细胞表位的长度通常不超过30个氨基酸,优选长度不超过20个氨基酸,且更优选地,长度为约10至20个氨基酸。如本文中进一步说明的,发明人已发现较长的通用t细胞表位可被有效修剪至10至20个氨基酸的长度,同时保留免疫原性。基于计算机模拟(insilico)预测的t细胞表位热点,通过在每个单独的t细胞表位的序列中选择最具免疫原性的较短亚序列(通常约15个氨基酸长)来设计经修剪的肽。多种软件程序可用于协助执行此分析,包括epivax免疫原性筛查平台(通过http://www.epivax.com访问)。另外的实例包括syfpeithi(参见hans-georgrammensee,juttabachmann,nielsnikolausemmerich,oskaralexanderbachor,stefanstevanovic:syfpeithi:databaseformhcligandsandpeptidemotifs.immunogenetics(1999)50:213-219;通过http://www.syfpeithi.com访问)、svmhc(httds://www.ncbi.nlm..nih.gov/pubmed/16844990)和iebd数据库(vitar,overtonja,greenbaumja,ponomarenkoj,clarkjd,cantrelljr,wheelerdk,gabbardjl,hixd,settea,petersb.theimmuneepitopedatabase(iedb)3.0.nucleicacidsres.2014oct9.pii:gku938.[epubaheadofprint]pubmedpmid:25300482;通过http://www.iedb.org/访问)。
在一些实施方案中,每种通用t细胞表位包含至少30%的疏水性氨基酸。这意味着单独的通用t细胞表位中至少30%的氨基酸是疏水性氨基酸。对于特定的表位,该数值可高达80%的疏水性氨基酸。在一些情况下,可存在至少40%、45%、50%、55%、60%、65%、70%、75%或80%的疏水性氨基酸。在一些实施方案中,最大值可以是80%的疏水性氨基酸,这意味着最宽范围可以是30%至80%的疏水性氨基酸。
为了平衡改善的免疫原性与包封的实际挑战,疫苗组合物可包含包封在载体内的两个、三个或四个不同的通用t细胞表位。当包封大量不同的通用t细胞表位(尤其是3或4个)时,优选将它们修剪成约10至20个氨基酸,例如约15个氨基酸的长度。优选多个不同的通用t细胞表位包含在包封的相同肽中。因此,包含多个不同的通用t细胞表位的合成肽构建体代表了本发明的一种优选实施。在某些实施方案中,肽包含至少两个不同的通用t细胞表位。在一些更具体的实施方案中,肽包含两个、三个或四个通用t细胞表位。当合成肽构建体中包含至少两个通用t细胞表位时,它们可通过接头连接。接头用于以不损害所连接表位之免疫原性的方式将通用t细胞表位彼此物理连接。用于将氨基酸彼此连接的合适接头是本领域中公知的。优选的接头本身是基于氨基酸的接头,即肽接头。因此,它们可通过肽键将通用t细胞表位彼此连接。接头是使得能够正确处理通用t细胞表位的接头。mhcii类分子的抗原呈递需要抗原进入到内体-溶酶体区室中。然后通过蛋白水解酶处理这些抗原,其中木瓜蛋白酶家族的溶酶体半胱氨酸蛋白酶构成重要的子集。产生的肽与mhcii类分子结合,其随后在专职抗原呈递细胞(apc)(包括巨噬细胞、树突细胞(dendriticcell,dc)和b细胞)的表面展示(lutznerandkalbacher2008)。因此,优选地,接头包含木瓜蛋白酶家族的溶酶体半胱氨酸蛋白酶的底物。接头可包含组织蛋白酶s、组织蛋白酶b和组织蛋白酶l中一种或更多种的底物。在一些实施方案中,接头包含至少两个或至少三个氨基,基本上由其组成或由其组成。在一些实施方案中,接头包含氨基酸vvr、tvglr、kvsvr、pmgap或pmglp,基本上由其组成或由其组成。
因此,包含两个通用t细胞表位的肽可以是以下形式的线性肽:
[通用t细胞表位1]-[接头]-[通用t细胞表位2]
因此,包含三个通用t细胞表位的肽可以是以下形式的线性肽:
[通用t细胞表位1]-[接头]-[通用t细胞表位2]-[接头]-[通用t细胞表位3]
因此,包含四个通用t细胞表位的肽可以是以下形式的线性肽:
[通用t细胞表位1]-[接头]-[通用t细胞表位2]-[接头]-[通用t细胞表位3]-[接头]-[通用t细胞表位4]
应注意,在每对连接的通用t细胞表位之间接头不必相同。因此,例如,通用t细胞表位1与通用t细胞表位2之间的接头可以不同于通用t细胞表位2与通用t细胞表位3之间的接头。在四个通用t细胞表位的情况下,三个接头中的每一个可以不同,或者两个可以相同,而第三个不同(以任何顺序)。在其中在肽中包含多个接头的一些实施方案中,它们都是相同的。
当设计用于包封的合适肽时,本发明入筛选了一系列通用t细胞表位来源。在一些实施方案中,通用t细胞表位来源自白喉毒素、破伤风毒素、eb病毒(epsteinbarrvirus)、流感血凝素和/或匙孔
a.白喉毒素和破伤风毒素通用t细胞表位的组合
b.eb病毒和破伤风毒素通用t细胞表位的组合
c.eb病毒、破伤风毒素和匙孔
d.流感血凝素、白喉毒素、破伤风毒素和eb病毒通用t细胞表位的组合。
这样的组合优选以如上说明的接头形式提供。为了避免疑问,虽然所述组合优选地以指定顺序包括在内,但是其可以以替代的顺序包括在内。例如,如果存在三个通用t细胞表位,a、b和c,则它们可以以顺序abc、acb、bac、bca、cab或cba中的任一种包括在内。
包含多个不同的通用t细胞表位的特定肽构成本发明的另一方面。这样的肽优选包含在本发明的疫苗组合物中。因此,可用于本发明的肽包含选自以下的氨基酸序列、基本上由其组成或由其组成:seqidno:1(sat42)、seqidno:2(sat43)、seqidno:3(sat44)、seqidno:4(sat47)。参照下表2更详细地说明这些肽的组成。
包含单种通用t细胞表位的特定肽也构成本发明的另一方面。这样的肽优选包含在本发明的疫苗组合物中。因此,可用于本发明的肽包含选自以下的氨基酸序列、基本上由其组成或由其组成:seqidno:5(sat6)、seqidno:6(sat13)、seqidno:7(sat15)、seqidno:8(sat17)。参照下表1更详细地说明了这些肽的组成。适当时修剪至10至20个氨基酸长度的这些肽的组合也可包含在本发明的疫苗组合物中。组合的肽优选通过本文中定义的一个或更多个接头连接。
aβ来源的肽抗原展示在脂质体的表面上。这通常通过插入到脂质体的外表面中。通过将aβ来源的肽抗原连接至插入到脂质体外表面中的部分,可促进插入到脂质体的外表面中。脂质体可以是适于在表面上呈现aβ来源的肽抗原并且还包封含有通用t细胞表位之肽的任何脂质体。通常来说,所述部分包含疏水性部分以确保插入到脂质体的脂质双层中。所述部分可以是任何合适的部分,但优选是脂肪酸。脂肪酸可包含棕榈酰残基。优选的构建(如在aci-24中)包含连接至肽的n端和c端区域中的两个棕榈酰残基的aβ来源的肽抗原(aci-24中aβ(1-15))。因此,肽抗原是四棕榈酰化的。这可通过在aβ来源的肽抗原的n端和c端区域中并入两个赖氨酸残基来促进。赖氨酸残基是棕榈酰化的。
在一些实施方案中,脂质体具有负表面电荷;脂质体是阴离子性的。优选地,脂质体包含磷脂,并且甚至更优选地,磷脂包含二豆蔻酰磷脂酰胆碱(dimyrsitoylphosphatidyl-choline,dmpc)和二豆蔻酰磷脂酰甘油(dimyrsitoylphosphatidyl-glycerol,dmpg)。脂质体可还包含胆固醇。在一些实施方案中,这三种组分的摩尔比可以是9∶1∶7。
因此,最优选的构建包含在脂质体中重构的aβ来源的肽抗原。因此,本发明的这些组合物在本文中通常可称为“本发明的脂质体疫苗组合物”。
aβ来源的肽抗原在对象中诱导b细胞应答。其为“b细胞抗原”。如已经说明的,aβ斑块由39至43个氨基酸长的aβ肽形成,该肽在其天然非病理形式中呈无规卷曲构象。在向病理状态转变期间,其主要转变为β-片层二级结构,自发地聚集成不溶性沉积物。因此,本文中将aβ来源的肽抗原限定为来源于aβ的(最多)43个氨基酸,但不是全长aβ的肽抗原。更具体地,aβ来源的肽抗原包括aβ(1-42)的免疫显性(immunodominant)b细胞表位,但缺少aβ(1-42)中存在的t细胞表位。因此,在一些实施方案中,aβ来源的肽抗原包含来自aβ的n端17个氨基酸的13至15个连续氨基酸、基本上由其组成或由其组成。应当注意,aβ来源的肽抗原可在较大肽分子的背景下提供,较大肽分子的其余部分不来源于aβ氨基酸序列。例如,肽可包含另外的残基(例如赖氨酸残基)以促进棕榈酰化。这些残基通常存在于肽的n端和c端。在这种情况下,术语“基本上由……组成”意指aβ来源的肽抗原包含来自aβ的n端17个氨基酸的13至15个连续氨基酸,但可包含有限数量的另外的残基(例如4个赖氨酸残基)以促进棕榈酰化。优选的aβ来源的肽抗原包含aβ的第1至15位氨基酸(其可称为“aβ(1-15)”(wo2007/068411,aci-24))、基本上由其组成或由其组成。
包含在本发明组合物中的aβ来源的肽抗原采用复制aβ病理形式的二级结构。优选地,aβ来源的肽抗原采用包含β-片层构象的二级结构。甚至更优选地,当展示在脂质体的表面上时,aβ来源的肽抗原主要采用β-片层构象。
本发明的组合物通常包含至少一种佐剂。在本发明的一些实施方案中,本发明的组合物包含两种佐剂。佐剂的目的是提高或刺激对象的免疫应答。优选地,所述至少一种佐剂是载体的一部分(与包封在载体内相反)。因此,所述至少一种佐剂可形成脂质体的一部分;其可形成脂质双层的一部分。因此,佐剂可以是基于脂质的佐剂。佐剂可至少部分地展示在脂质体的表面上;这可以是由于佐剂形成脂质双层的一部分。在一些实施方案中,形成脂质体一部分的一种或更多种佐剂可与包封的佐剂组合。在另一些实施方案中,当形成脂质体时,形成脂质体一部分的一种或更多种佐剂可与另外的佐剂(例如alum或cpg)混合。载体(脂质体)可用作佐剂,其中向脂质体添加单磷酰脂质a(mpla),该术语涵盖mpla衍生物,例如(合成)3-脱酰基单磷酰基六酰基脂质a(monophosphorylhexa-acyllipida,3-deacyl(synthetic))(3d-(6-酰基)
可根据本发明使用的其他佐剂包括氢氧化铝(alum)和/或cpg等。
将本发明的疫苗组合物施用于对象,以治疗以下、预防以下、诱导针对以下的保护性免疫应答或减轻与以下相关的症状:淀粉样蛋白-β相关疾病或病症或者以认知记忆能力受损为特征或与其相关的病症。因此,疫苗组合物可具有预防性应用和治疗性应用这二者。对象是哺乳动物,且通常是人。
淀粉样蛋白-β相关疾病或病症可以是神经病症,例如阿尔茨海默病(ad)。根据本发明的淀粉样蛋白-β相关疾病或病症的其他实例包括轻度认知损伤(mildcognitiveimpairment,mci)、唐氏综合征(down(’s)syndrome)、心脏淀粉样变性、脑淀粉样血管病(cerebralamyloidangiopathy,caa)、多发性硬化、帕金森病(parkinson’sdisease)、路易体痴呆(lewybodydementia)、als(肌萎缩侧索硬化)、成年型糖尿病(adultonsetdiabetes)、包涵体肌炎(inclusionbodymyositis,ibm)、眼部淀粉样变性、青光眼、黄斑变性、网络状营养不良(latticedystrophy)和视神经炎。这些病症中的许多以认知记忆能力受损为特征或与其相关。因此,根据本发明以认知记忆能力受损为特征或与其相关的病症包括ad、轻度认知损伤(mci)、唐氏综合征、心脏淀粉样变性、脑淀粉样血管病(caa)、多发性硬化、帕金森病、路易体痴呆、als(肌萎缩侧索硬化)和包涵体肌炎(ibm)。
因此,本发明提供了在对象中治疗以下、预防以下、诱导针对以下的保护性免疫应答或减轻与以下相关的症状的方法:淀粉样蛋白-β相关疾病或病症或者以认知记忆能力受损为特征或与其相关的病症,所述方法包括向对象施用本发明的疫苗组合物。
这样的方法也可以以本发明疫苗组合物的医学用途的形式表示。因此,本发明还提供了本发明的疫苗组合物,其用于在对象中治疗以下、预防以下、诱导针对以下的保护性免疫应答或减轻与以下相关的症状:淀粉样蛋白-β相关疾病或病症或者以认知记忆能力受损为特征或与其相关的病症。
类似地,本发明提供了本发明的疫苗组合物在制备药物中的用途,所述药物用于在对象中治疗以下、预防以下、诱导针对以下的保护性免疫应答或减轻与以下相关的症状:淀粉样蛋白-β相关疾病或病症或者以认知记忆能力受损为特征或与其相关的病症。
本文中的所有实施方案均适用于这样的方法或医学用途,无论其以何种方式表示。向对象施用本发明的疫苗组合物导致产生与aβ的病理形式结合的通常为多克隆的igg抗体。如已经说明的那样,aβ的那些病理形式包含β-片层多聚体。所产生的抗体可因此被称为“aβ特异性”抗体。
抗体结合靶抗原的能力主要受两个参数调节:亲和力(affinity)和亲合力(avidity)。抗体的亲和力测量抗体与其抗原之间的单价相互作用的强度。抗体亲合力包括通过抗原与抗体之间的多于一个相互作用点来增强结合。由疫苗接种诱导的多克隆血清的结合能力依赖于上述两个参数(siegrist,2013)。其通常被称为多克隆应答的亲合力,因为很难独立地评价亲和力和亲合力。如本文中进一步详细说明的,参见实施例4(4.2节),发明人开发了elisa测定,其中平行评价含有多克隆抗体的血清与较低和较高浓度抗原的总体结合(martineau,2010)。低包被信号和高包被信号之间的比值(信号表示结合抗体的浓度)表示为亲合力指数。与较低的指数评分(接近0)相比,较高的指数评分(接近1)表明改善的总体结合强度。亲合力指数随时间的提高提供了疫苗诱导的抗体的总体亲合力成熟的指示。本文中显示(参见实施例4和图4),与使用aci-24(没有含有通用t细胞表位的包封的肽)的免疫接种相比,使用本发明疫苗组合物(其包含含有通用t细胞表位的包封的肽)的免疫接种产生改善的成熟效果。
本发明的疫苗组合物可通过任何适当的施用途径施用于对象。如技术人员会知晓的,疫苗组合物可通过表面(topical)、经口、经直肠、经鼻或肠胃外(例如静脉内、皮内、皮下或肌内)途径施用。另外,疫苗组合物可以并入持续释放基质中,例如生物可降解的聚合物中,将聚合物植入期望递送的地方附近或邻近。然而,在一些优选的实施方案中,疫苗组合物肌内或皮下施用。
本发明的疫苗组合物可单次施用于对象以产生保护性免疫应答。然而,在一些实施方案中,将本发明的疫苗组合物多次施用于同一对象。因此,根据本发明可采用所谓的致敏-加强方案(prime-boostregimen)。疫苗的施用通常间隔至少1周、且通常为约1至12个月的间隔期。不希望受特定假设的束缚,可能的是,通过向aci-24添加通用t细胞表位通过提供来自对同源t细胞表位具有特异性的活化t细胞的第二信号来增强抗aβ抗体应答。本发明的疫苗组合物代表了用于预防和治疗淀粉样蛋白-β相关疾病或病症(例如ad)的强有力的新治疗选择。在一些实施方案中,每次施用相同的疫苗组合物——同源疫苗接种(homologousvaccination)方案。同源疫苗接种是指使用相同疫苗进行致敏(第一次免疫接种)和加强(第二次或任何进一步的免疫接种)的免疫接种方案。
在另一方面,异源致敏-加强免疫接种需要在首次和至少一些随后的免疫接种中使用不同的疫苗。在一些实施方案中,本发明的疫苗组合物在与其他“抗aβ”疫苗的异源致敏-加强组合中多次施用于同一对象,所述其他“抗aβ”疫苗携带来自aβ蛋白的任何部分的肽抗原,其可包括来自aβ(1-15)区域外部的肽抗原。在一些实施方案中,本发明的疫苗组合物在与其他“抗aβ”疫苗的异源致敏-加强组合中多次施用于同一对象,所述其他“抗aβ”疫苗携带与本发明的脂质体疫苗组合物中包含的相同的肽抗原,其可包括aβ(1-15)肽抗原。在一些实施方案中,本发明的疫苗组合物(优选包含aβ(1-15)肽抗原)在与其他“抗aβ”疫苗的异源致敏-加强组合中多次施用于同一对象,所述其他“抗aβ”疫苗携带相应的aβ来源的肽抗原,优选aβ(1-15)肽抗原。可在异源致敏-加强疫苗接种中与本发明包含aβ来源的抗原的疫苗组合物一起施用的“抗aβ”疫苗的一些实例包括但不限于aβ(1-15)-padre疫苗(agadjanyanetal.,2005;ghochikyanetal.,2006)、aβ(1-15)-白喉类毒素(diphtheriatoxoid,dt)或crm疫苗(wo2010016912)、赖氨酸连接的aβ(1-15)的串联重复(maieretal.,2006)、树枝状aβ(1-15)疫苗(seabrooketal.,2006)、aβ(1-15)dt缀合物(liuetal.,2013)、偶联至噬菌体qβ包被蛋白的aβ(1-6)(windbladetal.2012)、aβ(1-7)-crm(araietal.2015)、ntermaβ-klh(schneebergeretal2010)。
本发明还提供了包含根据本发明的疫苗组合物的药盒(kit)。因此,提供了用于在对象中治疗以下、预防以下、诱导针对以下的保护性免疫应答或减轻与以下相关的症状的药盒:淀粉样蛋白-β相关疾病或病症或者以认知记忆能力受损为特征或与其相关的病症,所述药盒包含如本文中所述的本发明的(脂质体)疫苗组合物。可为这样的药盒提供合适的使用说明书。使用说明书可说明组合物的施用方案。因此,药盒可包含多个(分开)剂量的本发明疫苗组合物。使用说明书可进一步说明组合物的储存条件,特别是在施用疫苗组合物剂量之间的时间段内。这些药盒可应用于本文中公开的本发明的所有相关方法。
本发明还提供了用于产生本发明脂质体疫苗组合物的方法。这样的方法可包括以下步骤:
a.产生脂质膜
b.将脂质膜在包含含有通用t细胞表位的肽的缓冲液中再水化
c.从再水化的脂质膜产生脂质体,其包封含有通用t细胞表位的肽以形成包含脂质体的溶液,所述脂质体包含包封的通用t细胞表位
d.向溶液添加β-淀粉样蛋白(aβ)来源的肽抗原,并将溶液保持在导致β-淀粉样蛋白(aβ)来源的肽抗原插入到脂质体脂质双层中的条件下。
这样的方法在本文中举例说明,其细节可应用于本发明的这些方面。一般而言,该方法可涉及薄脂质膜形成,然后均质化和挤出。因此,在一些实施方案中,通过将脂质溶解在乙醇中并随后在真空下蒸发乙醇来产生脂质膜。优选的脂质组分在关于本发明的脂质体疫苗组合物中进行了说明,并且包括dmpc、dmpg、胆固醇和mpla(作为佐剂)。这些组分的摩尔比可以是9∶1∶7∶0.05。这样的摩尔比也适用于本发明的脂质体疫苗组合物。脂质组分可需要在升高的温度下溶解。升高的温度可以是40℃至80℃,例如约60℃。
在步骤b中,用于再水化的缓冲液可取决于使用哪一种含有通用t细胞表位的肽。通常来说,可使用任何合适的缓冲液。在一些实施方案中,缓冲液包含乙酸钠或pbs。如果要包封sat42,则缓冲液可以是乙酸钠。如果要包封sat43、sat44或sat47中的任一种或更多种,则缓冲液可以是pbs。在所有情况下,可将dmso(例如5%dmso)添加至缓冲液。可在搅拌样品的情况下进行再水化。
在步骤c中,可通过在珠的存在下进行涡旋来产生脂质体。可使用任何合适的珠。珠可以是例如玻璃珠。该步骤可产生多层囊泡,其随后转化为包含脂质双层的脂质体。这种转化可依赖于多个(例如5至15个,优选10个)冻融循环。冻融循环之后可进行均质化。这之后可进行基于尺寸的挤出。在一些实施方案中,通过约0.08至0.1μm直径(或最大尺寸)的孔挤出脂质体。这可通过例如聚碳酸酯膜的膜。挤出的脂质体可被浓缩,例如使用过滤形式(例如超滤)。
步骤d导致β-淀粉样蛋白(aβ)来源的肽抗原插入到脂质体的脂质双层中。必要条件可包括在25至35℃(例如约30℃)的温度下搅拌10至60分钟(例如约30分钟)。优选的β-淀粉样蛋白(aβ)来源的肽抗原是包含aβ1-15的四棕榈酰化肽。该肽在任一端包含两个赖氨酸残基以产生四棕榈酰化肽。在注入到脂质体溶液中之前,肽可预先溶解在磷酸氢二钠中。
该方法还可包括作为最后步骤的将疫苗组合物过滤。这可在无菌条件下。过滤可通过孔径为0.2μm的膜。合适的膜包括聚醚砜(polyethersulfone,pes)膜,其可以以注射器过滤器的形式提供。然后可将所产生的疫苗组合物储存在合适的条件下直至使用,例如在冷藏(例如约5℃)下。
用于产生本发明脂质体疫苗组合物的替代方法可依赖于如本文中所例示的横流注射(crossflowinjection)。因此,本发明还提供了用于通过横流注射产生本发明脂质体疫苗组合物的方法。这些方法可特别应用于包封sat44或sat47的组合物。这样的方法可包括以下步骤:
a.将形成脂质体的脂质(和佐剂,如果是基于脂质的话)溶解在溶液中
b.将含有通用t细胞表位的肽溶解在溶液中
c.使用横流注射模块混合来自步骤a.和b.的溶液以形成包封所述含有通用t细胞表位的肽的中间脂质体
d.将中间脂质体通过膜挤出以减小其尺寸和多分散性
e.使用横流注射模块将包含β-淀粉样蛋白(aβ)来源的肽抗原的溶液与来自步骤d的溶液混合,使得β-淀粉样蛋白(aβ)来源的肽抗原插入到所述脂质体的脂质双层中。
这样的方法在本文中例示,其细节可应用于本发明的这些方面。一般而言,该方法使用横流注射来包封含有通用t细胞表位的肽,并将β-淀粉样蛋白(aβ)来源的肽抗原插入到脂质体的脂质双层中。
在步骤a中,脂质(其可包含佐剂,例如本文中所述的mpla佐剂)通常溶解在乙醇中。乙醇可以是90%至100%乙醇,例如96%乙醇。可通过加热(例如加热到40至80℃,例如约60℃的温度)来加速溶解。针对本发明的脂质体疫苗组合物说明了一些优选的脂质组分,并且其包括dmpc、dmpg、胆固醇和mpla(作为佐剂)。这些组分的摩尔比可以是9∶1∶7∶0.05。这样的摩尔比也适用于本发明的脂质体疫苗组合物。
在步骤b中,溶解含有通用t细胞表位的肽。在一些实施方案中,可借助于搅拌(例如声处理)将肽溶解在合适的缓冲液(例如his-蔗糖缓冲液)中。
在步骤c中,使用横流注射模块将来自步骤a和b的溶液混合以形成包封含有通用t细胞表位的肽的中间脂质体。在该步骤之前,可过滤来自步骤a和b的溶液。过滤器的合适孔径可以是约0.2μm。溶液可以以任何合适的浓度使用。一旦过滤,可将溶液加热至30至60℃(例如约40℃)的温度。通过将两种溶液(来自步骤a和b)注射通过横流模块(其中2种溶液相遇)形成脂质体。这通常在特定的流量和温度下进行,如技术人员容易理解的(合适的温度如上所述)。在一些实施方案中,在脂质体形成之后,通常可添加缓冲液以降低乙醇浓度。可使用任何合适的缓冲液,例如his-蔗糖缓冲液。
在步骤d中,将中间脂质体通过膜挤出以减小其尺寸和多分散性。溶液中的所形成脂质体包封含有通用t细胞表位的肽。可使用任何合适的膜。合适的孔径可以是约100nm。合适的膜类型是聚碳酸酯膜。该步骤可在任何合适的温度下进行,优选在室温(例如约25℃)下进行。在该步骤之后,可进行过滤步骤(例如超滤/渗滤)以除去乙醇。任何合适的膜可用于该步骤,例如截留分子量为约500kd的中空纤维膜。可进行进入到分散缓冲液中的缓冲液交换步骤。优选的分散缓冲液是pbs。pbs可处于合适的ph,例如6至8,特别是约6.9。这可需要5至15(例如约10)体积交换。在步骤e之前,可将脂质体在分散缓冲液中稀释至期望浓度。期望浓度可以是0.1至10mg/ml,例如约1mg/ml。在步骤e之前,可将含脂质体的溶液加热至合适的温度,例如30至60℃,优选约35℃。
步骤e涉及使用横流注射模块将包含β-淀粉样蛋白(aβ)来源的肽抗原的溶液与来自步骤d的溶液混合。如本文中所讨论的,β-淀粉样蛋白(aβ)来源的肽抗原优选是脂质化的(例如四棕榈酰化的),该讨论适用于必要的变更(mutatismutandis)。在混合之前,通常将β-淀粉样蛋白(aβ)来源的肽抗原溶解在合适的缓冲溶液中,例如β-og在10mmna2hpo4ph11.4缓冲液中的10%w/v的溶液。通常将溶液加热至合适的温度,例如30至80℃(例如约60℃)的温度。可根据需要进一步稀释溶液以确保β-淀粉样蛋白(aβ)来源的肽抗原的合适的浓度。合适的浓度可以是0.1至10mg/ml,例如约1mg/ml。ph通常保持11至12,例如约11,优选11.4。使用横流注射模块将包含β-淀粉样蛋白(aβ)来源的肽抗原的溶液与来自步骤d的溶液混合,使得β-淀粉样蛋白(aβ)来源的肽抗原插入到脂质体的外脂质双层中。可将混合物在合适的温度下孵育固定的一段时间,以促进β-淀粉样蛋白(aβ)来源的肽抗原插入到脂质体的脂质双层中。合适的时间段可以是20至120分钟,例如约30分钟。合适的温度可以是30至60℃,优选约35℃。孵育可在搅动(例如搅拌)下进行。
在步骤e之后,产物可被回收以包含在本发明的组合物中。因此可将产物配制成本发明的脂质体疫苗组合物。这可涉及超滤/渗滤步骤,以从缓冲溶液中除去β-og。任何合适的膜可用于该步骤,例如截留分子量为约500kd的中空纤维膜。超滤/渗滤可涉及进入到最终缓冲液中的缓冲液交换步骤。优选的最终缓冲液是his-蔗糖缓冲液,其可以是10mm组氨酸、250mm蔗糖。这可需要5至15(例如约10)体积交换。可进行浓缩步骤以获得优选的最终体积。还可进行最终(无菌)过滤步骤。这可使用筒式过滤器。过滤步骤可通过具有任何合适孔径(例如约0.2μm)的过滤器。过滤可在无菌条件下进行。然后可将所产生的疫苗组合物储存在合适的条件下直至使用,例如在冷藏(例如约5℃)下。
附图说明
图1.(a)在用指定的疫苗第1次免疫接种(箭头指示免疫接种时间点)之前(预取血(prebleeding))21天(aci-24.046)或7天(aci-24、aci-24.043、aci-24.044)以及之后7天、21天和35天,c57bl/6小鼠血浆中通过elisa对aβ1-42特异性igg抗体的分析。结果表示为每组n=5只小鼠的ng/ml的几何平均值+/-95%置信区间(confidenceinterval,ci)。x轴表示处理/取血的天数,而y轴表示以ng/ml表示的抗体效价。(b)在用指定的疫苗第1次免疫接种之前(预取血)21天(aci-24.046)或7天(aci-24、aci-24.043、aci-24.044)以及之后21天,c57bl/6小鼠血浆中通过elisa对aβ1-42特异性igg抗体的分析。结果表示为每组5只小鼠的ng/ml的几何平均值+/-95%ci。第21天不同组间的统计学检验:kruskal-wallis检验和dunn多重比较。*p<0.05;**p<0.01。x轴表示来自用指定疫苗免疫接种的组的个体血浆,而y轴表示以ng/ml表示的抗体效价。
图2.(a)在用指定疫苗第1次免疫接种之前21天或7天(虚线)和之后21天(实线),c57bl/6小鼠血浆中通过igg抗体的elisa对aβ1-42自缔合抑制的分析。结果表示为aβ1-42自缔合抑制百分比的每组5只小鼠的平均值+/-标准偏差。x轴表示系列稀释的血浆,而y轴表示aβ1-42自缔合的抑制百分比。(b)aβ1-42自缔合的抑制表示为1/25稀释的血浆中第21天的抑制百分比(%)减去第-21天或第-7天(背景——预取血)的抑制%。x轴表示用指定疫苗处理的组,而y轴表示减去背景之后aβ1-42自缔合的抑制百分比。
图3.在用指定的疫苗第1次免疫接种之前21天(aci-24.046)或7天(aci-24、aci-24.043、aci-24.044)以及之后21天,c57bl/6小鼠血浆中通过elisa对aβ1-42寡聚体特异性igg抗体的分析。结果表示为每组5只小鼠的ng/ml的几何平均值+/-95%ci。第21天组间的统计学检验:kruskal-wallis检验和dunn多重比较。*p<0.05;**p<0.01。x轴表示用指定疫苗免疫接种的组,而y轴表示以ng/ml表示的抗体效价。
图4.在用指定疫苗第一次免疫接种之后7和21天,c57bl/6小鼠血浆中通过elisa对igg抗体的aβ1-42亲合力的分析。结果表示为每组5只小鼠的亲合力指数的几何平均值+/-95%ci。每个组在第7天与第21天之间的统计学检验:曼恩-惠特尼检验(mann-whitneytest)。*p<0.05;**p<0.01。x轴表示用指定疫苗免疫接种的组,而y轴表示亲合力指数。
图5.在aci-24.046(sat44,n=8)、aci-24.045(sat43,n=4)或aci-24.043(sat47,n=4)免疫接种的猴中,第1次免疫接种之前(第1天)和第3次免疫接种之后1周(第64天),食蟹猴(cynomolgusmonkey)血清中通过msd对aβ寡聚体特异性igg抗体的分析。结果表示为au/ml的几何平均值+/-95%ci。x轴表示来自用指定疫苗免疫接种的组的个体血浆,而y轴表示以au/ml表示的抗体效价。
图6.在用aci-24和aci-24.046(sat44)疫苗(a)或用aci-24和aci-24.043(sat47)疫苗(b)第3次免疫接种之后7天(第36天),c57bl/6小鼠血浆中通过elisa对aβ1-42特异性igg抗体的分析。结果表示为每组n=10只小鼠的ng/ml的几何平均值+/-95%ci。x轴表示用于每个特定组免疫接种的疫苗,而y轴表示以ng/ml表示的抗体效价。aci-24与指定疫苗之间的统计学检验:曼恩-惠特尼检验。*p<0.05;**p<0.01,***p<0.001。
缩写表
将参照以下非限制性实施例进一步理解本发明:
实施例1.新t细胞表位的设计
t细胞表位活化t细胞的能力(免疫原性评分)是两种互补性质的结果:i)对hla的亲和力和ii)以混杂方式结合不同hla单倍型的能力。进行来自不同来源的数种t细胞表位的计算机模拟评价(epivax),目的是选择具有最高免疫原性评分的肽。在初步阶段,评价了来自不同来源(匙孔
表1.
根据单独肽的筛选结果,设计了由来自不同来源的2或3种免疫原性t细胞表位构成的组合的混杂肽(命名为sat42、sat43和sat44)和由经修剪的肽构成的混杂肽(例如sat47和sat43)(表2)。基于计算机模拟预测的t细胞表位热点,通过在每个单独的t细胞表位的序列中选择最具免疫原性的15聚体肽序列来设计经修剪的肽。由于肽合成和疫苗包封过程的限制,目的是在不提高最终混杂肽的大小的情况下提高免疫原性评分。简单地说,在肽长度提高时,尤其是长度超过30个氨基酸时,肽合成产率和成功率降低,且此外,其主要由疏水性残基构成,如对于本文中公开的t细胞表位肽。此外,肽包封率随着肽长度的提高而降低,这是因为随着肽长度的提高,将其容纳在脂质体腔中的机会降低。这4种混杂t细胞表位的计算机模拟免疫原性评分非常高,并且重要地是高于单独组分肽,因此证实组合来自不同来源的肽可改善hla亲和力和hla单倍型覆盖度(混杂的t细胞表位序列示于表2中)。
表2.
实施例2.疫苗合成和配制
通用t细胞表位肽合成和纯化的通用方法
使用标准fmoc化学,通过线性固相肽合成(solidphasepeptidesynthesis,spps)在2-氯三苯甲基树脂上制备t细胞肽。使用3.0当量的氨基酸和偶联试剂,在dmf中的3.0当量的碱存在下在室温下进行标准偶联操作1小时。对于难以偶联的序列,实施双偶联,并延长反应时间。在氨基酸偶联完成之后,使用吡啶中的5.0当量的ac2o引入乙酰化封端步骤以中止不希望的肽链延伸。用dmf洗涤树脂,并通过使用dmf中的20%哌啶去除fmoc基团5分钟。在完成spps之后,使用标准切割混合物(tfa/tis/水)在室温下进行树脂的总体脱保护和肽切割2小时。滤出树脂并用tfa洗涤。随后用10倍过量体积的冷异丙醚/己烷沉淀粗制产物,通过使用玻璃料(glassfrit)滤出固体并在真空下干燥。在制备型hplc系统上,使用溶剂a(水,0.1%tfa)和溶剂b(乙腈,0.1%tfa)的梯度在反相c18柱上纯化粗制肽。将含有纯度高于90%的期望肽的hplc级分合并在一起,在水中稀释并进行离子交换。将期望的离子交换级分冻干。通过hplc-ms分析表征并确认最终肽的身份(identity)和纯度。
aci-24.043/aci-24.044/aci-24.045/aci-24.046/疫苗的制备(薄脂质膜)
通过薄脂质膜技术产生含有包封的t细胞表位肽的疫苗,然后均质化并挤出。首先,通过在60℃下以9:1:7:0.05的摩尔比分别将dmpc、dmpg(lipoid,germany)、胆固醇和合成3-脱酰基单磷酰基六酰基脂质a或3d-(6-酰基)phadtm(avantipolarlipids,usa)溶解在乙醇中。在真空旋转蒸发仪下蒸发乙醇以获得薄脂质膜。
用这些缓冲液之一将脂质膜再水化(取决于待包封的t细胞表位肽):
·milliq水中的20mm乙酸钠ph4(fluka),5%dmso(sigmaaldrich),含有0.8mg/ml的t细胞表位肽sat42,或
·milliq水中的0.1×pbsph7.4,5%dmso(全部为sigma-aldrich),含有0.3至0.4mg/mlt细胞表位肽sat43、sat44或sat47。
将溶液轻轻搅拌15分钟。将样品在玻璃珠的存在下进一步剧烈涡旋。将得到的多层囊泡进行10个冻融循环(液n2和37℃的水浴)并进行均质化,然后通过孔径为0.1/0.08μm的聚碳酸酯膜(whatman,uk)连续挤出。使用emulsiflex-c5(avestin,canada)进行均质化和挤出这两个步骤。将挤出的脂质体通过超滤浓缩,并通过渗滤(10次交换)将缓冲液交换为pbsph7.4。在pal1-15添加之前,将得到的脂质体在pbsph7.4中稀释并加热至30℃。
将四棕榈酰化的人肽pal1-15(bachemag,switzerland)溶解在含有1%β-og(sigma-aldrich,usa)的milliq水中的10mmna2hpo4,ph11.4中,在30℃下注射到脂质体溶液中并搅拌30分钟,然后通过超滤进行浓缩步骤,并通过渗滤在pbsph7.4中稀释。然后通过使得到的脂质体通过0.2μm聚醚砜(pes)膜注射器过滤器对其进行无菌过滤,并在5℃下储存。
aci-24.043疫苗的制备(横流注射)
将脂质(dmpg、dmpc、胆固醇和3d-(6-酰基)phadtm(avantipolarlipids,usa))在60℃的加热箱中溶解于96%etoh中。在脂质完全溶解之后,将溶液通过0.2μm孔径的过滤器过滤到加热至60℃的注射系统中。详细地,在室温下借助声处理将适量的aci-24.043(sat47)分散在etoh中(最终sat47溶液中etoh浓度通常为2%v/v)。在肽完全分散之后,添加his-蔗糖缓冲液(10mm组氨酸、250mm蔗糖)以达到药物与脂质的比例为按质量计1/50。将sat47溶液通过0.2μm孔径的过滤器(sartoscale过滤器)过滤到注射缓冲液瓶中,然后将其加热至40℃。当脂质/etoh溶液与注射缓冲液混合时,在注射部位形成脂质体。在脂质体形成之后立即进行利用10mm组氨酸、250mm蔗糖的在线稀释步骤,以降低etoh浓度。将中间脂质体在rt下通过100nm孔径的聚碳酸酯膜挤出(1次通过)。使用中空纤维膜(mwco:500kd)进行超滤/渗滤(udf)以除去etoh,并将缓冲液交换为ph6.9的pbs(10体积交换)。然后使用分散缓冲液(pbsph6.9)将sat47脂质体稀释至总脂质浓度为1mg/ml并温热至35℃。将pal1-15在60℃下溶解在10mmna2hpo4ph11.4缓冲液中的10%w/v的β-og溶液中,并用相同的缓冲液进一步稀释至终浓度为1mg/ml。将ph调节至11.4。在使用横流注射模块混合这两种溶液之后,将脂质体悬液在35℃下在搅拌下进一步孵育30分钟以允许pal1-15的完全插入。进行使用中空纤维膜(mwco:500kd)的第二udf步骤以除去β-og并将缓冲液交换为10mm组氨酸、250mm蔗糖(10体积交换)。将产物浓缩至其最终体积,并通过0.2μmacrodiscmpes注射器过滤器进行过滤。
aci-24.046疫苗的制备(横流注射)
将脂质(dmpg、dmpc、胆固醇和3d-(6-酰基)phadtm(avantipolarlipids,usa))在60℃的加热箱中溶解于96%etoh中。在脂质完全溶解之后,将溶液通过0.2μm孔径的过滤器过滤到加热至60℃的注射系统中。平行地,将aci-24.046(sat44)在40℃下溶解于注射缓冲液(10mm组氨酸、250mm蔗糖)中。在sat44完全溶解之后,将溶液通过0.2μm孔径的过滤器(sartoscale)过滤到加热至40℃的注射缓冲液瓶中。当脂质/etoh溶液与注射缓冲液混合时,在注射部位形成脂质体。在脂质体形成之后立即进行利用10mm组氨酸、250mm蔗糖的在线稀释步骤,以降低etoh浓度。将中间脂质体在室温下通过100nm孔径的聚碳酸酯膜挤出(1次通过)。使用中空纤维膜(mwco:500kd)进行超滤/渗滤(udf)以除去etoh,并将缓冲液交换为ph6.9的pbs(10体积交换)。然后使用分散缓冲液(pbsph6.9)将sat44脂质体稀释至总脂质浓度为1mg/ml并温热至35℃。将pal1-15在60℃下溶解在10mmna2hpo4ph11.4缓冲液中的10%w/v的β-og溶液中,并用相同的缓冲液进一步稀释至终浓度为1mg/ml。检查ph并将其小心地调回至11.4。在使用注射模块混合这两种溶液之后,将脂质体悬液在35℃下在搅拌下进一步孵育30分钟以允许pal1-15的完全插入。进行使用中空纤维膜(mwco:500kd)的第二udf步骤以除去β-og并将缓冲液交换为10mm组氨酸、250mm蔗糖(10体积交换)。将产物浓缩至其最终体积,并通过0.2μmacrodiscmpes注射器过滤器进行最终过滤。
实施例3.具有包封的t细胞表位的疫苗的概念验证(proof-of-concept,poc)体内免疫原性研究
在成功包封不同t细胞表位之后,与aci-24疫苗相比,在体内测试了含有具有高免疫原性评分的包封的t细胞表位sat42、sat44和sat47的疫苗(分别为aci-24.044、aci-24.046和aci-24.043疫苗)的免疫原性。野生型c57bl/6小鼠在第0天、第14天和第28天接受aci-24、aci-24.044(具有包封的sat42)、aci-24.046(具有包封的sat44)和aci-24.043(具有包封的sat47)的总共3次皮下(s.c.)免疫接种。在第-21天(aci-24.046)或第-7天(aci-24、aci-24.043、aci-24.044)(预取血)、第7天、第21天和第35天收集血液样品以通过elisa测量aβ1-42特异性igg效价。
将板用10μg/ml人aβ1-42肽膜(bachem,switzerland)在4℃下包被过夜。用0.05%tween20/pbs洗涤并用1%bsa/0.05%tween/pbs封闭之后,将系列稀释的血浆添加至板并在37℃下孵育2小时。洗涤之后,将板与碱性磷酸酶(alkalinephosphatase,ap)缀合的抗小鼠igg抗体(jacksonimmunoresearch,pa,usa)在37℃下孵育2小时。最后的洗涤之后,将板与ap底物(pnpp)一起孵育2.5小时,并使用elisa板读数器在405nm下读数。通过参照市售抗体(6e10,biolegend,uk,目录号803002)的系列稀释来表示结果。图1a示出了随时间的具有或不具有包封的t细胞表位的aci-24疫苗诱导的aβ1-42特异性igg效价。尽管aci-24疫苗在第1次免疫接种之后第7天显示出最高的aβ1-42特异性igg效价,但当在aci-24疫苗中包封t细胞表位时,在第2次和第3次免疫接种之后观察到抗体效价的提高。
图1b中的结果显示,与aci-24相比,用包含包封的t细胞表位的aci-24疫苗免疫接种诱导了aβ特异性抗体效价的提高,其对于用aci-24.043(具有包封的sat47)疫苗免疫接种的组达到统计学显著性。
在食蟹猴研究中测试具有包封的sat42、sat43、sat44或sat47的疫苗。每组四只猴每月一次接受3次用以下进行的s.c.疫苗接种(第1天、第29天和第57天):aci-24.044(包封的sat42--两组总共8只猴),aci-24.046疫苗(包封的sat44--两组总共8只猴),aci-24.045疫苗(包封的sat43--4只猴)或aci-24.043疫苗(包封的sat47--4只猴)。在第一次免疫接种之前(第1天)和每次免疫接种之后1周和3周(第8天、第22天、第36天、第50天、第64天和第78天)收集血液,以通过elisa测量aβ1-42特异性igg效价。
将板用10μg/ml人aβ1-42肽膜(bachem,switzerland)在4℃下包被过夜。用0.05%tween20/pbs洗涤并用1%bsa/0.05%tween20/pbs封闭之后,向板添加8份2倍系列稀释的血清并在37℃下孵育2小时。洗涤之后,将板用辣根过氧化物酶(hrp)缀合的抗猴igg抗体(kpl,目录号n°07411021)在37℃下孵育2小时。洗涤之后,将板与50μlabts/h2o2(2,2’-连氮基-双(3-乙基苯并噻唑啉-6-磺酸)(hrp底物)一起孵育,并在1小时之后使用elisa板读数器在405nm处读数。通过参照用作标准的阳性猴合并物(pool)的系列稀释来表示结果。
将具有不同t细胞表位的疫苗的免疫原性与aci-24疫苗进行比较。表3显示第三次免疫接种之后1周,与aci-24疫苗相比的aβ特异性抗体效价倍数提高。与aci-24疫苗诱导的效价相比,所有受试疫苗aci-24.046(sat44)、aci-24.043(sat47)、aci-24.045(sat43)和aci-24.044(sat42)诱导抗体效价提高至少7倍(aci-24.044具有包封的sat42)。在第三次免疫接种之后1周,与aci-24相比,aci-24.043疫苗(具有包封的sat47)和aci-24.046(具有包封的sat44)诱导显著更高的aβ特异性抗体效价(表3)。aci-24.043疫苗(具有包封的sat47)和aci-24.046(具有包封的sat44)各自具有高epivax评分(分别为142.89和57.2)。
表3.与aci-24相比的aβ特异性抗体效价倍数提高(第三次免疫接种之后1周,第64天)
统计学检验:kruskal-wallis检验和dunn多重比较。*p<0.05;**p<0.01;***p<0.001;ns:非显著性。
在用根据薄脂膜技术制备的疫苗aci-24.046(包封的sat44)和aci-24.043(包封的sat47)在体内获得的结果(图1)之后,我们测试了根据横流注射方法制备的相同疫苗的体内免疫原性。野生型c57bl/6小鼠在第0天、第14天和第28天接受aci-24、aci-24.046(具有包封的sat44)和aci-24.043(具有包封的sat47)疫苗的总共三次皮下(s.c.)免疫接种。在第-7天、第7天、第21天和第35天收集血液样品以通过elisa测量aβ1-42特异性igg效价。
图6中的结果显示,与aci-24相比,用包含包封的t细胞表位的aci-24疫苗免疫接种诱导aβ特异性抗体效价的显著提高。
实施例4.诱导的aβ特异性抗体的品质
4.1人aβ1-42自缔合的体外抑制
通过测量aβ1-42自缔合/聚集的抑制体外测试诱导的aβ特异性抗体的品质。该测定基于小鼠免疫接种之前和免疫接种之后血浆损害人aβl-42对自缔合之天然易感性的能力。
将标准elisa板用1μg/mlaβ1-42在4℃下包被过夜。将板用300μl0.05%tween20/pbs洗涤4次。通过添加0.5%bsa/pbs并在37℃下孵育1小时实现饱和。洗涤之后,将四份2倍系列稀释的血浆在室温下在搅拌下添加至板,持续20分钟。向每个孔添加生物素化的aβ1-42至终浓度为0.1μg/ml,并在室温下搅拌孵育2小时。将没有血浆的生物素化的aβ1-42用作aβ1-42自缔合的阳性对照(被认为是100%的自缔合,0%的抑制)。在洗涤步骤之后,将板用辣根过氧化物酶(hrp)缀合的链霉亲和素(r&dsystems,canada,ref.890803)以1/200稀释度在0.5%bsa/0.05%tween20/pbs中在室温下搅拌孵育1小时。洗涤之后,将板与surebluereservetmb底物(seracare,目录号5120-0081)一起孵育10分钟。用bethyl终止溶液(bethyllaboratories,inc,目录号e115)终止反应,并使用elisa板读数器在450nm下对板进行读数。使用不含血浆的生物素化的aβ1-42作为阳性对照(0%抑制)作为参照计算自缔合的抑制百分比。
结果显示,用含有t细胞表位的所有疫苗进行2次免疫接种之后产生的aβ特异性抗体比aci-24诱导的抗体更高效地损害aβ1-42自缔合(图2a)。由于预取血血浆诱导自缔合的背景抑制,因此将第21天的百分比通过减去预取血血浆的背景进行标准化。与aci-24相比,用含有t细胞表位的所有aci-24疫苗免疫接种产生的aβ1-42特异性抗体显示出更高的aβ1-42自缔合抑制;在用aci-24.046(具有包封的sat44)免疫接种的组中,该抑制达到统计学显著性(图2b)。
4.2识别aβ寡聚体的抗体的产生
为了评价c57bl/6小鼠中诱导的抗体结合病理性aβ的特异性,通过elisa测定aβ1-42-寡聚体特异性igg应答。将板用10μg/ml如先前所述(adolfsson,2012)制备的寡聚体在4℃下包被过夜。用0.05%tween20/pbs洗涤并用1%bsa/0.05%tween20/pbs封闭之后,将系列稀释的血浆添加至板并在37℃下孵育2小时。洗涤之后,将板与碱性磷酸酶(ap)缀合的抗小鼠igg抗体(jacksonimmunoresearch,目录号115-055-164,pa,usa)在37℃下孵育2小时。最后的洗涤之后,将板与ap底物(pnpp)一起孵育2.5小时,并使用elisa板读数器在405nm处读数。通过参照市售抗体(6e10,biolegend,uk,目录号803002)的系列稀释来表示结果。
基于aβ1-42抗体效价,在以1/100、1/400、1/800或1/1600稀释度开始的8份或4份2倍系列稀释中测试每个样品。图3中的结果显示,与aci-24相比,用含有t细胞表位的所有aci-24疫苗免疫接种诱导了aβ1-42寡聚体特异性抗体效价的提高,其对于用aci-24.043(具有包封的sat47)疫苗免疫接种的组达到统计学显著性。通过elisa测定确定免疫接种之后7和21天c57bl/6小鼠中诱导的抗体的亲合力指数。标准elisa板的一半用10μg/ml的aβ1-42肽膜且另一半用1μg/ml的aβ1-42肽膜在4℃下包被过夜。在用0.05%tween20/pbs洗涤并用1%bsa/0.05%tween20/pbs封闭之后,将8份2倍系列稀释的血浆添加至这两种包被条件,并在37℃下孵育2小时。洗涤步骤之后,将板与碱性磷酸酶(ap)缀合的抗小鼠igg抗体(jacksonimmunoresearch,目录号115-055-164,pa,usa)在37℃下孵育2小时。最后的洗涤之后,将板与ap底物(pnpp)一起孵育2.5小时,并使用elisa板读数器在405nm处读数。通过参照市售抗体(6e10,biolegend,uk,目录号803002)的系列稀释来表示结果。
为了确定亲合力指数,使用在10μg/ml的aβ1-42肽上获得的标准曲线计算这两种包被上每种样品的au/ml。0.6至2.8的o.d.值用于反向计算浓度。亲合力指数计算为较低包被浓度(1μg/mlaβ1-42肽)与饱和包被(10μg/mlaβ1-42肽)上的抗体浓度之间的比值。
图4中的结果显示用含有t细胞表位的所有aci-24疫苗免疫接种在第1次免疫接种与第2次免疫接种(分别为第7天和第21天)之间诱导aβ1-42特异性抗体亲合力成熟,这在用aci-24.044(具有包封的sat42)和aci-24.043(具有包封的sat47)免疫接种的组中达到统计学显著性。
为了评价食蟹猴中诱导的抗体结合病理性aβ的特异性,在用aci-24.046(具有包封的sat44--两组总共8只猴)、aci-24.045疫苗(包封的sat43--4只猴)或aci-24.043疫苗(包封的sat47--4只猴)免疫接种的食蟹猴的血清中,在第64天(第三次免疫接种之后1周)通过mesoscalediscovery(msd)技术测量aβ1-42寡聚体特异性igg效价。将msd链霉亲和素板在4℃下用5%的阻断剂a(msd,ref.r93ba-4)饱和过夜。第二天,将板用0.05%tween20/pbs洗涤4次,并用25μl的0.5μg/ml的在pbs中的捕获抗体生物素化的6e10(biolegend,ref.803008)在摇床(shaker)上在37℃下包被1小时。洗涤之后,将板与25μl的在pbs中的10μg/mlaβ1-42寡聚体(adolfsson,2012)在摇床上在37℃下孵育1小时。将板洗涤并与8份2倍稀释的猴血清(1%脱脂乳/0.05%tween/pbs中的起始稀释1/50)一起孵育。将样品在摇床上在37℃下孵育2小时。将板洗涤4次并添加用sulfo-tag标记的抗人igg检测抗体(jackson,ref.109-005-098),在1%脱脂乳/0.05%tween20/pbs中在摇床上在37℃下稀释1小时。洗涤4次之后,添加msd读数缓冲液t2x(msd,ref.r92tc-2),并在5分钟内对板进行读数。通过参照用作标准的猴合并物的系列稀释来表达结果。
结果显示,与第1天(第一次免疫接种之前)相比,所有受试疫苗aci-24.046(包封的sat44)、aci-24.043(包封的sat47)和aci-24.045(包封的sat43)在第64天(第三次免疫接种之后1周)诱导能够识别aβ寡聚体的抗体的提高;参见图5。
参考文献
adolfssono.,pihlgrenm.,tonin.,variscoy.,buccarelloa.l.,antoniellok.,lohmanns.,
piorkowskak.,gafnerv.,atwalj.k.,maloneyj.,chenm.,goginenia.,weimerr.m.,
mortensend.l.,friesenhahnm.,hoc.,paulr.,pfeifera.,muhsa.,wattsr.j.,aneffector-reducedanti-β-amyloid(aβ)antibodywithuniqueaβbindingpropertiespromotesneuroprotectionandglialengulfmentofaβ.jneurosci.jul11;32(28):9677-89(2012).
agadjanyanm.g.,ghochikyana.,petrushinal,vasilevkov.,movsesyann.,mkrtichyanm.,saingt.andcribbsd.h.,prototypealzheimer′sdiseasevaccineusingtheimmunodominantbcellepitopefromβ-amyloidandpromiscuoustcellepitopepanhladr-bindingpeptide.jimmunol174(3)1580-1586(2005).
araih,suzukih,yoshiyamat.vanutidecridificarandtheqs-21adjuvantinjapanesesubjectswithmildtomoderatealzheimer′sdisease:resultsfromtwophase2studies.curralzheimerres.12(3):242-54(2015).
ghochikyana.,mkrtichyanm.,petrushinal.,movsesyann.,karapetyana.,cribbsd.h.,agadjanyanm.g.,prototypealzheimer′sdiseaseepitopevaccineinducedstrongth2-typeanti-abetaantibodyresponsewithalumtoquilaadjuvantswitch.vaccine.20;24(13):2275-82(2006).
gilmans.,kollerm.,blackr.s.,jenkinsl.,griffiths,g.,foxn.c.,eisnerl.,kirbyl.,boadaroviram.,forettef.,orgogozoj.m.,clinicaleffectofaβimmunization(an1792)inpatientswithadinaninterruptedtrial,neurology64,1553-1562(2005).
liub,frostj.l.,sunj.,fuh.,grimess.,blackburnp.,lemerec.a.,mer5101,anovelaβ1-15:dtconjugatevaccine,generatesarobustanti-aβantibodyresponseandattenuatesaβpathologyandcognitivedeficitsinappswe/ps1δe9transgenicmice.jneurosci.33(16):7027-37(2013).
lutznern.,kalbacherh.,quantifyingcathepsinsactivityinantigenpresentingcellsusinganovelspecificsubstrate.j.biol.chem.vol.283no.52p.36185(2008).
maierm.,seabrookt.j.,lazon.d.,jiangl.,dasp.,janusc,lemerec.a,shortamyloid-beta(abeta)immunogensreducecerebralabetaloadandlearningdeficitsinanalzheimer′sdiseasemousemodelintheabsenceofanabeta-specificcellularimmuneresponse.jneurosci.3;26(18):4717-28(2006).
martineaup,chapter41:affinitymeasurementsbycompetitionelisa,pages657-665,frombook:antibodyengineering,vol.1;r.kontermannands.dübel(2010)
monsonegoa.,weinerh.l.,immunotherapeuticapproachestoalzheimer′sdisease.science.31;302(5646):834-8(2003).
muhsa.,hickmand.t.,pihlgrenm.,chuardn.,giriensv.,meerschmanc.,vanderauweral.,vanleuvenf.,sugawaram.,weingertnerm.-c.,bechingerb.,greferathr.,kolonkon.,nagel-stegerl.,riesnerd.,bradyr.o.,pfeifera.,nicolauc.,liposomalvaccineswithconformation-specificamyloidpeptideantigensdefineimmuneresponseandefficacyinapptransgenicmice.pnas,10423:9810-9815(2007).
orgogozoj.m.,gilmans.,dartiguesj.f.,laurentb.,puelm.,kirbyl.c.,jouannyp.,duboisb.,eisnerl.,flitmans.,michelb.f.,boadam.,franka.,hockc.,subacutemeningoencephalitisinasubsetofpatientswithadafterabet42immunization.neurology61:46-54(2003).
pihlgrenm.,silvaa.b.madanir,giriensv.,waeckerle-meny.,fettelschossa.,hickmand.t.,lópez-deberm.p.,ndaod.m.,vukicevicm.,buccarelloa.l.,gafnerv.,chuardn.,reisp.,piorkowskak.,pfeifera.,kündigt.m.,muhsa.,johansenp.,tlr4-andtrif-dependentstimulationofblyvmphocytesbypeptideliposomesenablestcell-independentisotypeswitchinmice,blood.jan3;121(1):85-94(2013).
sallustof.,lanzavecchiaa.,arakik.,ahmedr.,fromvaccinestomemoryandback.immunity.oct29;33(4):451-63(2010).
schneebergera.,mandlerm.,mattnerf.,schmidtw.,
seabrookt.j.,thomask.,jiangl.,bloomj.,spoonere.,maierm.,bitang.,lemerec.a.,dendrimericabeta1-15isaneffectiveimmunogeninwildtypeandapp-tgmice.neurobiolaging.28(6):813-23(2006).
siegristca,chapter2:vaccineimmunology,pages14-32frombook:vaccine(6thedition,2013).
sotoc.,plaquebusters:strategiestoinhibitamyloidformationinalzheimer′sdisease.molecularmedicinetoday(vol5),august1999.
winbladb.,grafa.,rivierem.e.,andreasenn.,ryanj.m.,activeimmunotherapyoptionsforalzheimer′sdisease.alzheimersresther.2014jan30;6(1):7.
winbladb.,andreasenn.,minthonl.,floessera.,imbertg.,dumortiert.,maguirer.p.,blennowk.,lundmarkj.,staufenbielm.,orgogozoj.m.,grafa.,safety,tolerability,andantibodyresponseofactiveaβimmunotherapywithcad106inpatientswithalzheimer′sdisease:randomised,double-blind,placebo-controlled,first-in-humanstudy.lancetneurol.11(7):597-604(2012).
除非另外定义,否则本文中使用的所有技术和科学术语具有与本发明所属领域的普通技术人员通常理解的相同的含义。出于与本发明有关的所有目的,本文中具体提及的所有出版物和专利均通过引用整体并入。
本发明不限于本文中所述的具体实施方案的范围。实际上,通过前面的描述和附图,除了本文中所述的那些之外的本发明的多种修改对于本领域技术人员来说将变得明显。这样的修改旨在落入所附权利要求书的范围内。此外,本文中描述的本发明的所有方面和实施方案被认为是广泛适用的并且可与任何和所有其他一致的实施方案组合,包括适当地从本发明的其他方面获得的那些(包括单独的(inisolation))。
序列表
<110>acimmunesa
<120>抗aβ治疗性疫苗
<130>p158775wo00
<150>ep18166659
<151>2018-04-10
<150>ep18202366.3
<151>2018-10-24
<160>8
<170>patentinversion3.5
<210>1
<211>40
<212>prt
<213>人工序列
<220>
<223>合成肽
<400>1
valhishisasnthrglugluilevalalaglnserilealaleuser
151015
serleumetvalprometglyalaproglntyrilelysalaasnser
202530
lyspheileglyilethrgluleu
3540
<210>2
<211>57
<212>prt
<213>人工序列
<220>
<223>合成肽
<400>2
valtyrglyglyserlysthrserleutyrasnleuargargglythr
151015
alaleualailevalvalargglntyrilelysalaasnserlysphe
202530
ileglyilethrgluleuvalvalargproilephepheleuhishis
354045
serasnthraspargleutrpalaile
5055
<210>3
<211>38
<212>prt
<213>人工序列
<220>
<223>合成肽
<400>3
valtyrglyglyserlysthrserleutyrasnleuargargglythr
151015
alaleualailevalvalargglntyrilelysalaasnserlysphe
202530
ileglyilethrgluleu
35
<210>4
<211>52
<212>prt
<213>人工序列
<220>
<223>合成肽
<400>4
sermetglyvaltyrglnileleualailetyrserthrvalvalarg
151015
ilevalalaglnserilealaleuserservalvalargtyrilelys
202530
alaasnserlyspheileglyvalvalargleutyrasnleuargarg
354045
glythralaleu
50
<210>5
<211>50
<212>prt
<213>大锁孔帽贝(megathuracrenulata)
<400>5
serthrleuglutyrpheleutyraspproilephepheleuhishis
151015
serasnthraspargleutrpalailetrpglnalaleuglnlystyr
202530
argglylysprotyrasnthralaasncysalailevalarghisasp
354045
thrtyr
50
<210>6
<211>20
<212>prt
<213>白喉棒状杆菌(corynebacteriumdiphtheriae)
<400>6
valhishisasnthrglugluilevalalaglnserilealaleuser
151015
serleumetval
20
<210>7
<211>25
<212>prt
<213>流感病毒(influenzavirus)
<400>7
ileaspglyvallysleuglusermetglyvaltyrglnileleuala
151015
iletyrserthrvalalaserserleu
2025
<210>8
<211>20
<212>prt
<213>人疱疹病毒4(humanherpesvirus4)
<400>8
valtyrglyglyserlysthrserleutyrasnleuargargglythr
151015
alaleualaile
20
起点商标作为专业知识产权交易平台,可以帮助大家解决很多问题,如果大家想要了解更多知产交易信息请点击 【在线咨询】或添加微信 【19522093243】 与客服一对一沟通,为大家解决相关问题。
与客服一对一沟通,为大家解决相关问题。
此文章来源于网络,如有侵权,请联系删除

 热门咨询
热门咨询



