
 商标分类
商标分类  商标转让
商标转让 
一种水稻籽粒粒长和粒重相关蛋白及其编码基因与应用的制作方法
 2021-02-02 18:02:19|
2021-02-02 18:02:19| 425|
425| 起点商标网
起点商标网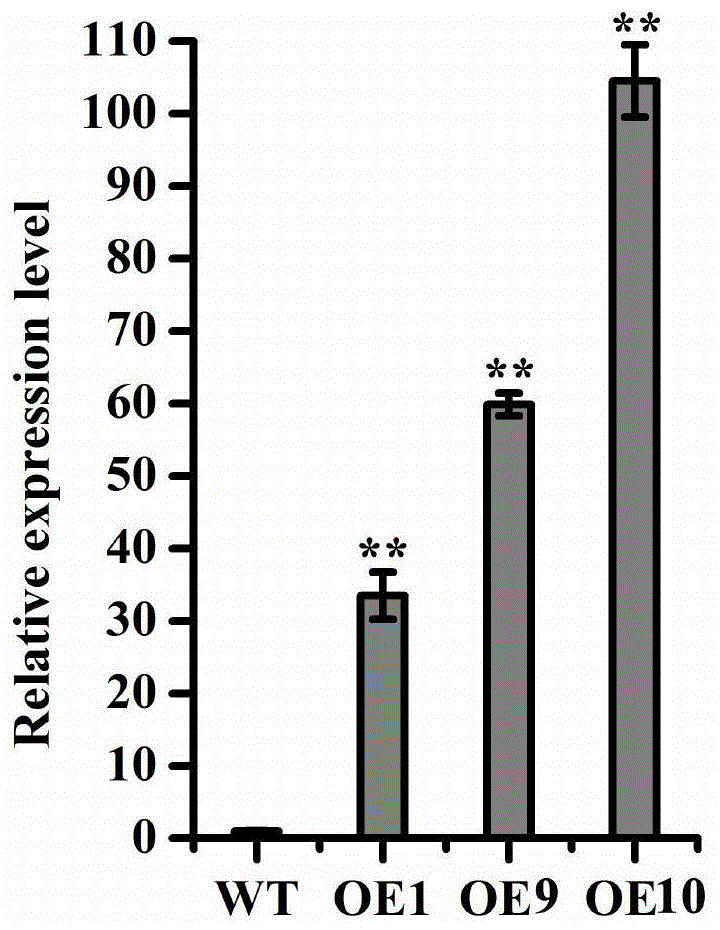
本发明涉及植物基因工程技术领域,具体涉及一种水稻籽粒粒长和粒重相关蛋白及其编码基因与应用。
背景技术:
水稻是世界上最重要的粮食作物之一,世界一半以上的人口以水稻为主食。近年来,随着耕地面积减少、能源短缺、气候变化及世界人口增加等问题的日益严峻,粮食安全问题愈来愈严重。因此,提高水稻的产量,改善稻米品质已成为我国植物育种的重要研究目标。水稻产量的三大要素:单株穗数、每穗粒数、粒重。水稻的粒型性状包括粒长、粒宽、粒厚及粒长宽比,与粒重呈正相关,是粒重的决定因子。
目前,研究报道关于参与调控水稻粒型或粒重的基因和qtl已有近60个基因。其中与水稻籽粒大小变异关系较为密切的基因包括粒宽基因gw2、gs5、gw8、gw5/qsw5、gw7;粒长基因gl3.1、gs3、gs2、glw7等。
gw2编码一个ring型的e3泛素连接酶,位于细胞质中,调控籽粒的宽度。gw2通过将其底物锚定到蛋白酶体进行降解,从而负调控细胞的分裂。该基因的功能缺失突变使得本应降解的底物不能被特异识别,从而激活颖壳的细胞分裂,使得颖壳细胞数目增加,导致其颖壳宽度增加。此外,其籽粒的灌浆速率和胚乳提高,最终粒宽、粒重以及产量都得到了提高。gs5编码一个色氨酸羧肽酶,该基因的高表达促进细胞分裂,增加细胞数目,并且细胞周期相关基因的表达上调,是水稻籽粒大小的正调控因子。最新研究报道gs5对osbak1-7和osmsbp1之间的互作具有竞争抑制作用,表明gs5可能影响br信号。gw8编码一个包含sbp结构域的转录因子osspl16,调控水稻粒宽。研究表明gw8能够调节细胞周期,正向调控细胞分裂相关基因的表达,增加细胞增殖。该基因的高表达能促进细胞分裂和籽粒灌浆,从而增加水稻粒宽和产量。gw5/qsw5编码一个核定位蛋白,其能与多聚泛素互作。gw5可能通过泛素蛋白酶体途径来调控水稻粒宽和粒重。gw5蛋白位于细胞质膜上,能与水稻gsk2(拟南芥bin2激酶的同源蛋白)互作,并抑制水稻gsk2激酶的活性,导致在下游bzr1和dlt转录因子的非磷酸化蛋白积累增加,从而调控油菜素内脂的下游基因表达,引起籽型的改变。gw7编码一个tonneau招募蛋白质,与c端人类中心体蛋白质同源。过表达gw7基因能够产生细长的籽粒,其可能是由于促进了籽粒纵向细胞分裂而抑制横向细胞分裂所造成的结果。gl3.1/qgl3编码一个具有kelch重复结构域的蛋白磷酸酶(osppkl1),通过去磷酸化细胞周期蛋白cyclin-t1;3抑制颖壳的细胞分裂,从而负调控籽粒的长度。gl3包含有两个kelch结构域,其第二个kelch结构域上保守的avldt区域的第364位天冬氨酸替换成谷氨酸的稀有等位变异会导致籽粒变长。最新的研究表明,qgl3是拟南芥bsu1基因的同源蛋白。在拟南芥中bsu1正调控br信号,而水稻中qgl3的过表达产生br功能缺失突变的表型。qgl3能够与水稻osgsk3激酶(拟南芥bin2基因的同源蛋白)互作,且去磷酸化osgsk3并稳定其状态,表明qgl3通过调控osgsk3的磷酸化和稳定性而抑制br信号。gs3编码一个由232个氨基酸组成的跨膜蛋白,通过控制颖壳的细胞数目来负调控籽粒长度。该蛋白包含4个结构域:一个调控器官大小的结构域osr、一个跨膜结构域m、一个肿瘤坏死因子受体家族中富含半胱氨酸同源区域t和一个c型血管性血友病因子vwfc。其中osr结构域是gs3实现其功能的一个关键结构域,osr结构域缺失会产生长粒表型。gs2编码一个定位在核内的转录激活因子osgrf4,过表达该基因能够促进细胞分裂和扩张,从而显著提高水稻的粒重和产量。gl2/gs2能与br信号转导通路中的负调控因子gsk2互作,其转录激活活性受gsk2抑制,其表达受osmir396调节。gs2还能与转录共激活因子osgif1/2/3互作,osgif1的过表达增加籽粒的大小和产量。glw7编码一个在植物中特有的转录因子osspl13。glw7的高表达通过促进细胞伸长,进而增加籽粒的长度和产量,而其功能缺失突变体产生短粒。此外,glw7可以结合srs5和dep1的启动子,促进其表达,但其具体的遗传互作关系仍然未知。
尽管已经有许多调控种子粒型的基因被克隆,但它们的遗传调控通路网络还不太清楚,其之间的相互关系还有待进一步深入研究。因此,克隆水稻的粒型基因,对解析水稻粒型相关基因的遗传调控机制具有重要的意义。
技术实现要素:
本发明的目的是提供一种水稻籽粒粒长和粒重相关蛋白及其编码基因与应用。
第一方面,本发明保护osmob1a蛋白在调控植物籽粒长度和/或籽粒重中的应用;
所述osmob1a蛋白为如下任一所示蛋白质:
(a1)由序列表中序列1所示的氨基酸序列组成的蛋白质;
(a2)序列表的序列1所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且有相同功能的由序列1衍生的蛋白质;
(a3)与(a1)-(a2)中任一所限定的氨基酸序列具有99%以上、95%以上、90%以上、85%以上或者80%以上同源性且具有相同功能的蛋白质;
(a4)在(a1)-(a3)中任一所限定的蛋白质的n端和/或c端连接标签后得到的融合蛋白。
所述osmob1a蛋白来源于水稻。
上述蛋白质可人工合成,也可先合成其编码基因,再进行生物表达得到。
上述蛋白质中,所述标签可为flag标签、his标签、mbp标签、ha标签、myc标签、gst标签和/或sumo标签等。
第二方面,本发明保护osmob1a蛋白相关生物材料在调控植物籽粒长度和/或籽粒重中的应用;所述osmob1a蛋白为前文(a1)-(a4)中任一所示蛋白质。
所述相关生物材料为能够表达所述osmob1a蛋白的核酸分子或含有所述核酸分子的表达盒、重组载体、重组菌或转基因细胞系。
上述应用的具体体现如下:
所述osmob1a蛋白或其编码基因在所述植物中的活性和/或表达量提高,植物籽粒长度增加;
所述osmob1a蛋白或其编码基因在所述植物中的活性和/或表达量提高,植物籽粒重增加。
第三方面,本发明保护培育籽粒长度提高和/或籽粒重提高的植物品种的方法,包括使受体植物中osmob1a蛋白的表达量和/或活性提高的步骤;所述osmob1a蛋白为前文(a1)-(a4)中任一所示蛋白质。
进一步地,本发明保护培育转基因植物的方法,包括如下步骤:向受体植物中导入能够表达osmob1a蛋白的核酸分子,得到转基因植物;所述转基因植物籽粒长度大于受体植物和/或籽粒重大于受体植物;所述osmob1a蛋白为前文(a1)-(a4)中任一所示蛋白质。
所述“向受体植物中导入能够表达osmob1a蛋白的核酸分子”是通过向所述受体植物中导入含有所述osmob1a蛋白的编码基因的重组表达载体实现的。
可用现有的表达载体构建含有所述osmob1a蛋白的编码基因的重组表达载体。所述植物表达载体包括双元农杆菌载体和可用于植物微弹轰击的载体等。所述植物表达载体还可包含外源基因的3′端非翻译区域,即包含聚腺苷酸信号和任何其它参与mrna加工或基因表达的dna片段。所述聚腺苷酸信号可引导聚腺苷酸加入到mrna前体的3′端,如农杆菌冠瘿瘤诱导(ti)质粒基因(如胭脂碱合成酶基因nos)、植物基因(如大豆贮存蛋白基因)3′端转录的非翻译区均具有类似功能。
使用osmob1a蛋白的编码基因构建重组植物表达载体时,在其转录起始核苷酸前可加上任何一种增强型启动子或组成型启动子(如花椰菜花叶病毒(camv)35s启动子、玉米的泛素启动子(ubiquitin)),或组织特异表达启动子(如种子特异表达的启动子),它们可单独使用或与其它植物启动子结合使用。此外,使用osmob1a蛋白的编码基因构建植物表达载体时,还可使用增强子,包括翻译增强子或转录增强子,这些增强子区域可以是atg起始密码子或邻接区域起始密码子等,但必需与编码序列的阅读框相同,以保证整个序列的正确翻译。所述翻译控制信号和起始密码子的来源是广泛的,可以是天然的,也可以是合成的。翻译起始区域可以来自转录起始区域或结构基因。
为了便于对转基因植物细胞或植物进行鉴定及筛选,可对所用植物表达载体进行加工,如加入可在植物中表达的编码可产生颜色变化的酶或发光化合物的基因(gus基因、萤光素酶基因等)、具有抗性的抗生素标记物(庆大霉素标记物、卡那霉素标记物等)或是抗化学试剂标记基因(如抗除莠剂基因)等。
在本发明中,所述重组表达载体具体可为将pun1301载体的bamhi和kpni酶切位点间的片段替换为序列表的序列2所示的dna分子得到的重组表达载体。
在上述方法中,将携带有所述osmob1a蛋白的编码基因的所述重组表达载体导入所述受体植物,具体可为:通过使用ti质粒、ri质粒、植物病毒载体、直接dna转化、显微注射、电导、农杆菌介导等常规生物学方法转化植物细胞或组织,并将转化的植物组织培育成植株。
转化的细胞、组织或植物理解为不仅包含转化过程的最终产物,也包含其转基因子代。
所述“osmob1a蛋白的编码基因”是如下任一所述的dna分子:
(b1)序列表的序列2所示的dna分子;
(b2)在严格条件下与(b1)限定的dna分子杂交且编码所述osmob1a蛋白的dna分子;
(b3)与(b1)或(b2)限定的dna序列具有99%以上、95%以上、90%以上、85%以上或者80%以上同一性且编码所述osmob1a蛋白的dna分子。
上述基因中,所述严格条件可为如下:50℃,在7%十二烷基硫酸钠(sds)、0.5mnapo4和1mmedta的混合溶液中杂交,在50℃,2×ssc,0.1%sds中漂洗;还可为:50℃,在7%sds、0.5mnapo4和1mmedta的混合溶液中杂交,在50℃,1×ssc,0.1%sds中漂洗;还可为:50℃,在7%sds、0.5mnapo4和1mmedta的混合溶液中杂交,在50℃,0.5×ssc,0.1%sds中漂洗;还可为:50℃,在7%sds、0.5mnapo4和1mmedta的混合溶液中杂交,在50℃,0.1×ssc,0.1%sds中漂洗;还可为:50℃,在7%sds、0.5mnapo4和1mmedta的混合溶液中杂交,在65℃,0.1×ssc,0.1%sds中漂洗;也可为:在6×ssc,0.5%sds的溶液中,在65℃下杂交,然后用2×ssc,0.1%sds和1×ssc,0.1%sds各洗膜一次。
第四方面,本发明保护osmob1a蛋白或其相关生物材料,或,以上任一所述的方法在植物育种中的应用;
所述相关生物材料为能够表达所述osmob1a蛋白的核酸分子或含有所述核酸分子的表达盒、重组载体、重组菌或转基因细胞系;所述osmob1a蛋白为前文(a1)-(a4)中任一所示蛋白质。
所述育种的目的是选育籽粒长度大和/或籽粒重大的植物。
在上述各方面中,所述植物为(c1)或(c2)或(c3):
(c1)双子叶植物或单子叶植物;
(c2)禾本科植物;
(c3)水稻。
所述水稻具体可为中花11号水稻。
本发明的实验证明,利用重组载体将osmob1a基因导入野生型水稻中,得到转基因水稻,其与野生型水稻相比,osmob1a基因过量表达,水稻的粒长显著增加,粒重显著增加。因此osmob1a基因与粒型相关,为提高粒型改变的转基因植物培育奠定基础。本发明对于进一步阐明植物籽粒发育分子机理并通过基因工程手段培育优质、高产的作物新品种具有重要的理论意义和现实意义。
附图说明
图1为转基因水稻的real-timepcr表达鉴定。
图2为osmob1a超表达转基因水稻的表型观察。
图3为osmob1a超表达转基因水稻的粒长、千粒重的统计数据。
具体实施方式
以下的实施例便于更好地理解本发明,但并不限定本发明。下述实施例中的实验方法,如无特殊说明,均为常规方法。下述实施例中所用的试验材料,如无特殊说明,均为自常规生化试剂商店购买得到的。以下实施例中的定量试验,均设置三次重复实验,结果取平均值。
pun1301载体:记载于文献“zhangc,xuy,guos,zhuj,huanq,liuh,wangl,luog,wangx,chongk.dynamicsofbrassinosteroidresponsemodulatedbynegativeregulatorlicinrice.plosgenetics.2012,8:e1002686”,公众可从中国科学院植物研究所获得。
中花11号水稻:记载于文献“倪丕冲,水稻花培新品种—中花11号,农业科技通讯,1989(04):42”,公众可从中国科学院植物研究所获得。
实施例1、osmob1a蛋白及其编码基因的获得
提取中花11号水稻幼苗的总rna,并反转录为cdna。经过大量序列分析、表达量分析与功能验证,从cdna中发现了一个dna编码序列,如序列表的序列2所示,其编码的蛋白质如序列表的序列1所示。
将序列表的序列1所示的蛋白质命名为osmob1a蛋白。将编码osmob1a蛋白的基因命名为osmob1a基因,其开放阅读框如序列表的序列2所示。
实施例2、重组表达载体构建和重组农杆菌的获得
一、重组表达载体的构建
将pun1301载体的bamhi和kpni酶切位点间的片段替换为序列表的序列2所示的dna分子,得到重组表达载体pun1301-osmob1a(已经测序验证)。
二、重组农杆菌的获得
1、将重组质粒pun1301-osmob1a导入农杆菌eh105,得到重组农杆菌,将其命名为重组农杆菌eh105/pun1301-osmob1a。
2、将pun1301载体导入农杆菌eh105,得到重组农杆菌,将其命名为重组农杆菌eh105/pun1301。
实施例3、osmob1a蛋白在水稻中的功能验证
一、转osmob1a基因水稻植株的获得
通过农杆菌介导的水稻遗传转化体系将实施例2制备的重组农杆菌eh105/pun1301-osmob1a转入中花11号水稻(oryzasatival.cvzhonghua11,以下简称野生型水稻)胚的愈伤组织中,经过诱导、预培养、侵染、共培养、筛选具有潮霉素抗性的愈伤、分化、生根、移苗得到转基因植株。转化分别获得10个株系的t0代转基因osmob1a水稻植株。具体步骤如下:
1)水稻成熟胚胚性愈伤组织的诱导和继代:挑选籽粒饱满的去除颖壳后的野生型中花11号水稻种子,加入70%乙醇进行表面消毒5min。然后用20%次氯酸钠(naclo)消毒20min,消毒完成后用灭菌双蒸水溶液冲洗4-5次,无菌滤纸吸干后转移至诱导培养基nb2上,切记让切面跟培养基接触且不可将胚按压进培养基中,轻轻放置在培养基表面即可。25℃条件下暗培养4-7天后,将从成熟胚盾片处诱导出初生愈伤组织,同时需要在超净台中用镊子夹断长出的新芽。25℃条件下暗培养4-5周。为了得到较好的愈伤,培养两周后更换一次培养基,以保证充足的营养供应。选择浅黄色质地较致密的胚性愈伤组织,转接于继代培养基nb1上,25℃条件下暗培养2-3周;之后再挑选浅黄色质地致密的愈伤块,转移到新的nb1继代培养基上,25℃条件下继续暗培养2-3周。继代培养2-4次后,选择生长良好的黄色致密愈伤组织,转移到新的nb1培养基上,25℃条件下继续暗培养4-6天即可用于农杆菌侵染转化。
2)农杆菌菌株活化:将重组农杆菌eh105/pun1301-osmob1a在含50mg/l卡那霉素的yep固体培养基上划线,于28℃倒置培养2-3天。挑取重组农杆菌eh105/pun1301-osmob1a单克隆菌落接种到5ml含50mg/l卡那霉素的yep液体培养基中,28℃,200rpm振荡培养至对数生长晚期,吸取0.5ml菌液转移到50ml含有50mg/l卡那霉素的yep液体培养基中,28℃,200rpm继续震荡培养至od600为0.5左右。将上述菌液转入已灭菌的50ml离心管,室温,4,000rpm离心10min,弃上清,用等体积的液体培养基aam-as重悬菌体,准备侵染愈伤。
3)愈伤组织与农杆菌的共培养及转化:将预培养4-6天的愈伤组织块加入aam-as重悬的菌液浸染20min。浸染后的愈伤块用无菌吸水纸吸干后,转移到铺有一层滤纸的nb2c共培养培养基上,25℃条件下暗培养2-4天。
4)愈伤组织的筛选培养:将共培养好的愈伤组织转移至筛选培养基nbs1上,25℃条件下暗培养2周。筛选过程中大部分愈伤块都会变成褐色,含转基因的愈伤块变成褐色后会在愈伤边缘再次长出黄白色的愈伤组织,即抗性愈伤。挑取抗性愈伤块转移至筛选培养基nbs2上,25℃暗培养,继续筛选2周。
5)愈伤的诱导分化及成苗:将黄色的抗性愈伤组织转移至预分化培养基re1上,暗培养1周后,光照培养2-3周。将分化产生的不定芽转移至分化培养基re2上,光照培养2-3周即可分化出植株。分化产生的植株长至2-3cm时可以转移至含生根培养基的1/2ms培养基中,光照培养2周左右。待小苗长至10cm左右时,打开封口膜,加入少量蒸馏水。光照下炼苗5-7天后,可将组织培养苗移栽至土中生长。
上述步骤中所用培养基见表1。
表1培养基配方
二、转空载体水稻植株的获得
采用重组农杆菌eh105/pun1301替代重组农杆菌eh105/pun1301-osmob1a,按照步骤一进行操作,得到转空载体水稻植株。
三、转osmob1a基因水稻植株的鉴定
取生长3-4周的t0代转基因osmob1a水稻植株叶片,提取基因组dna,以基因组dna为模板,采用引物1301-f和引物osmob1a-r组成的引物对进行pcr扩增,如果pcr扩增出大小约为696bp的条带,则表明植株为阳性转基因植株,共鉴定出10个株系的t0代转osmob1a水稻植株。将阳性转osmob1a水稻移植至温室栽培,分别单株收种,得到t1代转基因种子,之后繁殖得到纯合t2代种子,在之后的实验中选取编号为1(oe1)、9(oe9)、10(oe10)的t2代转osmob1a水稻为材料。
1301-f:5’-gctacgagtgggacaacaaa-3’;
osmob1a-r:5’-gtatggaataatcgattcgatg-3’。
三、转osmob1a基因水稻植株的mrna表达量检测
待测植株:水稻野生型中花11号、编号为1(oe1)、9(oe9)、10(oe10)的t2代转osmob1a水稻植株和t2代转空载体植株。
1、提取待测植株三叶期的幼苗叶片总rna,并反转录为cdna。
2、以步骤1得到的cdna为模板,进行实时定量rt-pcr检测。采用引物osmob1a-qrt-f和引物osmob1a-qrt-r组成的引物对检测osmob1a基因的表达水平,采用引物osactin-qrt-f和osactin-qrt-r组成的引物对检测内参基因osactin。
osmob1a-qrt-f:5’-gatgccactcttggcagtgg-3’;
osmob1a-qrt-r:5’-gttggccttcaatccagtcc-3’。
osactin-qrt-f:5’-aggccaatcgtgagaagatgac-3’;
osactin-qrt-r:5’-aggtcgagacgaaggatagcat-3’。
每种材料每个株系进行了3次生物学重复实验检测。
结果如图1所示。结果显示,与野生型水稻(wt)相比,t2代转osmob1a水稻株系oe1、oe9和oe10中的osmob1a基因表达量显著上调,说明osmob1a基因在转基因水稻中成功大量表达。t2代转空载体植株与野生型水稻osmob1a基因表达水平无显著差异。
四、转osmob1a基因水稻植株的表型观察
待测植株:水稻野生型中花11号、编号为1(oe1)、9(oe9)、10(oe10)的t2代转osmob1a水稻植株和t2代转空载体植株。
将待测植株种植在中国科学院植物研究所试验田中,待水稻抽穗灌浆后,对水稻的粒型进行性状考察和拍照。
每种材料每个株系统计了20棵。
结果如图2所示。结果显示,与野生型水稻相比,t2代转osmob1a水稻的籽粒长度显著增加。野生型水稻(wt)和t2代转空载体水稻结果无显著差别。
五、转osmob1a基因水稻植株的农艺学性状统计
待测植株:水稻野生型中花11号、编号为1(oe1)、9(oe9)、10(oe10)的t2代转osmob1a水稻植株和t2代转空载体植株。
收集待测植株成熟籽粒,收获烘干后,统计20株平均千粒重和种子的平均长度。
结果如图3和表2所示。图3a为籽粒的长度,图3b为1000粒饱满水稻籽粒的千粒重。
表2野生型及转基因植株粒型相关性状的统计
结果显示,与野生型水稻相比,t2代转osmob1a水稻的籽粒长度、千粒重显著增加,表明osmob1a基因的超表达可以提高籽粒长度和千粒重。野生型水稻(wt)和t2代转空载体水稻结果无显著差别。
序列表
<110>中国科学院植物研究所
<120>一种水稻籽粒粒长和粒重相关蛋白及其编码基因与应用
<160>2
<170>siposequencelisting1.0
<210>1
<211>214
<212>prt
<213>水稻(oryzasatival.)
<400>1
metserleupheglyleuglyargasnglnlysthrpheargprolys
151015
lysseralaproserglyserlysglyalaglnleuarglyshisile
202530
aspalathrleuglyserglyasnleuargglualavalargleupro
354045
proglygluaspileasnglutrpleualavalasnthrvalaspphe
505560
pheasnglnvalasnleuleutyrglythrleualagluphecysthr
65707580
proglusercysprothrmetthralaglyprolystyrglutyrarg
859095
trpalaaspglyvalglnilelyslysproilegluvalseralapro
100105110
lystyrvalglutyrleumetasptrpilegluglyglnleuaspasp
115120125
gluserilepheproglnlysleuglythrpropheproproasnphe
130135140
lysgluvalvallysthrilephelysargleupheargvaltyrala
145150155160
hisiletyrhisserhispheglnlysilevalserleulysgluglu
165170175
alahisleuasnthrcysphelyshispheileleuphethrthrglu
180185190
pheglyleuileasplyslysgluleualaproleuglngluleuile
195200205
gluserileileprotyr
210
<210>2
<211>645
<212>dna
<213>水稻(oryzasatival.)
<400>2
atgagtctcttcgggctcggccggaatcagaagacgttccgtcccaagaagagtgctcct60
tcaggaagcaagggtgcacagcttcggaagcacatagatgccactcttggcagtggaaac120
cttagggaagctgtaaggttgccgcctggagaagatatcaatgaatggctggctgtgaat180
actgtggatttctttaaccaagttaacctgctgtacggcacactcgctgagttctgcaca240
cctgagagctgcccaacaatgactgctggcccaaagtatgagtacagatgggctgatggt300
gtacagataaagaagccgatagaagtatcagcaccaaaatatgtggagtacctaatggac360
tggattgaaggccaacttgatgatgaatctatatttccccaaaagcttggcacaccattc420
ccgccaaacttcaaggaggttgtaaagacaattttcaagcgcttgtttcgtgtttatgcc480
cacatatatcactcccattttcagaagattgtcagcctcaaggaggaggcccatcttaac540
acgtgcttcaagcacttcatcctgtttacaactgaatttggcctgattgacaagaaggag600
ctggctccactccaggagctcatcgaatcgattattccatactga645
起点商标作为专业知识产权交易平台,可以帮助大家解决很多问题,如果大家想要了解更多知产交易信息请点击 【在线咨询】或添加微信 【19522093243】 与客服一对一沟通,为大家解决相关问题。
与客服一对一沟通,为大家解决相关问题。
此文章来源于网络,如有侵权,请联系删除

 热门咨询
热门咨询



