
 商标分类
商标分类  商标转让
商标转让 
新颖腺相关病毒(AAV)载体、具有降低的衣壳脱酰胺化的AAV载体及其用途的制作方法
 2021-02-02 17:02:38|
2021-02-02 17:02:38| 307|
307| 起点商标网
起点商标网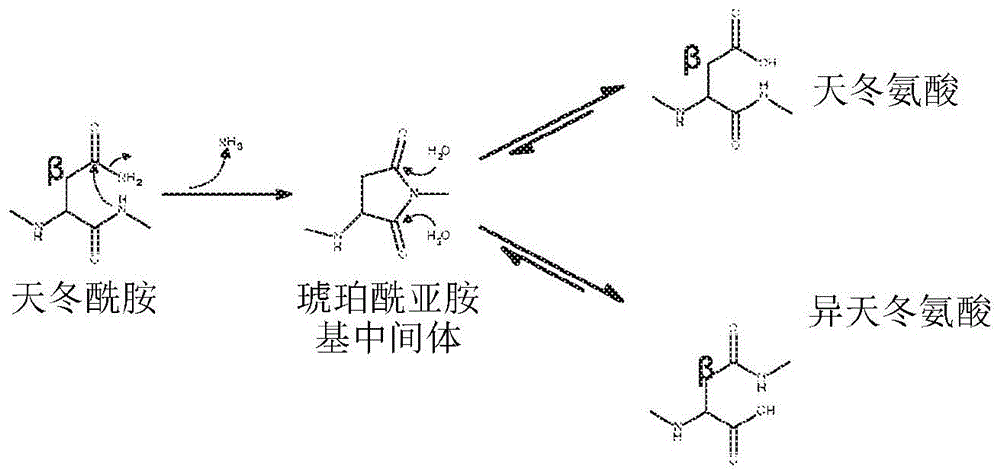
关于联邦政府资助研究的声明本发明是在国立卫生研究院授予的p01hl059407下由政府支持进行的。政府享有本发明的一定权利。对通过efs-web提交的序列表的引用创建于2019年2月27日并通过efs-web以电子方式提交的命名为“18-8592pct_sequence_listing_st25”的序列表的文本文件的内容在此与本申请通过引用整体并入本文中。
背景技术:
:腺相关病毒(aav)衣壳的结构为二十面体,并且包含比率为1:1:10的60个病毒蛋白(vp)单体(vp1、vp2和vp3)(xieq等人,《美国国家科学院院刊(procnatlacadsciusa)》2002;99(16):10405-10)。vp1和vp2两者的c端内均含有整个vp3蛋白序列(约535aa),并且共享的vp3序列主要负责总体衣壳结构。由于vp1/vp2独特区的结构柔性以及组装衣壳中vp1和vp2单体相对于vp3单体的低呈现,因此vp3是可通过x射线晶体学分辨的唯一衣壳蛋白(namhj等人,《病毒学杂志(jvirol)》2007;81(22):12260-71)。vp3含有作为aav血清型之间序列变异的主要来源的九个高变区(hvr)(govindasamyl等人,《病毒学杂志》2013;87(20):11187-99)。考虑到其在衣壳表面上的柔性和位置,hvr主要负责与靶细胞以及免疫系统相互作用(huangly等人,《病毒学杂志》2016;90(11):5219-30;rauppc等人,《病毒学杂志》2012;86(17):9396-408)。虽然公开了分别用于aav2、aavrh.8、aav6、aav9、aav3b、aav8和aav4的结构条目的多种血清型的结构(来自结构生物信息学研究合作实验室(rcsb)数据库的蛋白质数据库(pdb)id1lp3、4rso,4v86、3ux1、3kic、2qa0、2g8g),但文献中关于这些衣壳的表面上的修饰的信息很少。研究表明衣壳的细胞内磷酸化在特定酪氨酸残基处发生(zhongl等人,《病毒学(virology)》2008;381(2):194-202)。尽管初级vp3序列中存在推定的糖基化位点,但尚未在aav2中鉴定出糖基化事件(murrays等人,《病毒学杂志》2006;80(12):6171-6;jinx等人,《人类基因疗法方法(humgenethermethods)》2017;28(5):255-267);其它aav血清型尚未针对衣壳糖基化进行评估。aav基因疗法载体经历较少的分子水平审查,所述分子水平审查通常伴随重组蛋白治疗剂的开发和制造。aav衣壳转译后修饰(ptm)很大程度上尚未被开发,因此,对关于其影响功能的潜力或关于控制制造的aav疗法中ptm水平的策略知之甚少。非基因疗法蛋白治疗剂的转译后修饰的变化使得其开发作为药物变得复杂。jenkins,n、murphy,l和tyther,r(2008),“重组蛋白的转译后修饰:对于生物制药的意义(post-translationalmodificationsofrecombinantproteins:significanceforbiopharmaceuticals)”《分子生物技术(molbiotechnol)》39:113-118;houde,d、peng,y、berkowitz,sa和engen,jr(2010),“转译后修饰差异地影响igg1构象和受体结合(post-translationalmodificationsdifferentiallyaffectigg1conformationandreceptorbinding)”《分子细胞蛋白质组学(molcellproteomics)》9:1716-1728。例如,所选氨基酸的脱酰胺化调节了基于重组保护性抗原的炭疽疫苗的稳定性和免疫应答。(powellbs等人,《蛋白质(proteins)》2007;68(2):458-79;vermaa等人,《临床与疫苗免疫学(clinvaccineimmunol)》2016;23(5):396-402)。在一些情况下,此过程通过病毒或细菌脱酰胺酶进行催化,以调节宿主细胞信号传导途径或先天免疫应答(zhaoj等人,《病毒学杂志》2016;90(9):4262-8;zhaoj等人,《细胞宿主微生物(cellhostmicrobe)》2016;20(6):770-84)。更常见的是,内源性脱酰胺化是一种不依赖酶的自发过程。尽管尚未完全阐明自发脱酰胺化的目的,但先前的研究已经表明此事件用作分子时钟以指示蛋白质的相对年龄并调节其转换(robinsonne和robinsonab,《美国国家科学院院刊》2001;98(3):944-9)。脱酰胺化在天冬酰胺或更不常见的谷氨酰胺的酰胺基经历来自相邻氮原子的亲核攻击并且酰胺基丢失时发生。此过程产生琥珀酰亚胺基中间体(yangh和zubarevra,《电泳(electrophoresis)》2010;31(11):1764-72),所述琥珀酰亚胺基中间体通过水解分解为天冬氨酸和异天冬氨酸(或谷氨酸和异谷氨酸)的混合物(cataks等人,《物理化学杂志a(jphyschema)》2009;113(6):1111-20)。短合成肽的研究估计,这种水解会产生比例为3:1的异天冬氨酸与天冬氨酸混合物(geigert.和clarkes,《生物化学杂志(jbiolchem)》1987;262(2):785-94。仍然需要包括基于aav的构建体的组合物以递送具有稳定的受体结合和/或稳定的衣壳的异源分子,从而避免中和抗体和/或在储存时保持纯度。技术实现要素:在一个实施例中,提供了一种组合物,其包括重组腺相关病毒(raav)的混合群体,所述raav中的每个raav包括:(a)aav衣壳,所述aav衣壳包括约60个衣壳vp1蛋白、vp2蛋白和vp3蛋白,其中所述vp1蛋白、所述vp2蛋白和所述vp3蛋白为:vp1蛋白的异质群体,所述vp1蛋白由对所选aavvp1氨基酸序列进行编码的核酸序列产生;vp2蛋白的异质群体,所述vp2蛋白由对所选aavvp2氨基酸序列进行编码的核酸序列产生;vp3蛋白的异质群体,所述vp3蛋白由对所选aavvp3氨基酸序列进行编码的核酸序列产生,其中所述vp1蛋白、所述vp2蛋白和所述vp3蛋白含有具有氨基酸修饰的亚群体,所述氨基酸修饰包括所述aav衣壳中的天冬酰胺-甘氨酸对中的至少两个高度脱酰胺化的天冬酰胺(n),并且任选地进一步包括包含其它脱酰胺化的氨基酸的亚群体,其中脱酰胺化引起氨基酸变化;以及(b)所述aav衣壳中的载体基因组,所述载体基因组包括核酸分子,所述核酸分子包括aav反向末端重复序列和对产物进行编码的非aav核酸序列,所述非aav核酸序列可操作地连接到引导所述产物在宿主细胞中的表达的序列。在某些实施例中,所述脱酰胺化的天冬酰胺被脱酰胺化为天冬氨酸、异天冬氨酸、相互转化的天冬氨酸/异天冬氨酸对或其组合。在某些实施例中,所述衣壳进一步包括一种或多种脱酰胺化的谷氨酰胺,所述一种或多种脱酰胺化的谷氨酰胺被脱酰胺化为(α)-谷氨酸、γ-谷氨酸、相互转化的(α)-谷氨酸/γ-谷氨酸对或其组合。在另外的方面,提供了一种重组腺相关病毒(raav),所述raav包括:(a)aavrh79衣壳,所述aavrh79衣壳包括以下中的一种或多种:(1)aavrh79衣壳蛋白,所述aavrh79衣壳蛋白包括:aavrh79vp1蛋白的异质群体,所述aavrh79vp1蛋白选自以下:通过从对seqidno:2的1到738的预测的氨基酸序列进行编码的核酸序列的表达产生的vp1蛋白、从seqidno:1产生的vp1蛋白或从与seqidno:1至少70%相同的对seqidno:2的1到738的预测的氨基酸序列进行编码的核酸序列产生的vp1蛋白;aavrh79vp2蛋白的异质群体,所述aavrh79vp2蛋白选自以下:通过从对seqidno:2的至少约氨基酸138到738的预测的氨基酸序列进行编码的核酸序列的表达产生的vp2蛋白、从包括seqidno:1的至少核苷酸412到2214的序列产生的vp2蛋白或从与seqidno:1的至少核苷酸412到2214至少70%相同的对seqidno:2的至少约氨基酸138到738的预测的氨基酸序列进行编码的核酸序列产生的vp2蛋白;aavrh79vp3蛋白的异质群体,所述aavrh79vp3蛋白选自以下:通过从对seqidno:2的至少约氨基酸204到738的预测的氨基酸序列进行编码的核酸序列的表达产生的vp3蛋白、从包括seqidno:1的至少核苷酸610到2214的序列产生的vp3蛋白或从与seqidno:1的至少核苷酸610到2214至少70%相同的对seqidno:2的至少约氨基酸204到738的预测的氨基酸序列进行编码的核酸序列产生的vp3蛋白;和/或(2)vp1蛋白的异质群体,所述vp1蛋白作为对seqidno:2的氨基酸序列进行编码的核酸序列的产物;vp2蛋白的异质群体,所述vp2蛋白作为对seqidno:2的至少约氨基酸138到738的氨基酸序列进行编码的核酸序列的产物;以及vp3蛋白的异质群体,所述vp3蛋白作为对seqidno:2的至少氨基酸204到738进行编码的核酸序列的产物,其中:所述vp1蛋白、所述vp2蛋白和所述vp3蛋白含有具有氨基酸修饰的亚群体,所述氨基酸修饰包括seqidno:2中的天冬酰胺-甘氨酸对中的至少两个高度脱酰胺化的天冬酰胺(n),并且任选地进一步包括包含其它脱酰胺化的氨基酸的亚群体,其中脱酰胺化引起氨基酸变化;以及(b)所述aavrh79衣壳中的载体基因组,所述载体基因组包括核酸分子,所述核酸分子包括aav反向末端重复序列和对产物进行编码的非aav核酸序列,所述非aav核酸序列可操作地连接到引导所述产物在宿主细胞中的表达的序列。另一方面,提供了一种转导靶组织的方法。在一个实施例中,所述方法包含施用具有如本文所描述的aavrh79衣壳的aav。在一个实施例中,提供了一种转导肝脏组织的方法,所述方法包括施用具有aavrh79衣壳的aav。在另一个实施例中,提供了一种转导肌肉组织的方法,所述方法包括施用具有aavrh79衣壳的aav。在又另一方面,提供了一种降低aavrh79衣壳的脱酰胺化的方法。在一个实施例中,所述方法包含由含有经过修饰的aavrh79vp密码子的核酸序列产生aavrh79衣壳,所述核酸序列包括定位在seqidno:2中的位置n57、n263、n385和/或n514处的天冬酰胺-甘氨酸对中的一到四个天冬酰胺-甘氨酸对处的独立地经过修饰的甘氨酸密码子,使得经过修饰的密码子对除甘氨酸之外的氨基酸进行编码。在另一个实施例中,所述方法包含由含有经过修饰的aavrh79vp密码子的核酸序列产生aavrh79衣壳,所述核酸序列包括定位在seqidno:2的位置n94、n254、n305、n410和/或n479处的天冬酰胺-甘氨酸对中的一到四个天冬酰胺-甘氨酸对处的独立地经过修饰的甘氨酸密码子。在另外的实施例中,提供了一种raav8.ar2.08,其包括:(a)aav8.ar2.08衣壳,所述aav8.ar2.08衣壳包括以下中的一种或多种:(1)aav8.2.08衣壳蛋白,所述aav8.2.08衣壳蛋白包括:aav8.ar2.08vp1蛋白的异质群体,所述aav8.ar2.08vp1蛋白选自以下:通过从对seqidno:18的1到738的预测的氨基酸序列进行编码的核酸序列的表达产生的vp1蛋白、从seqidno:17产生的vp1蛋白或从与seqidno:17至少70%相同的对seqidno:18的1到738的预测的氨基酸序列进行编码的核酸序列产生的vp1蛋白;aav8.ar2.08vp2蛋白的异质群体,所述aav8.ar2.08vp2蛋白选自以下:通过从对seqidno:18的至少约氨基酸138到738的预测的氨基酸序列进行编码的核酸序列的表达产生的aav8.ar2.08vp2蛋白、从包括seqidno:17的至少核苷酸412到2214的序列产生的vp2蛋白或从与seqidno:17的至少核苷酸411到2214至少70%相同的对seqidno:18的至少约氨基酸138到738的预测的氨基酸序列进行编码的核酸序列产生的vp2蛋白;aav8.ar2.08vp3蛋白的异质群体,所述aav8.ar2.08vp3蛋白选自以下:通过从对seqidno:18的至少约氨基酸204到738的预测的氨基酸序列进行编码的核酸序列的表达产生的vp3蛋白、从包括seqidno:17的至少核苷酸607到2214的序列产生的vp3蛋白或从与seqidno:17的至少核苷酸607到2214至少70%相同的对seqidno:18的至少约氨基酸204到738的预测的氨基酸序列进行编码的核酸序列产生的vp3蛋白;和/或(2)vp1蛋白的异质群体,所述vp1蛋白作为对seqidno:18的氨基酸序列进行编码的核酸序列的产物;vp2蛋白的异质群体,所述vp2蛋白作为对seqidno:18的至少约氨基酸138到738的氨基酸序列进行编码的核酸序列的产物;以及vp3蛋白的异质群体,所述vp3蛋白作为对seqidno:18的至少氨基酸204到738进行编码的核酸序列的产物,其中:所述vp1蛋白、所述vp2蛋白和所述vp3蛋白含有具有氨基酸修饰的亚群体,所述氨基酸修饰包括seqidno:18中的天冬酰胺-甘氨酸对中的至少两个高度脱酰胺化的天冬酰胺(n),并且任选地进一步包括包含其它脱酰胺化的氨基酸的亚群体,其中脱酰胺化引起氨基酸变化;以及(b)所述aav8.ar2.08衣壳中的载体基因组,所述载体基因组包括核酸分子,所述核酸分子包括aav反向末端重复序列和对产物进行编码的非aav核酸序列,所述非aav核酸序列可操作地连接到引导所述产物在宿主细胞中的表达的序列。另一方面,提供了一种转导靶组织的方法。在一个实施例中,所述方法包含施用具有如本文所描述的aav8.ar2.08衣壳的aav。在一个实施例中,提供了一种转导肝脏组织的方法,所述方法包括施用具有aav8.ar2.08衣壳的aav。在另一个实施例中,提供了一种转导肌肉组织的方法,所述方法包括施用具有aav8.ar2.08衣壳的aav。在又另一方面,提供了一种降低aav8.ar2.08衣壳的脱酰胺化的方法。在一个实施例中,所述方法包含由含有经过修饰的aav8.ar2.08vp密码子的核酸序列产生aav8.ar2.08衣壳,所述核酸序列包括定位在seqidno:18中的位置n57、n263、n385、n514和/或n540处的天冬酰胺-甘氨酸对中的一到四个天冬酰胺-甘氨酸对处的独立地经过修饰的甘氨酸密码子,使得经过修饰的密码子对除甘氨酸之外的氨基酸进行编码。在另一个实施例中,所述方法包含由含有经过修饰的aav8.ar2.08vp密码子的核酸序列产生aav8.ar2.08衣壳,所述核酸序列包括定位在seqidno:18的位置n94、n254、n305、n521、n590、q601、n653和/或n665处的天冬酰胺-甘氨酸对中的一到四个天冬酰胺-甘氨酸对处的独立地经过修饰的甘氨酸密码子。在某些实施例中,提供了一种raav5.5.9,其包括:(a)aav5.5.9衣壳,所述aav5.5.9衣壳包括以下中的一种或多种:(1)aavg5衣壳蛋白,所述aavg5衣壳蛋白包括:aav5.5.9vp1蛋白的异质群体,所述aav5.5.9vp1蛋白选自以下:通过从对seqidno:10的1到736的预测的氨基酸序列进行编码的核酸序列的表达产生的vp1蛋白、从seqidno:9产生的vp1蛋白或从与seqidno:9至少70%相同的对seqidno:1的1到736的预测的氨基酸序列进行编码的核酸序列产生的vp1蛋白;aav5.5.9vp2蛋白的异质群体,所述aav5.5.9vp2蛋白选自以下:通过从对seqidno:10的至少约氨基酸138到736的预测的氨基酸序列进行编码的核酸序列的表达产生的aavg5vp2蛋白、从包括seqidno:9的至少核苷酸412到2211的序列产生的vp2蛋白或从与seqidno:9的至少核苷酸412到2211至少70%相同的对seqidno:10的至少约氨基酸138到736的预测的氨基酸序列进行编码的核酸序列产生的vp2蛋白;aav5.5.9vp3蛋白的异质群体,所述aav5.5.9vp3蛋白选自以下:通过从对seqidno:10的至少约氨基酸203到736的预测的氨基酸序列进行编码的核酸序列的表达产生的aav5.5.9vp3蛋白、从包括seqidno:9的至少核苷酸607到2211的序列产生的vp3蛋白或从与seqidno:9的至少核苷酸607到2211至少70%相同的对seqidno:10的至少约氨基酸203到736的预测的氨基酸序列进行编码的核酸序列产生的vp3蛋白;和/或(2)vp1蛋白的异质群体,所述vp1蛋白作为对seqidno:10的氨基酸序列进行编码的核酸序列的产物;vp2蛋白的异质群体,所述vp2蛋白作为对seqidno:10的至少约氨基酸138到736的氨基酸序列进行编码的核酸序列的产物;以及vp3蛋白的异质群体,所述vp3蛋白作为对seqidno:10的至少氨基酸203到726进行编码的核酸序列的产物,其中:所述vp1蛋白、所述vp2蛋白和所述vp3蛋白含有具有氨基酸修饰的亚群体,所述氨基酸修饰包括seqidno:10中的天冬酰胺-甘氨酸对中的至少两个高度脱酰胺化的天冬酰胺(n),并且任选地进一步包括包含其它脱酰胺化的氨基酸的亚群体,其中脱酰胺化引起氨基酸变化;以及(b)所述aav5.5.9衣壳中的载体基因组,所述载体基因组包括核酸分子,所述核酸分子包括aav反向末端重复序列和对产物进行编码的非aav核酸序列,所述非aav核酸序列可操作地连接到引导所述产物在宿主细胞中的表达的序列。另一方面,提供了一种转导靶组织的方法。在一个实施例中,所述方法包含施用具有如本文所描述的aav5.5.9衣壳的aav。在一个实施例中,提供了一种转导肝脏组织的方法,所述方法包括施用具有aav5.5.9衣壳的aav。在另一个实施例中,提供了一种转导肌肉组织的方法,所述方法包括施用具有aav5.5.9衣壳的aav。在又另一方面,提供了一种降低aav5.5.9衣壳的脱酰胺化的方法。在一个实施例中,所述方法包含由含有经过修饰的aav5.5.9vp密码子的核酸序列产生aav5.5.9衣壳,所述核酸序列包括定位在seqidno:10中的位置n57、n319、n442和/或n502处的天冬酰胺-甘氨酸对中的一到四个天冬酰胺-甘氨酸对处的独立地经过修饰的甘氨酸密码子,使得经过修饰的密码子对除甘氨酸之外的氨基酸进行编码。在另一个实施例中,所述方法包含由含有经过修饰的aav5.5.9vp密码子的核酸序列产生aav5.5.9衣壳,所述核酸序列包括定位在seqidno:10的位置n35、n113、n204、n217、n243、n249、n293/294、n304、n399/400、n505、q589、n618、n641、n653、n658和/或n699处的天冬酰胺-甘氨酸对中的一到四个天冬酰胺-甘氨酸对处的独立地经过修饰的甘氨酸密码子。另一方面,提供了一种组合物,所述组合物包括如本文所描述的重组aavrh79、aav8.ar2.08或aav5.5.9的混合群体。在又另一方面,提供了如本文所描述的重组aav(raav),所述raav用于将所期望的基因产物递送到有需要的受试者。另一方面,提供了一种可用于产生如本文所描述的raav的raav产生系统。在一个实施例中,所述系统包含:(a)aavrh79、aav8.ar2.08或aav5.5.9衣壳核酸序列,所述衣壳核酸序列对seqidno:2、10或18的预测的氨基酸序列进行编码;(b)适合于包装到所述aav衣壳中的核酸分子,所述核酸分子包括至少一个aav反向末端重复序列(itr)和对基因产物进行编码的非aav核酸序列,所述非aav核酸序列可操作地连接到引导所述产物在宿主细胞中的表达的序列;以及(c)足以允许将所述核酸分子包装到重组aav衣壳中的aavrep功能和辅助功能。通过以下对本发明的详细描述,本发明的这些和其它方面将变得显而易见。附图说明图1a-图1g:aav8vp同种型的电泳分析。(图1a)图展示了天冬酰胺残基通过相邻氮原子经历亲核攻击从而形成琥珀酰亚胺基中间体的机制。然后此中间体经历水解,从而分解为天冬氨酸和异天冬氨酸的混合物。如此标记β碳。图是在bioviadraw2018中产生的。(图1b)在变性的一维sds-page上运行1μg的aav8载体。(图1c)示出了碳酸酐酶pi标志物点的等电点。(图1d)通过二维凝胶电泳分析5μg的aav8载体,并用考马斯蓝对其进行染色。点1-20是氨基甲酰化的碳酸酐酶pi标志物。加框区如下:a=vp1,b=vp2,c=vp3,d=内部原肌球蛋白标志物(箭头:原肌球蛋白点的mw=33kda,pi=5.2)。在4-8的pi范围内进行等电聚焦。(图1e-图1g)在4-8的pi范围内进行的等电聚焦的结果。通过2d凝胶电泳分析1e11gc的wtaav8载体(图1e)或突变载体(图1f和图1g),并用syproruby对其进行染色。蛋白质标记:a=vp1;b=vp2;c=vp3,d=鸡蛋清伴清蛋白标记物,e=turbouclease标记物。在6-10的pi范围内进行等电聚焦。初级vp1/2/3同种型点被圈出,并且标志物的主要点的迁移距离由竖直线指示(turbonuclease=虚线,伴清蛋白=实线)。图2a-图2e:aav8衣壳蛋白中的天冬酰胺和谷氨酰胺脱酰胺化的分析。(图2a-图2b)示出了电喷雾电离(esi)质谱以及含有asn-94(图2a)和asp-94(图2b)的3+肽(93-103)的理论和观察到的质量。(图2c-图2d)示出了esi质谱以及含有asn-254(图2c)和asp-254(图2d)的3+肽(247-259)的理论和观察到的质量。asn-94和asn-254的观察到的质量偏移分别为0.982da和0.986da,而理论质量偏移为0.984da。(图2e)示出了通过不同方法纯化的aav8胰蛋白酶肽在所关注的特定天冬酰胺残基和谷氨酰胺残基处的脱酰胺化百分比。用交叉阴影线标记指示具有n+1个甘氨酸的天冬酰胺残基处的脱酰胺化的条。包含在所分析的至少一种制剂中确定为至少2%脱酰胺化的残基。数据表示为平均值±标准偏差。图3a-图3e:aav8vp3单体的结构建模和脱酰胺化的位点的分析。(图3a)以卷曲表示形式示出了aav8vp3单体(pdb标识符:3ra8)。带的颜色指示相对柔性程度(蓝色=最高刚性/正常温度系数,红色=最高柔性/高温系数)。球体指示所关注的残基。展开图是所关注残基及其周围残基的球状和棒状表示,以展示局部蛋白质结构(蓝色=氮,红色=氧)。带下划线的残基是ng基序中的残基。图3b-图3e:示出了具有n+1个甘氨酸的脱酰胺化的天冬酰胺的异天冬氨酸模型。与(图3c)n263、(图3d)n514和(图3e)n540的异天冬氨酸模型相比,用(图3b)n410的天冬酰胺模型从aav8晶体结构(pdbid:3ra8)的精修产生2fofc电子密度图(1σ水平)。电子密度图以洋红色网格示出。如此标记β碳。箭头指示与所关注的残基的r基团对应的电子密度。图4a-图4d:影响aav8衣壳脱酰胺化的因素的确定。将aav8制剂(图4a)在70℃下温育三天或七天,(图4b)暴露于ph2或ph10持续七天,或使用d2o代替h2o来制备(图4c)以用于质谱,以确定对aav衣壳形成而言不是固有的脱酰胺化的可能来源。(图4d)使用b1抗体(与变性的衣壳反应)和aav8构象特异性抗体(与完整衣壳反应)如图4a那样处理载体的斑点印迹以评估衣壳结构完整性。图5a-图5b:非aav蛋白中的脱酰胺化频率。为了与aav脱酰胺化百分比进行比较,示出了两种含有可能被脱酰胺化的ng基序的非aav重组蛋白(人碳酸酐酶(图5a)和大鼠苯丙氨酸-羟化酶(图5b))的脱酰胺化百分比。图6:使用来自两个机构的数据分析流水线计算的aav8脱酰胺化百分比的比较。示出了在两个不同机构处评估的aav8胰蛋白酶肽在所关注的特定天冬酰胺残基和谷氨酰胺残基处的脱酰胺化百分比。图7a-图7c展示了在批次之间具有高变异性的非ng位点处的功能性天冬酰胺取代。(图7a)通过在293个细胞中的小规模三重转染产生wtaav8载体和突变载体的效价,如通过定量pcr(qpcr)测量的。报告了相对于wtaav8对照物的效价。如图8b中所描述的测量转导效率。使效价和转导效率相对于wtaav8对照物的值归一化。(图7b)示出了接受wtaav8.cb7.ffluc和n499q衣壳突变载体的小鼠在注射后第14天的代表性荧光素酶图像。(图7c)在研究期间的第14天,通过荧光素酶成像测量来自用wtaav8载体或突变载体(n=3或4)静脉内注射的c57bl/6小鼠的荧光素酶表达,并以总通量单位报告。所有数据表示为平均值+标准偏差。图8a和图8b示出了基因脱酰胺化对载体性能的影响的体外分析结果。(图8a)通过在293个细胞中的小规模三重转染产生wtaav8载体和基因脱酰胺化突变载体的效价,如通过定量pcr(qpcr)测量的。报告了相对于wtaav8对照物的效价。用wtaav8和阴性对照物呈现高脱酰胺化的ng位点(图案化条)、低脱酰胺化的位点(白条)和高度可变的位点(黑条)。(图8b)相对于wtaav8对照物报告的产生萤火虫荧光素酶的突变aav8载体的转导效率。转导效率以添加到huh7细胞中的每个gc产生的发光单位来测量,并通过在多个稀释度下用粗载体进行转导来确定。使转导效率数据相对于wt参考归一化。所有数据表示为平均值±标准偏差。图9a-图9d展示了随着时间的推移,载体活性丧失与进行性脱酰胺化有关。(图9a)持续三重转染的hek293细胞的时程的载体产生(dnasei抗性基因组拷贝,gc)产生包装荧光素酶报告基因的aav8载体。使gc水平相对于最大观察值归一化。(图9b)使用经过纯化的时程载体转导huh7细胞。使用多个稀释度的经过纯化的时程载体样品如图8b中那样测量转导效率(以添加到靶细胞的每个gc的发光单位)。误差条表示对于每个采样时间进行至少10次技术重复的标准偏差。转染后1天、2天和5天收集的载体的aav8ng位点(图9c)和非ng位点(图9d)的脱酰胺化。图10a-图10d展示了稳定天冬酰胺对载体性能的影响。图10a示出了通过在293个细胞中的小规模三重转染产生wtaav8载体和+1位置突变载体的效价,如通过定量pcr(qpcr)测量的。报告了相对于wtaav8对照物的效价。图10b示出了相对于wtaav8对照物报告的产生萤火虫荧光素酶的突变aav8载体的转导效率。使用粗载体材料如图8b中那样测量转导效率。运行双样品t测试(*p<0.005)以确定wtaav8与g264a/g515a和g264a/g541a的突变转导效率之间的显著性。图10c示出了在研究期间的第14天,通过荧光素酶成像测量的来自用wtaav8载体或突变载体(n=3到5)静脉内注射的c57bl/6小鼠的肝脏区中的荧光素酶表达,并以总通量单位报告。图10d示出了相对于wtaav8对照物报告的产生萤火虫荧光素酶的多位点aav8突变载体的效价和转导效率。所有数据表示为平均值±标准偏差。图11a-图11c:aav9衣壳蛋白中的天冬酰胺和谷氨酰胺脱酰胺化的分析。(图11a)通过2d凝胶电泳分析1e11gc的wtaav9,并用syproruby对其进行染色。蛋白质标记:a=vp1;b=vp2;c=vp3,d=鸡蛋清伴清蛋白标记物,e=turbouclease标记物。在6-10的pi范围内进行等电聚焦。(图11b)示出了通过不同方法纯化的aav9胰蛋白酶肽在所关注的特定天冬酰胺残基和谷氨酰胺残基处的脱酰胺化百分比。用交叉阴影线标记指示具有n+1个甘氨酸的天冬酰胺残基处的脱酰胺化的条。包含在所分析的至少一种制剂中确定为至少2%脱酰胺化的残基。数据表示为平均值±标准偏差。(图11c)在由aav9晶体结构(pdbid:3ux1)的未偏置精修产生的2fofc电子密度图中示出了n512的异天冬氨酸模型。箭头指示与残基n512的r基团对应的电子密度。图11d-图11f:影响aav9衣壳脱酰胺化的因素的确定。(图11d)将两种aav9制剂在70℃下温育三天或七天,或(图11f)暴露于ph2或ph10持续七天,以确定对aav衣壳形成而言不是固有的脱酰胺化的可能来源。数据表示为平均值±标准偏差。(图11f)使用b1抗体(与变性的衣壳反应)如图11d那样处理载体的斑点印迹以评估衣壳结构完整性。图11g和图11h展示了aav9的基因脱酰胺化对载体性能的影响的体外分析。(图11g)通过在293个细胞中的小规模三重转染产生wtaav9载体和基因脱酰胺化突变载体的效价,如通过定量pcr(qpcr)测量的。报告了相对于wtaav9对照物的效价。用wtaav8和阴性对照物呈现高脱酰胺化的ng位点(图案化条)、低脱酰胺化的位点(白条)和高度可变的位点(黑条)。(图11h)相对于wtaav9对照物,报告了产生萤火虫荧光素酶的突变aav9载体的转导效率。所有数据表示为平均值±标准偏差。图11i-图11k示出了随着时间的推移的aav9载体体外效力。(图11i)持续三重转染的hek293细胞的时程的载体产生(dnasei抗性基因组拷贝,gc)产生包装荧光素酶报告基因的aav9载体。使gc水平相对于最大观察值归一化。(图11j)使用粗时程载体转导huh7细胞。(图11k)示出了针对粗载体样品和经过纯化的载体样品的转染后1天对比转染后5天收集的载体的转导效率。转导效率表达为荧光素酶活性/gc,相对于第1天的值归一化。图12a提供了使用用于比对的clustalω1.2.2及其默认参数制备的aav5.5.9[seqidno:10](有时也被称为aavg5)、aav9[seqidno:4]和aavphp.b[seqidno:12]的氨基酸序列的比对。图12b-12e提供了aav5.5.9[seqidno:9]、php.b[seqidno:11]、aav9[seqidno:3]和aavhu68[seqidno:14]的核苷酸序列的比对。图13a提供了使用用于比对的clustalω1.2.2及其默认参数制备的aav8三重突变体(aav8t)[seqidno:16]、aav8.ar2.08[seqidno:18](有时也被称为aavg3或ar2或aav.ar2)和aav8[seqidno:20]的氨基酸序列的比对。图13b-13d提供了aav8三重突变体[seqidno:15]、aav8.ar2.08[seqidno:17]、aav8[seqidno:19]的核苷酸序列的比对。图14a提供了使用用于比对的clustalω1.2.2及其默认参数的aavrh79[seqidno:2](有时也被称为aavg2)、aavrh10[seqidno:24]和aavhu37[seqidno:22]的氨基酸序列的比对。图14b-14d提供了aavrh79[seqidno:1]、aavrh10[seqidno:22]和aavhu37[seqidno:21]的核苷酸序列的比对。图15a和15b展示了在参考的载体的小规模或大规模制剂中的aav8三重、aavhu68、aav9、aav9和aavrh79的产生产率。图16提供了图15b的大规模制剂的产生纯度。图17a至17d示出了使用表达萤火虫荧光素酶的载体将3×1011gc/小鼠肌内(im)施用于雄性c57bl/6小鼠(n=5/组)的腓肠肌后,荧光素酶在肝脏和肌肉组织中的表达。图18a示出了实例5中用于条形码实验的质粒。图18b示出了注射到6只黑色小鼠的每种aav条形码变体的量。处死动物,采集组织样品,并从这些动物中的每只动物中分离出dna。图18c中示出了三只动物的总载体分布。图18d中示出了注射的载体混合物的实际频率对比理论频率。图19a-图20c示出了实例5的条形码生物分布实验的结果。分析单独组织样品的针对基因组和cdna的样品中的单个条形码频率对比注射混合物中的单个条形码频率。示出了针对肌肉(图19a,19b);心脏(图19c和图19d)和肝脏(图19e和图19f)的结果。图20a-20c中示出了与理论频率相比的倍数变化。图21和22示出了与aav8相比,aav8.ar2.08在小鼠中的生物分布。结果表明,aav8.ar2.08比aav8具有更高的肝脏特异性。图23比较了aav8对比aav8.ar2.08对比aavrh79的与可制造性有关的效价和产率。图24示出了与aav8(中条和右条)相比,aav8.ar2.08在组织中的生物分布(最左边的条)。图25-28示出了将aav载体施用于非人灵长类动物后的结果。图25提供了用于研究的载体和动物的细节。图26定量了从接受aav8、aavrh79或aav8.ar2.08载体的动物的肝脏中检测到的gc和gfp的水平。图27总结了hnp肝脏中的gfp表达的水平。图28示出了从施用aav8、aavrh79或aav8.ar2.08载体的hnp的组织中检测到的载体的水平。图29示出了在各种组织中检测到的aavrh79载体的生物分布。具体实施方式本文提供了一种重组腺相关病毒(raav)以及含有其的组合物,在重组aav的衣壳内存在的三个衣壳蛋白群体vp1、vp2和vp3中的每个衣壳蛋白群体中均具有序列和电荷异质性。本文提供了新颖raav以及用于降低脱酰胺化的方法和任选地其它衣壳单体修饰。本文进一步提供了具有减少的修饰的经过修饰的raav,所述减少的修饰可用于提供具有保持更大的稳定性、效力和/或纯度的衣壳的raav。“重组aav”或“raav”是含有两个元件的抗dnase的病毒颗粒、aav衣壳和至少含有包装在aav衣壳内的非aav编码序列的载体基因组。除非另有说明,否则此术语可以与短语“raav载体”互换使用。raav是“复制缺陷型病毒”或“病毒载体”,因为其缺少任何功能性aavrep基因或功能性aavcap基因并且不能产生后代。在某些实施例中,仅aav序列是aav反向末端重复序列(itr),通常定位在载体基因组的5'和3'最末端处,以允许定位在itr之间的基因和调节序列包装在aav衣壳内。如本文所使用的,“载体基因组”是指包装在形成病毒颗粒的raav衣壳内部的核酸序列。这种核酸序列含有aav反向末端重复序列(itr)。在本文的实例中,载体基因组至少含有从5'到3'的aav5'itr、一个或多个编码序列和aav3'itr。可以选择来自aav2的itr、不同于衣壳来源或除全长itr之外的aav。在某些实施例中,itr来自与在产生或反式补充aav期间提供rep功能的aav来源相同的aav。进一步地,可以使用其它itr。进一步地,载体基因组含有引导基因产物表达的调节序列。在本文中更详细地讨论载体基因组的合适组分。raav由aav衣壳和载体基因组构成。aav衣壳是vp1的异质群体、vp2的异质群体和vp3蛋白的异质群体的组装。如本文所使用的,当用于指vp衣壳蛋白时,术语“异源”或其任何语法变型是指由不相同的元件组成的群体,例如具有含有不同的经过修饰的氨基酸序列的vp1、vp2或vp3单体(蛋白)。如本文所使用的,与vp1、vp2和vp3蛋白(可替代地被称为同种型)结合使用的术语“异质群体”是指衣壳内的vp1、vp2和vp3蛋白的氨基酸序列中的差异。aav衣壳含有具有来自预测的氨基酸残基的修饰的vp1蛋白内、vp2蛋白内和vp3蛋白内的亚群体。这些亚群体至少包含某些脱酰胺化的天冬酰胺(n或asn)残基。例如,某些亚群体包括天冬酰胺-甘氨酸对中的至少一个、两个、三个或四个高度脱酰胺化的天冬酰胺(n)位置,并且任选地进一步包括其它脱酰胺化的氨基酸,其中脱酰胺化引起氨基酸变化和其它任选的修饰。如本文所使用的,除非另有说明,否则vp蛋白的“亚群体”是指一组vp蛋白,所述一组vp蛋白具有至少一个限定的特性,并且由至少一个组成员到少于参考组的所有成员组成。例如,除非另有说明,否则vp1蛋白的“亚群体”可以是至少一种(1)vp1蛋白,并且少于组装的aav衣壳中的所有vp1蛋白。除非另有说明,否则vp3蛋白的“亚群体”可以是一种(1)vp3蛋白到少于组装的aav衣壳中的所有vp3蛋白。例如,vp1蛋白可以是vp蛋白的亚群体;vp2蛋白可以是vp蛋白的不同单独的亚群体,并且vp3是组装的aav衣壳中的vp蛋白的又另外的亚群体。在另一个实例中,vp1、vp2和vp3蛋白可以含有具有不同修饰的亚群体,所述修饰例如至少一个、两个、三个或四个高度脱酰胺化的天冬酰胺,例如在天冬酰胺-甘氨酸对处。除非另有说明,否则高度脱酰胺化的是指与在参考氨基酸位置处的预测的氨基酸序列相比,在参考的氨基酸位置处被至少45%脱酰胺化的、至少50%脱酰胺化的、至少60%脱酰胺化的、至少65%脱酰胺化的、至少70%、至少75%、至少80%、至少85%、至少90%、至少95%、至少97%、至少99%或最高约100%脱酰胺化的(例如,在seqidno:2的氨基酸57处的天冬酰胺的至少80%可以基于总vp1蛋白脱酰胺化或可以基于总vp1、vp2和vp3蛋白脱酰胺化)。此类百分比可以使用2d凝胶、质谱技术或其它合适的技术来确定。在不希望受理论束缚的情况下,aav衣壳中的vp蛋白中的至少高度脱酰胺化的残基的脱酰胺化被认为本质上主要是非酶促的,这是由衣壳蛋白内的功能基团引起的,所述功能基团使所选天冬酰胺脱酰胺化,并在较小程度上使谷氨酰胺残基脱酰胺化。大多数脱酰胺化vp1蛋白的有效衣壳组装表明这些事件在衣壳组装之后发生,或者单独单体(vp1、vp2或vp3)中的脱酰胺化在结构上具有良好的耐受性,并且在很大程度上不会影响组装动力学。vp1独特(vp1-u)区(约aa1-137)中的广泛的脱酰胺化通常被认为在细胞进入之前定位在内部,这表明vp脱酰胺化可以在衣壳组装之前发生。在不希望受理论束缚的情况下,n的脱酰胺化可以通过其c端残基的骨架氮原子对asn的侧链酰胺基碳原子进行亲核攻击而发生。据信形成了中间闭环的琥珀酰亚胺残基。然后,琥珀酰亚胺残基进行快速水解以产生最终产物天冬氨酸(asp)或异天冬氨酸(isoasp)。因此,在某些实施例中,天冬酰胺(n或asn)的脱酰胺化产生asp或isoasp,其可以通过琥珀酰亚胺中间体相互转化,例如如下文所展示的。如本文所提供的,vp1、vp2或vp3中的每个脱酰胺化的n可以独立地是天冬氨酸(asp)、异天冬氨酸(isoasp)、天冬氨酸和/或asp和isoasp的互相转化的共混物或其组合。可以存在任何合适比率的α-和异天冬氨酸。例如,在某些实施例中,比率可以为10:1到1:10天冬氨酸:异天冬氨酸、约50:50天冬氨酸:异天冬氨酸或约1:3天冬氨酸:异天冬氨酸或另一所选比率。在某些实施例中,一种或多种谷氨酰胺(q)可以脱酰胺化为谷氨酸(glu),即α-谷氨酸、γ-谷氨酸(glu)或α-和γ-谷氨酸的共混物,其可以通过常见的戊二酰亚胺(glutarinimide)中间体相互转化。可以存在任何合适比率的α-和γ-谷氨酸。例如,在某些实施例中,比率可以为10:1到1:10α:γ、约50:50α:γ或约1:3α:γ或另一所选比率。因此,raav包含vp1、vp2和/或vp3蛋白的raav衣壳内具有脱酰胺化的氨基酸的亚群体,至少包含至少一种包括至少一个高度脱酰胺化的天冬酰胺的亚群体。另外,其它修饰可以包含异构化,特别是在所选天冬氨酸(d或asp)残基位置处。在仍其它实施例中,修饰可以包含在asp位置处的酰胺化。在某些实施例中,aav衣壳含有具有至少1个、至少2个、至少3个、至少4个、至少5个到至少约25个脱酰胺化的氨基酸残基位置的vp1、vp2和vp3的亚群体,与vp蛋白的经过编码的氨基酸序列相比,所述vp1、vp2和vp3的至少1%到10%、至少10%到25%、至少25%到50%、至少50%到70%、至少70%到100%、至少75%到100%、至少80%到100%或至少90%到100%被脱酰胺化。这些中的大多数可以是n残基。然而,q残基也可以被脱酰胺化。如本文所使用的,“经过编码的氨基酸序列”是指基于被转译成氨基酸的参考的核酸序列的已知dna密码子的转译而预测的氨基酸。下表展示了dna密码子和二十种常见氨基酸,分别示出了单字母代码(slc)和三个字母代码(3lc)。在某些实施例中,raav具有含有vp1、vp2和vp3蛋白的aav衣壳,所述蛋白具有包括在本文中提供的表中列出的位置处的两个、三个、四个、五个或更多个脱酰胺化的残基的组合的亚群体,并通过引用并入本文中。raav中的脱酰胺化可以使用2d凝胶电泳和/或质谱和/或蛋白质建模技术来确定。在线色谱可以使用acclaimpepmap柱和与qexactivehf和nanoflex源(赛默飞世尔科技公司(thermofisherscientific))联接的thermoultimate3000rslc系统(赛默飞世尔科技公司)执行。ms数据是使用用于qexactivehf的数据依赖性前20种方法获取的,所述方法从调查扫描(200-2000m/z)中动态选择最丰富的尚未测序的前体离子。测序通过高能碰撞解离片段进行,其中通过预测性自动增益控制确定的靶值为1e5离子,并且以4m/z的窗口进行前体分离。以m/z200以120,000的分辨率获取调查扫描。hcd光谱的分辨率可以在m/z200下设置为30,000,其中最大离子注入时间为50毫秒,并且归一化碰撞能量为30。s-透镜rf水平可以设置为50,以使消化肽所占据的m/z区达到最佳传输。可以从片段选择中排除具有单个、未分配或六个和更高电荷状态的前体离子。biopharmafinder1.0软件(赛默飞世尔科技公司)可以用于分析所获取的数据。对于肽作图,使用单进入蛋白fasta数据库进行搜索,其中脲基甲基化设置为固定的修饰;并将氧化、脱酰胺化和磷酸化设置为可变修饰、10ppm质量准确度、高蛋白酶特异性和置信水平为0.8的ms/ms光谱。合适的蛋白酶的实例可以包含例如胰蛋白酶或胰凝乳蛋白酶。脱酰胺化的肽的质谱鉴定相对简单,因为脱酰胺化向完整分子的质量添加了+0.984da(-oh基团与-nh2基团之间的质量差)。特定肽的脱酰胺化百分比通过将脱酰胺化的肽的质量面积除以脱酰胺化的和天然的肽的面积之和来确定。考虑到可能的脱酰胺化位点的数量,在不同位点处被脱酰胺化的同量异位物种可以在单个峰中共迁移。因此,源自具有多个潜在的脱酰胺化位点的肽的片段离子可以用于定位或区分多个脱酰胺化位点。在这些情况下,观察到的同位素图案内的相对强度可以用于特异性确定不同的脱酰胺化的肽异构体的相对丰度。此方法假设所有异构物种的片段化效率是相同的,并且在脱酰胺化位点上是独立的。本领域的技术人员将理解的是,可以使用这些说明性方法的多种变型。例如,合适的质谱仪可以包含例如四极杆飞行时间质谱仪(qtof),如watersxevo或agilent6530,或orbitrap仪器,如orbitrapfusion或orbitrapvelos(赛默飞世尔科技公司)。合适的液相色谱系统包含例如来自沃特世(waters)的acquityuplc系统或agilent系统(1100或1200系列)。合适的数据分析软件可以包含例如masslynx(沃特世)、pinpoint和petfinder(赛默飞世尔科技公司)、mascot(矩阵科学公司(matrixscience))、peaksdb(生物信息学解决方案公司(bioinformaticssolutions))。可以在例如x.jin等人,于2017年6月16日在线公开的《人类基因疗法方法》,第28卷,第5期,第255-267页中描述仍其它技术。除脱酰胺化之外,可能发生不会导致一个氨基酸转化为不同的氨基酸残基的其它修饰。这种修饰可以包含乙酰化残基、异构化、磷酸化或氧化。脱酰胺化的调节:在某些实施例中,aav被修饰成改变天冬酰胺-甘氨酸对中的甘氨酸,以降低脱酰胺化。在其它实施例中,将天冬酰胺改变为不同的氨基酸,例如以较慢速率进行脱酰胺化的谷氨酰胺;或改变为缺乏酰胺基的氨基酸(例如,含有酰胺基的谷氨酰胺和天冬酰胺);和/或改变为缺乏酰胺基的氨基酸(例如,含有胺基的赖氨酸、精氨酸和组氨酸)。如本文所使用的,缺乏酰胺或胺侧基的氨基酸是指例如甘氨酸、丙氨酸、缬氨酸、亮氨酸、异亮氨酸、丝氨酸、苏氨酸、胱氨酸、苯丙氨酸、酪氨酸或色氨酸和/或脯氨酸。如所描述的修饰可以在经过编码的aav氨基酸序列中存在的天冬酰胺-甘氨酸对中的一个、两个或三个天冬酰胺-甘氨酸对中。在某些实施例中,在所有四个天冬酰胺-甘氨酸对中没有进行此类修饰。因此,一种用于降低具有较低脱酰胺化速率的aav和/或工程化aav变体的脱酰胺化的方法。另外地或可替代地,可以将一种或多种其它酰胺氨基酸改变为非酰胺氨基酸以降低aav的脱酰胺化。在某些实施例中,本文所描述的突变aav衣壳含有天冬酰胺-甘氨酸对中的突变,使得甘氨酸变为丙氨酸或丝氨酸。突变aav衣壳可以含有一个、两个或三个突变体,其中参考aav天然地含有四个ng对。在某些实施例中,aav衣壳可以含有一个、两个、三个或四个此类突变体,其中参考aav天然地含有五个ng对。在某些实施例中,突变aav衣壳含有ng对中的仅单个突变。在某些实施例中,突变aav衣壳含有两个不同的ng对中的突变。在某些实施例中,突变aav衣壳含有定位在aav衣壳中的结构上分开的位置中的两个不同的ng对中的突变。在某些实施例中,突变不在vp1独特区中。在某些实施例中,突变中的一个突变不在vp1独特区中。任选地,突变aav衣壳不含有ng对中的修饰,但是含有突变以最小化或消除定位在ng对的外部的一个或多个天冬酰胺或谷氨酰胺中的脱酰胺化。在某些实施例中,提供了一种增加raav载体的效力的方法,所述方法包括使aav衣壳工程化,这消除了野生型aav衣壳中的ng中的一个或多个ng。在某些实施例中,“ng”的“g”的编码序列被工程为对另一种氨基酸进行编码。在下文的某些实例中,“s”或“a”被取代。然而,可以选择其它合适的氨基酸编码序列。参见例如下表,其中基于aav8的编号,修饰针对以下位置中的至少一个位置的编码序列:n57+1、n263+1、n385+1、n514+1、n540+1,或如下表所示出的。在某些实施例中,aav8突变体避免改变位置n57、n94、n263、n305、q467、n479和/或n653处的ng对。在某些实施例中,其它aav避免了在使用aav8编号作为参考如基于与aav8的比对所确定的对应n位置处的突变。这些氨基酸修饰可以通过常规的基因工程技术进行。例如,可以产生含有经过修饰的aavvp密码子的核酸序列,其中修饰精氨酸-甘氨酸对中对甘氨酸进行编码的密码子中的一到三个密码子以对除甘氨酸之外的氨基酸进行编码。在某些实施例中,含有经过修饰的精氨酸密码子的核酸序列可以在精氨酸-甘氨酸对中的一到三个精氨酸-甘氨酸对处被工程化,使得经过修饰的密码子对除精氨酸之外的氨基酸进行编码。每个经过修饰的密码子可以对不同的氨基酸进行编码。可替代地,改变的密码子中的一个或多个密码子可以对相同的氨基酸进行编码。在某些实施例中,这些经过修饰的aavrh79、aav8.ar2.08或aav5.5.9核酸序列可以用于产生具有比天然aavrh79、aav8.ar2.08或aav5.5.9衣壳脱酰胺化程度更低的衣壳的突变raav。此类突变raav可以具有降低的免疫原性和/或增加储存时的稳定性,特别是以悬浮液形式储存时的稳定性。本文还提供了对具有降低的脱酰胺化的aav衣壳进行编码的核酸序列。设计对此aav衣壳进行编码的核酸序列在本领域的技术范围内,包含dna(基因组或cdna)或rna(例如,mrna)。可以通过各种方法设计可以被密码子优化以在所选系统(即,细胞类型)中进行表达的此类核酸序列。可以使用在线可用的方法(例如,geneart)、公开的方法或提供密码子优化服务的公司(例如,dna2.0)(加利福尼亚州门洛帕克市)来执行此优化。例如,在国际专利公开第wo2015/012924号中描述了一种密码子优化方法,其通过引用整体并入本文中。还参见例如美国专利公开第2014/0032186号和美国专利公开第2006/0136184号。适合的是,修饰产物的开放阅读框(orf)的全长。然而,在一些实施例中,仅orf的片段可以被改变。通过使用这些方法中的一种方法,可以将频率应用于任何给定的多肽序列,并产生对多肽进行编码的经过密码子优化的编码区的核酸片段。许多选项可用于对密码子进行实际改变或者可用于合成如在此所述地设计的密码子优化的编码区。此类改变或合成可以使用本领域普通技术人员已熟知的标准和常规分子生物学操作来进行。在一种方法中,通过标准方法合成各自长度为80-90个核苷酸且跨越希望的序列的长度的一系列互补寡核苷酸对。这些寡核苷酸对被合成为使得在退火时它们形成80-90个碱基对的双链片段,这些双链片段含有黏性末端,例如在该对中的各寡核苷酸被合成来延伸超过与该对中另一寡核苷酸互补的区域3个、4个、5个、6个、7个、8个、9个、10个或更多个碱基。每对寡核苷酸的单链末端被设计为用另一对寡核苷酸的单链末端退火。允许这些寡核苷酸对退火,并且然后允许约五至六个这些双链片段经由粘性单链末端一起退火,并且随后它们连接在一起并克隆到标准细菌克隆载体中,例如可获自加利福尼亚州卡尔斯巴德英杰公司(invitrogencorporation,carlsbad,calif)的载体。然后通过标准方法对该构建体测序。制备由连接在一起的80到90个碱基对片段的5到6个片段(即约500个碱基对的片段)组成的这些构建体中的若干构建体,以使得整个希望的序列以一系列质粒构建体表示。然后用适当的限制性酶切割这些质粒的插入物并且将其连接在一起以形成最终构建体。然后将最终构建体克隆到标准细菌克隆载体中,并进行测序。另外的方法对于技术人员而言将立即是清楚的。另外,基因合成易于商购获得。在某些实施例中,提供了aav衣壳,所述aav衣壳具有含有多个高度脱酰胺化的“ng”位置的aav衣壳同种型(即,vp1、vp2、vp3)的异质群体。在某些实施例中,高度脱酰胺化的位置在下文参考预测的全长vp1氨基酸序列鉴定的位置中。在其它实施例中,衣壳基因被修饰成使得参考的“ng”被消融,并且突变体“ng”被工程化到另一个位置中。在某些实施例中,混合的raav群体由使用单个aav衣壳核酸序列的生产系统产生,所述核酸序列对一种aav类型的预测的aavvp1氨基酸序列进行编码。然而,产生和制造过程提供了上文所描述的衣壳蛋白的异质群体。在某些实施例中,提供了新颖的分离的aavrh79衣壳。seqidno:1中提供了对aav进行编码的核酸序列,并且seqidno:2中提供了经过编码的氨基酸序列。在某些实施例中,raav包括aavrh79衣壳。aavrh79衣壳包括aavrh79vp1蛋白、aavrh79vp2蛋白和aavrh79vp3蛋白的异质群体。在一个实施例中,aavrh79衣壳是通过从对seqidno:2的1到738的预测的氨基酸序列进行编码的核酸序列的表达产生的。任选地,序列共表达来自不包含vp1独特区(约aa1到137)或vp2独特区(约aa1到203)的核酸序列的vp3蛋白、从seqidno:1产生的vp1蛋白或从与seqidno:1至少70%相同的对seqidno:2的1到738的预测的氨基酸序列进行编码的核酸序列产生的vp1蛋白。在其它实施例中,通过从对seqidno:2的至少约氨基酸138到738的预测的氨基酸序列进行编码的核酸序列的表达产生的aavrh79vp2蛋白、从包括seqidno:1的至少核苷酸412到2214的序列产生的vp2蛋白或从与seqidno:1的至少核苷酸412到2214至少70%相同的对seqidno:2的至少约氨基酸138到738的预测的氨基酸序列进行编码的核酸序列产生的vp2蛋白、通过从对seqidno:2的至少约氨基酸204到738的预测的氨基酸序列进行编码的核酸序列的表达产生的aavrh79vp3蛋白、从包括seqidno:1的至少核苷酸610到2214的序列产生的vp3蛋白或从与seqidno:1的至少核苷酸610到2214至少70%相同的对seqidno:2的至少约氨基酸204到738的预测的氨基酸序列进行编码的核酸序列产生的vp3蛋白。在某些实施例中,aavrh79衣壳包括:vp1蛋白的异质群体,所述vp1蛋白作为对seqidno:2的氨基酸序列进行编码的核酸序列的产物;vp2蛋白的异质群体,所述vp2蛋白作为对seqidno:2的至少约氨基酸138到738的氨基酸序列进行编码的核酸序列的产物;以及vp3蛋白的异质群体,所述vp3蛋白作为对seqidno:2的至少氨基酸204到738进行编码的核酸序列的产物。aavrh79vp1蛋白、vp2蛋白和vp3蛋白含有具有氨基酸修饰的亚群体,所述氨基酸修饰包括seqidno:2中的天冬酰胺-甘氨酸对中的至少两个高度脱酰胺化的天冬酰胺(n),并且任选地进一步包括包含其它脱酰胺化的氨基酸的亚群体,其中脱酰胺化引起氨基酸变化。相对于seqidno:2的编号,在n-g对n57、n263、n385和/或n514处观察到高水平的脱酰胺化。如下表和实例中所示,在其它残基中也观察到脱酰胺化。在某些实施例中,aavrh79可以具有其它脱酰胺化的残基,例如通常小于10%和/或可以具有其它修饰,所述其它修饰包含甲基化(例如,约r487)(通常小于5%,在给定残基处更通常小于1%)、异构化(例如,在d97处)(通常小于5%,在给定残基处更通常小于1%)、磷酸化(例如,在存在的情况下,范围为约10%到约60%、或约10%到约30%、或约20%到约60%)(例如,在s149、约s153、约s474、约t570、约s665中的一个或多个处)或氧化(例如,在w248、w307、w307、m405、m437、m473、w480、w480、w505、m526、m544、m561、w621、m637和/或w697中的一个或多个处)。任选地,w可以氧化成犬尿氨酸。表a-aavrh79脱酰胺化在某些实施例中,在下文提供的如使用胰蛋白酶使用质谱所测定的范围内,在上表中鉴定的位置中的一个或多个位置中修饰aavrh79衣壳。在某些实施例中,如本文所描述的修饰以下位置中的一个或多个位置或n之后的甘氨酸。残基数量基于本文所提供的aavrh79序列。参见seqidno:2。在某些实施例中,seqidno:1中提供了对aavrh79vp1衣壳蛋白进行编码的核酸序列。在其它实施例中,可以选择与seqidno:1具有70%到99.9%同一性的核酸序列以表达aavrh79衣壳蛋白。在某些其它实施例中,核酸序列与seqidno:1至少约75%相同、至少80%相同、至少85%相同、至少90%相同、至少95%相同、至少97%相同或至少99%到99.9%相同。然而,可以选择对seqidno:2的氨基酸序列进行编码的其它核酸序列用于产生raav衣壳。在某些实施例中,核酸序列具有seqidno:1的核酸序列或与seqidno:1至少70%到99%相同、至少75%、至少80%、至少85%、至少90%、至少95%、至少97%、至少99%相同的对seqidno:2进行编码的序列。在某些实施例中,核酸序列具有seqidno:1的核酸序列或与seqidno:1的约nt412到约nt2214至少70%到99%、至少75%、至少80%、至少85%、至少90%、至少95%、至少97%、至少99%相同的对seqidno:2的vp2衣壳蛋白(约aa138到738)进行编码的序列。在某些实施例中,核酸序列具有seqidno:1的约nt610到约nt2214的核酸序列或与ntseqidno:1至少70%到99%、至少75%、至少80%、至少85%、至少90%、至少95%、至少97%、至少99%相同的对seqidno:2的vp3衣壳蛋白(约aa204到738)进行编码的序列。本发明还涵盖对突变体aavrh79进行编码的核酸序列,其中已经改变一个或多个残基以降低脱酰胺化或本文所鉴定的其它修饰。此类核酸序列可以用于产生突变raavrh79衣壳。在某些实施例中,提供了新颖aav8.ar2.08衣壳。seqidno:17中提供了对aav进行编码的核酸序列,并且seqidno:18中提供了经过编码的氨基酸序列。在一个实施例中,重组腺相关病毒(raav)具有aav8.ar2.08衣壳。图13a中提供了aav8t、aav8.ar2.08和aav8的氨基酸序列的比对。图13b-13d中提供了aav8t、aav8.ar2.08和aav8的核酸序列的比对。在某些实施例中,aav8.ar2.08衣壳包括aav8.ar2.08衣壳蛋白,所述aav8.ar2.08衣壳蛋白包括:通过从对seqidno:18的1到738的预测的氨基酸序列进行编码的核酸序列的表达产生的aav8.ar2.08vp1蛋白、从seqidno:17产生的vp1蛋白或从与seqidno:17至少70%相同的对seqidno:18的1到738的预测的氨基酸序列进行编码的核酸序列产生的vp1蛋白、通过从对seqidno:18的至少约氨基酸138到738的预测的氨基酸序列进行编码的核酸序列的表达产生的aav8.ar2.08vp2蛋白、从包括seqidno:17的至少核苷酸412到2214的序列产生的vp2蛋白或从与seqidno:17的至少核苷酸412到2214至少70%相同的对seqidno:18的至少约氨基酸138到738的预测的氨基酸序列进行编码的核酸序列产生的vp2蛋白、通过从对seqidno:18的至少约氨基酸204到738的预测的氨基酸序列进行编码的核酸序列的表达产生的aav8.ar2.08vp3蛋白、从包括seqidno:17的至少核苷酸610到2214的序列产生的vp3蛋白或从与seqidno:17的至少核苷酸610到2214至少70%相同的对seqidno:18的至少约氨基酸204到738的预测的氨基酸序列进行编码的核酸序列产生的vp3蛋白。另外地或可替代地,aav8.ar2.08衣壳包括:vp1蛋白的异质群体,所述vp1蛋白作为对seqidno:18的氨基酸序列进行编码的核酸序列的产物;vp2蛋白的异质群体,所述vp2蛋白作为对seqidno:18的至少约氨基酸138到738的氨基酸序列进行编码的核酸序列的产物;以及vp3蛋白的异质群体,所述vp3蛋白作为对seqidno:18的至少氨基酸204到738进行编码的核酸序列的产物,其中:所述vp1蛋白、所述vp2蛋白和所述vp3蛋白含有具有氨基酸修饰的亚群体,所述氨基酸修饰包括seqidno:18中的天冬酰胺-甘氨酸对中的至少两个高度脱酰胺化的天冬酰胺(n),并且任选地进一步包括包含其它脱酰胺化的氨基酸的亚群体,其中脱酰胺化引起氨基酸变化。aav8.ar2.08由例如在基于aav8.ar2.08vp1[seqidno:18]的编号的位置n57、n263、n385、n514和n540处具有高度脱酰胺化的残基表征。另外地,在本申请的下表和详细表中的位置处的残基示出在aav8.ar2.08衣壳中已经观察到的脱酰胺化。在某些实施例中,在下文提供的如使用胰蛋白酶使用质谱所测定的范围内,在以下位置中的一个或多个位置中修饰aav8.ar2.08衣壳。在某些实施例中,如本文所描述的修饰以下位置中的一个或多个位置或n之后的甘氨酸。例如,在某些实施例中,g可以例如在位置58、264、386、515或541处被修饰为s或a。当ng57/58改变为ns57/58或na57/58时,观察到脱酰胺化显著降低。然而,在某些实施例中,当ng改变为ns或na时,观察到脱酰胺化增加。在某些实施例中,ng对的n被修饰为q,同时保留g。在某些实施例中,ng对的两个氨基酸均被修饰。在某些实施例中,n385q引起所述位置中的脱酰胺化显著降低。在某些实施例中,n499q引起所述位置中的脱酰胺化显著增加。除了脱酰胺化之外,其它修饰可以包含异构化(例如,在d442和/或d584中的一个或多个处)(1-15%)、磷酸化(例如,在约s149、约t417、约t454、约t493、s600和/或约t663中的一个或多个处)和/或氧化(例如,在约w22、约m204、约m212、w248、w307、m405、m437、m473、w480、w505、m526、m561、m607、约w609、w621、m637、w697中的一个或多个处)。其它位置可以具有此类这些或其它修饰(例如,乙酰化或另外的脱酰胺化)。表b-aav8.ar2.08脱酰胺化在某些实施例中,seqidno:17中提供了对aav8.ar2.08vp1衣壳蛋白进行编码的核酸序列。在其它实施例中,可以选择与seqidno:17具有70%到99.9%同一性的核酸序列以表达aav8.ar2.08衣壳蛋白。在某些其它实施例中,核酸序列与seqidno:17至少约75%相同、至少80%相同、至少85%相同、至少90%相同、至少95%相同、至少97%相同或至少99%到99.9%相同。然而,可以选择对seqidno:18的氨基酸序列进行编码的其它核酸序列用于产生raav8.ar2.08衣壳。在某些实施例中,核酸序列具有seqidno:17的核酸序列或与seqidno:17至少70%到99%相同、至少75%、至少80%、至少85%、至少90%、至少95%、至少97%、至少99%相同的对seqidno:18进行编码的序列。在某些实施例中,核酸序列具有seqidno:17的核酸序列或与seqidno:17的约nt412到约nt2214至少70%到99%、至少75%、至少80%、至少85%、至少90%、至少95%、至少97%、至少99%相同的对seqidno:18的vp2衣壳蛋白(约aa138到738)进行编码的序列。在某些实施例中,核酸序列具有seqidno:17的约nt607到约nt2214的核酸序列或与ntseqidno:17至少70%到99%、至少75%、至少80%、至少85%、至少90%、至少95%、至少97%、至少99%相同的对seqidno:18的vp3衣壳蛋白(约aa204到738)进行编码的序列。本发明还涵盖对突变体aav8.2.08进行编码的核酸序列,其中已经改变一个或多个残基以降低脱酰胺化或本文所鉴定的其它修饰。此类核酸序列可以用于产生突变体raav8.2.08。在某些实施例中,提供了新颖aav5.5.9衣壳。seqidno:9中提供了对aav进行编码的核酸序列,并且seqidno:10中提供了经过编码的氨基酸序列。图12a中示出了aav5.5.9、aav9和aavphp.b的氨基酸序列的比对。图12b-12e中示出了aav5.5.9、aav9和aavphp.b的核酸序列的比对。在一个实施例中,重组腺相关病毒(raav)具有aav5.5.9衣壳,所述aav5.5.9衣壳包括:aav5.5.9衣壳蛋白,所述aav5.5.9衣壳蛋白包括:以下的异质群体:通过从对seqidno:10的1到726的预测的氨基酸序列进行编码的核酸序列的表达产生的aav5.5.9vp1蛋白、从seqidno:9产生的vp1蛋白或从与seqidno:9至少70%相同的对seqidno:1的1到726的预测的氨基酸序列进行编码的核酸序列产生的vp1蛋白;以下的异质群体:通过从对seqidno:10的至少约氨基酸137到726的预测的氨基酸序列进行编码的核酸序列的表达产生的aav5.5.9vp2蛋白、从包括seqidno:9的至少核苷酸409到2178的序列产生的vp2蛋白或从与seqidno:9的至少核苷酸577到2178至少70%相同的对seqidno:10的至少约氨基酸137到726的预测的氨基酸序列进行编码的核酸序列产生的vp2蛋白;以及以下的异质群体:通过从对seqidno:10的至少约氨基酸193到726的预测的氨基酸序列进行编码的核酸序列的表达产生的aav5.5.9vp3蛋白、从包括seqidno:9的至少核苷酸577到2178的序列产生的vp3蛋白或从与seqidno:9的至少核苷酸577到2178至少70%相同的对seqidno:10的至少约氨基酸193到726的预测的氨基酸序列进行编码的核酸序列产生的vp3蛋白。另外地或可替代地,aav5.5.9衣壳包括:vp1蛋白的异质群体,所述vp1蛋白作为对seqidno:10的氨基酸序列进行编码的核酸序列的产物;vp2蛋白的异质群体,所述vp2蛋白作为对seqidno:10的至少约氨基酸137到726的氨基酸序列进行编码的核酸序列的产物;以及vp3蛋白的异质群体,所述vp3蛋白作为对seqidno:10的至少氨基酸193到726进行编码的核酸序列的产物,其中:所述vp1蛋白、所述vp2蛋白和所述vp3蛋白含有具有氨基酸修饰的亚群体,所述氨基酸修饰包括seqidno:10中的天冬酰胺-甘氨酸对中的至少两个高度脱酰胺化的天冬酰胺(n),并且任选地进一步包括包含其它脱酰胺化的氨基酸的亚群体,其中脱酰胺化引起氨基酸变化。表c-aav5.5.9脱酰胺化在某些实施例中,seqidno:9中提供了对aav5.5.9vp1衣壳蛋白进行编码的核酸序列。在其它实施例中,可以选择与seqidno:9具有70%到99.9%同一性的核酸序列以表达aav5.5.9衣壳蛋白。在某些其它实施例中,核酸序列与seqidno:9至少约75%相同、至少80%相同、至少85%相同、至少90%相同、至少95%相同、至少97%相同或至少99%到99.9%相同。然而,可以选择对seqidno:10的氨基酸序列进行编码的其它核酸序列用于产生raav5.5.9衣壳。在某些实施例中,核酸序列具有seqidno:10的核酸序列或与seqidno:9至少70%到99%相同、至少75%、至少80%、至少85%、至少90%、至少95%、至少97%、至少99%相同的对seqidno:10进行编码的序列。在某些实施例中,核酸序列具有seqidno:9的核酸序列或与seqidno:9的约nt409到约nt2178至少70%到99%、至少75%、至少80%、至少85%、至少90%、至少95%、至少97%、至少99%相同的对seqidno:10的vp2衣壳蛋白(约aa137到726)进行编码的序列。在某些实施例中,核酸序列具有seqidno:9的约nt577到约nt2178的核酸序列或与seqidno:9的nt577到2178至少70%到99%、至少75%、至少80%、至少85%、至少90%、至少95%、至少97%、至少99%相同的对seqidno:10的vp3衣壳蛋白(约aa193到726)进行编码的序列。本发明还涵盖对突变体aav5.5.9进行编码的核酸序列,其中已经改变一个或多个残基以降低脱酰胺化或本文所鉴定的其它修饰。此类核酸序列可以用于产生突变体raav5.5.9。i.raav载体如上文所指示的,新颖aav序列和蛋白可用于产生raav,并且还可用于重组aav载体,所述重组aav载体可以是反义递送载体、基因疗法载体或疫苗载体。另外,本文描述的经过工程化的aav衣壳可以用于工程化raav载体,以将多种合适的核酸分子递送到靶细胞和组织。包装到aav衣壳中并递送到宿主细胞的基因组序列通常至少由转基因及其调节序列和aav反向末端重复序列(itr)构成。单链aav和自身互补(sc)aav两者都涵盖在raav内。转基因是与载体序列异质的核酸编码序列,所述核酸编码序列对多肽、蛋白质、功能性rna分子(例如,mirna、mirna抑制剂)或其它所关注的基因产物进行编码。核酸编码序列以允许转基因在靶组织的细胞中转录、转译和/或表达的方式与调节组分可操作地连接。载体的aav序列通常包括顺式作用的5'和3'反向末端重复序列(参见例如b.j.carter,《细小病毒手册(handbookofparvoviruses)》,p.tijsser编辑,crc出版社,第155到168页(1990))。itr序列的长度为约145bp。优选地,分子中使用了对itr进行编码的基本上整个序列,尽管允许对这些序列进行一定程度的微小修饰。修饰这些itr序列的能力在本领域的技术范围内。(参见例如文本,如sambrook等人,《分子克隆:实验室手册(molecularcloning.alaboratorymanual)》,第2版,冷泉港实验室(coldspringharborlaboratory),纽约(1989);以及k.fisher等人,《病毒学杂志》,70:520532(1996))。在本发明中采用的这种分子的实例是含有转基因的“顺式作用”质粒,其中所选转基因序列和相关的调节元件侧接5'和3'aavitr序列。在一个实施例中,itr来自与供应衣壳的aav不同的aav,从而产生假型载体。在一个实施例中,itr来自aav2。已经描述了被称为δitr的5'itr的缩短版本,其中缺失了d序列和末端解析位点(trs)。在其它实施例中,使用了全长aav5'和3'itr。然而,可以选择来自其它aav来源的itr。在itr的来源来自aav2并且aav衣壳来自另一个aav来源的情况下,所得载体可以被称为假型的。然而,这些元件的其它构型可以是合适的。除重组aav载体的上文鉴定的主要元件外,aav载体还包含必需的常规控制元件,所述常规控制元件以允许其在用质粒载体转染或用由本发明产生的病毒感染的细胞中转录、转译和/或表达的方式与转基因可操作地连接。如本文所使用的,“可操作地连接的”序列包含与所关注的基因邻接的表达控制序列和以反式或在远处起作用以控制所关注的基因的表达控制序列。调节控制元件通常含有作为表达控制序列的一部分的启动子序列,例如定位在所选5'itr序列与编码序列之间。可以在本文所描述的载体中使用组成型启动子、可调节启动子[参见例如wo2011/126808和wo2013/04943]、组织特异性启动子或对生理学线索有应答的启动子。一种或多种启动子可以选自不同的来源,例如人巨细胞病毒(cmv)立即早期增强子/启动子、sv40早期增强子/启动子、jc多瘤病毒启动子、髓鞘碱性蛋白(mbp)或神经胶质原纤维酸性蛋白(gfap)启动子、单纯疱疹病毒(hsv-1)潜伏期相关启动子(lap)、劳氏肉瘤病毒(rsv)长末端重复(ltr)启动子、神经元特异性启动子(nse)、血小板衍生生长因子(pdgf)启动子、hsyn、黑色素浓缩激素(mch)启动子、cba、基质金属蛋白启动子(mpp)和鸡β-肌动蛋白启动子。在一个实施例中,启动子是肝脏特异性启动子,如本文例示的被称为lsp的启动子。除了启动子之外,载体还可以含有一个或多个其它合适的转录起始、终止、增强子序列、有效的rna加工信号,如剪接和聚腺苷酸化(polya)信号;稳定胞质mrna的序列,例如wpre;增强转译效率的序列(即,kozak共有序列);增强蛋白稳定性的序列;以及在需要时,增强所编码产物的分泌的序列。合适的增强子的实例是cmv增强子。其它合适的增强子包含适合于所期望的靶组织适应症的增强子。在一个实施例中,表达盒包括一种或多种表达增强子。在一个实施例中,表达盒含有两种或更多种表达增强子。这些增强子可以相同或彼此不同。例如,增强子可以包含cmv立即早期增强子。这种增强子可以存在于彼此相邻定位的两个拷贝中。可替代地,增强子的双拷贝可以被一个或多个序列分开。在仍另一个实施例中,表达盒进一步含有内含子,例如,鸡β-肌动蛋白内含子。其它合适的内含子包含本领域已知的内含子,例如,如wo2011/126808中描述的内含子。合适的polya序列的实例包含例如sv40、sv50、牛生长激素(bgh)、人生长激素和合成polya。任选地,可以选择一个或多个序列来稳定mrna。这种序列的实例是经过修饰的wpre序列,其可以在polya序列的上游和编码序列的下游被工程化[参见例如mazanta-boussif等人,《基因疗法(genetherapy)》(2009)16:605-619]。这些raav特别适合于用于治疗目的和用于免疫的基因递送,包含诱导保护性免疫。进一步地,本发明的组合物还可以用于体外产生所期望的基因产物。对于体外产生,可以在用含有对所期望的产物进行编码的分子的raav转染宿主细胞并在允许表达的条件下培养细胞培养物之后从所期望的培养物中获得所期望的产物(例如,蛋白质)。然后可以根据需要纯化和分离所表达的产物。用于转染、细胞培养、纯化和分离的合适的技术是本领域的技术人员已知的。治疗性转基因由转基因编码的有用产物包含各种基因产物,所述基因产物替代缺陷型或有缺陷的基因,使失活或“敲除”、或“敲低”或减少以不期望的高水平表达或递送具有所期望的治疗效果的基因产物的基因的表达。在大多数实施例中,疗法将是“体细胞基因疗法”,即,将基因转移到不产生精子或卵子的人体细胞。在某些实施例中,转基因表达蛋白具有天然人序列的序列。然而,在其它实施例中,表达合成蛋白。此类蛋白质可以用于治疗人,或者在其它实施例中,被设计成用于治疗动物,包含如犬科或猫科群体等伴侣动物,或用于治疗与人群体接触的牲畜或其它动物。合适的基因产物的实例可以包含与家族性高胆固醇血症、肌营养不良、囊性纤维化以及罕见疾或孤儿病相关的基因产物。此类罕见疾病的实例可以包含脊髓性肌萎缩症(sma)、亨廷顿氏病(huntingdon'sdisease)、雷特综合征(rettsyndrome)(例如,甲基cpg结合蛋白2(mecp2);uniprotkb-p51608)、肌萎缩侧索硬化症(als)、杜兴型肌营养不良(duchennetypemusculardystrophy)、弗里德里希共济失调(friedrichsataxia)(例如,共济蛋白)、颗粒蛋白前体(prgn)(与非阿尔茨海默氏病的大脑变性相关,包含额颞叶痴呆(ftd)、进行性非流利性失语症(pnfa)和语义性痴呆)等等。参见例如www.orpha.net/consor/cgi-bin/disease_search_list.php;rarediseases.info.nih.gov/diseases。合适的基因的实例可以包含例如激素以及生长和分化因子,包含但不限于胰岛素、胰高血糖素、胰高血糖素样肽-1(glp1)、生长激素(gh)、甲状旁腺激素(pth)、生长激素释放因子(grf)、促卵泡激素(fsh)、促黄体激素(lh)、人绒毛膜促性腺激素(hcg)、血管内皮生长因子(vegf)、血管生成素、血管抑素、粒细胞集落刺激因子(gcsf)、促红细胞产生素(epo)(包含例如人、犬或猫epo)、结缔组织生长因子(ctgf)、神经营养因子包含例如碱性成纤维细胞生长因子(bfgf)、酸性成纤维细胞生长因子(afgf)、表皮生长因子(egf)、血小板衍生生长因子(pdgf)、胰岛素生长因子i和ii(igf-i和igf-ii)、转化生长因子α超家族(包含tgfα、激活素、抑制素)中的任一种、或骨形态发生蛋白(bmp)bmp1-15中的任一种、生长因子的调蛋白/神经调节蛋白/aria/neu分化因子(ndf)家族、神经生长因子(ngf)、脑源性神经营养因子(bdnf)、神经营养因子nt-3和nt-4/5、睫状神经营养因子(cntf)、神经胶质细胞系源性神经营养因子(gdnf)、神经秩蛋白、集聚蛋白中的任一种、信号素/脑衰蛋白、纺锤蛋白-1和纺锤蛋白-2、肝细胞生长因子(hgf)、肝配蛋白、头蛋白、音猬因子和酪氨酸羟化酶的家族中的任一种。其它有用的转基因产物包含调节免疫系统的蛋白质,包含但不限于细胞因子和淋巴因子,如血小板生成素(tpo)、白细胞介素(il)il-1到il-36(包含例如人白细胞介素il-1、il-1α、il-1β、il-2、il-3、il-4、il-6、il-8、il-12、il-11、il-12、il-13、il-18、il-31、il-35)、单核细胞趋化蛋白、白血病抑制因子、粒细胞-巨噬细胞集落刺激因子、fas配体、肿瘤坏死因子α和β、干扰素α、β和γ、干细胞因子、flk-2/flt3配体。由免疫系统产生的基因产物也可用于本发明。这些包含但不限于免疫球蛋白igg、igm、iga、igd和ige、嵌合免疫球蛋白、人源化抗体、单链抗体、t细胞受体、嵌合t细胞受体、单链t细胞受体、mhci类和ii类分子以及经过工程化的免疫球蛋白和mhc分子。例如,在某些实施例中,可以将raav抗体设计成递送犬或猫抗体,例如抗ige、抗il31、抗cd20、抗ngf、抗gnrh。有用的基因产物还包含补体调节蛋白,如补体调节蛋白、膜辅因子蛋白(mcp)、衰变加速因子(daf)、cr1、cf2、cd59和c1酯酶抑制剂(c1-inh)。仍其它有用的基因产物包含用于激素、生长因子、细胞因子、淋巴因子、调节蛋白和免疫系统蛋白的受体中的任一种。本发明涵盖用于胆固醇调节和/或脂质调节的受体,包含低密度脂蛋白(ldl)受体、高密度脂蛋白(hdl)受体、极低密度脂蛋白(vldl)受体和清除剂受体。本发明还涵盖基因产物,如类固醇激素受体超家族的成员,包含糖皮质激素受体和雌激素受体、维生素d受体和其它核受体。另外,有用的基因产物包含转录因子,如jun、fos、max、mad、血清响应因子(srf)、ap-1、ap2、myb、myod和肌生成素、含ets盒的蛋白质、tfe3、e2f、atf1、atf2、atf3、atf4、zf5、nfat、creb、hnf-4、c/ebp、sp1、ccaat盒结合蛋白、干扰素调节因子(irf-1)、威尔姆斯肿瘤蛋白、ets结合蛋白、stat、gata盒结合蛋白(例如,gata-3)和带翼螺旋蛋白的叉头家族。其它有用的基因产物包含氨基甲酰合成酶i、鸟氨酸转氨甲酰酶(otc)、精氨酸琥珀酸合成酶、用于治疗精氨琥珀酸裂解酶缺乏症的精氨琥珀酸裂解酶(asl)、精氨酸酶、延胡索酰乙酰乙酸水解酶、苯丙氨酸羟化酶、α-1抗胰蛋白酶、恒河猴甲胎蛋白(afp)、恒河猴绒毛膜促性腺激素(cg)、葡萄糖-6-磷酸酶、胆色素原脱氨酶、胱硫醚β合酶、支链酮酸脱羧酶、白蛋白、异戊酰辅酶a脱氢酶、丙酰辅酶a羧化酶、甲基丙二酰辅酶a变位酶、戊二酰辅酶a脱氢酶、胰岛素、β-葡糖苷酶、丙酮酸羧酸盐、肝磷酸化酶、磷酸化酶激酶、甘氨酸脱羧酶、h蛋白、t蛋白、囊性纤维化跨膜调节子(cftr)序列和肌营养不良蛋白基因产物[例如,迷你或微小肌营养不良蛋白]。仍其它有用的基因产物还包含如可以用于酶替代疗法的酶,所述酶替代疗法可用于由于酶活性不足而导致的多种病状。例如,可以将含有甘露糖-6-磷酸的酶用于溶酶体贮积病的疗法中(例如,合适的基因包含对β-葡糖醛酸酶(gusb)进行编码的基因)。在某些实施例中,可以在基因编辑系统中使用raav,所述系统可以涉及一种raav或多种raav原液的共同施用。例如,raav可以被工程化以递送spcas9、sacas9、arcus、cpf1和其它合适的基因编辑构建体。仍其它有用的基因产物包含用于治疗血友病的基因产物,所述血友病包含血友病b(包含因子ix)和血友病a(包含因子viii及其变体,如异二聚体和b缺失结构域的轻链和重链;美国专利第6,200,560号和美国专利第6,221,349号)。在一些实施例中,迷你基因包括因子viii重链的前57个碱基对,所述重链对10个氨基酸信号序列以及人生长激素(hgh)聚腺苷酸化序列进行编码。在替代性实施例中,迷你基因进一步包括a1和a2结构域以及来自b结构域的n端的5个氨基酸和/或b结构域的c端的85个氨基酸以及a3、c1和c2结构域。在又其它实施例中,在单个迷你基因中提供了对因子viii重链和轻链进行编码的核酸,所述单个迷你基因由对b结构域的14个氨基酸进行编码的42个核酸分开[美国专利第6,200,560号]。其它有用的基因产物包含非天然存在的多肽,如具有含有插入、缺失或氨基酸取代的非天然存在的氨基酸序列的嵌合或杂合多肽。例如,单链工程化的免疫球蛋白可能在某些免疫受损的患者中有用。其它类型的非天然存在的基因序列包含反义分子和催化核酸,如核酶,其可以用于减少靶标的过度表达。减少和/或调节基因表达对于治疗由细胞过度增殖表征的过度增殖性病状(如癌症和牛皮癣)是特别期望的。靶多肽包含与正常细胞相比在过度增殖性细胞中专门产生或以更高水平产生的那些多肽。靶抗原包含由癌基因如myb、myc、fyn和易位基因bcr/abl、ras、src、p53、neu、trk和egrf编码的多肽。除作为靶抗原的癌基因产物之外,用于抗癌治疗和保护方案的靶多肽包含由b细胞淋巴瘤产生的抗体的可变区和t细胞淋巴瘤的t细胞受体的可变区,在一些实施例中,所述可变区还被用作自身免疫疾病的靶抗原。其它肿瘤相关多肽也可以用作靶多肽,如在肿瘤细胞中以较高水平存在的多肽,包含由单克隆抗体17-1a识别的多肽和叶酸结合多肽。其它合适的治疗性多肽和蛋白质包含可以用于通过为针对与自身免疫相关的靶标赋予广泛基础的保护性免疫应答而治疗患有自身免疫性疾病和病症的个体的多肽和蛋白质,所述靶标包含细胞受体和产生“自身”定向抗体的细胞。t细胞介导的自身免疫性疾病包含类风湿关节炎(ra)、多发性硬化症(ms)、干燥综合征(syndrome)、结节病、胰岛素依赖型糖尿病(iddm)、自身免疫性甲状腺炎、反应性关节炎、强直性脊柱炎、硬皮病、多发性肌炎、皮肌炎、牛皮癣、韦格纳氏肉芽肿病(wegner'sgranulomatosis)、克罗恩氏病(crohn'sdisease)和溃疡性结肠炎。这些疾病中的每种疾病由与内源性抗原结合并引发与自身免疫性疾病相关的炎性级联的t细胞受体(tcr)表征。可以通过raav递送的另外的说明性基因包含但不限于与糖原贮积病或1a型缺乏症(gsd1)相关的葡萄糖-6-磷酸酶;与pepck缺乏症相关的磷酸烯醇丙酮酸羧激酶(pepck);细胞周期蛋白依赖性激酶样5(cdkl5),其也被称为与癫痫发作和严重的神经发育障碍相关的丝氨酸/苏氨酸激酶9(stk9);与半乳糖血症相关的半乳糖-1磷酸尿嘧啶转移酶;与苯丙酮尿症(pku)相关的苯丙氨酸羟化酶;与枫糖尿病相关的支链α-酮酸脱氢酶;与1型酪氨酸血症相关的延胡索酰乙酰乙酸水解酶;与甲基丙二酸血症相关的甲基丙二酰辅酶a变位酶;与中链乙酰辅酶a缺乏症相关的中链酰基辅酶a脱氢酶;与鸟氨酸转氨甲酰酶缺乏症相关的鸟氨酸转氨甲酰酶(otc);与瓜氨酸血症相关的精氨酸琥珀酸合成酶(ass1);卵磷脂胆固醇酰基转移酶(lcat)缺乏症;甲基丙二酸血症(mma);尼曼-皮克病(niemann-pickdisease),c1型;丙酸血症(pa);与家族性高胆固醇血症(fh)相关的低密度脂蛋白受体(ldlr)蛋白;与克里格勒-纳贾尔病(crigler-najjardisease)相关的udp-葡萄糖醛糖基转移酶;与严重联合免疫缺陷病相关的腺苷脱氨酶;与痛风和莱施-奈恩综合征(lesch-nyhansyndrome)相关的次黄嘌呤鸟嘌呤磷酸核糖基转移酶;与生物素酶缺乏症相关的生物素酶;与法布里病(fabrydisease)相关的α-半乳糖苷酶a(α-gala);与威尔逊氏病(wilson'sdisease)相关的atp7b;与戈谢病(gaucherdisease)2和3型相关的β-葡糖脑苷脂酶;与泽尔韦格氏综合征(zellwegersyndrome)相关的过氧化物酶体膜蛋白70kda;与变质性脑白质营养不良相关的芳基硫酸酯酶a(arsa);与克拉伯病(krabbedisease)相关的半乳糖脑苷脂酶(galc);与庞贝病(pompedisease)相关的α-葡糖苷酶(gaa);与尼曼-皮克病(niemanpickdisease)a型相关的鞘磷脂酶(smpd1)基因;与成人ii型瓜氨酸血症(ctln2)相关的精氨琥珀酸合酶;与脲循环病症相关的氨基甲酰磷酸合酶1(cps1);与脊髓性肌萎缩症相关的存活运动神经元(smn)蛋白;与法伯脂肪肉芽肿病相关的神经酰胺酶;与gm2神经节苷脂病和泰伊-萨克斯二氏病和山霍夫氏病(tay-sachsandsandhoffdiseases)相关的b-己糖胺酶;与天冬氨酰葡糖尿症相关的天冬氨酰葡糖胺酶;与岩藻糖苷贮积症相关的α岩藻糖苷酶;与α甘露糖苷贮积症相关的α-甘露糖苷酶;与急性间歇性卟啉症(aip)相关的胆色素原脱氨酶;用于治疗α-1抗胰蛋白酶缺乏症(肺气肿)的α-1抗胰蛋白酶;用于治疗因地中海贫血或肾衰竭引起的贫血的促红细胞生成素;用于治疗缺血性疾病的血管内皮生长因子、血管生成素-1和成纤维细胞生长因子;用于治疗如例如在动脉粥样硬化、血栓形成或栓塞中所看见的阻塞的血管的血栓调节蛋白和组织因子途径抑制剂;用于治疗帕金森氏病(parkinson'sdisease)的芳香族氨基酸脱羧酶(aadc)和酪氨酸羟化酶(th);与受磷蛋白、肌浆(内质)网腺苷三磷酸酶2(serca2)呈反义或为其突变体形式的β肾上腺素能受体;用于治疗充血性心力衰竭的心脏腺苷酸环化酶;用于治疗各种癌症的肿瘤抑制基因,如p53;用于治疗炎症和免疫病症以及癌症的细胞因子,如各种白细胞介素之一;用于治疗肌营养不良的肌营养不良蛋白或迷你肌营养不良蛋白以及肌萎缩相关蛋白或迷你肌萎缩相关蛋白;以及用于治疗糖尿病的胰岛素或glp-1。另外所关注的基因和疾病包含例如肌张力异常蛋白基因相关的疾病,如遗传性感觉和自主神经病vi型(dst基因对肌张力异常蛋白进行编码;由于蛋白质的大小(约7570aa)可能需要双重aav载体;scn9a相关疾病,其中功能突变体的丧失导致无法感觉疼痛,并且功能突变体的获得引起疼痛病状,如红斑性肢痛症。由于nefl基因(神经丝轻链)发生突变,另一种病状是腓骨肌萎缩症1f和2e型,其由具有可变临床和电生理表达的进行性周围运动和感觉神经病表征。在某些实施例中,本文所描述的raav可以用于治疗黏多糖贮积症(mps)病症。这种raav可以含有携带对用于治疗mpsi(贺勒、贺勒-施艾氏和施艾氏综合征(hurler,hurler-scheieandscheiesyndromes))的α-l-艾杜糖苷酸酶(idua)进行编码的核酸序列;对用于治疗mpsii(亨特氏综合征)的艾杜糖醛酸-2-硫酸酯酶(ids)进行编码的核酸序列;对用于治疗mpsiiia、b、c和d(沙费利波综合征(sanfilipposyndrome))的磺酰胺酶(sgsh)进行编码的核酸序列;对用于治疗mpsiva和b(莫基奥综合征(morquiosyndrome))的n-乙酰半乳糖胺-6-硫酸硫酸酯酶(galns)进行编码的核酸序列;对用于治疗mpsvi(马罗托-拉米氏综合征(maroteaux-lamysyndrome))的芳基硫酸酯酶b(arsb)进行编码的核酸序列;对用于治疗mpsiix(透明质酸酶缺乏症)的透明质酸酶进行编码的核酸序列;以及对用于治疗mpsvii(斯赖综合征(slysyndrome))的β-葡糖醛酸苷酶进行编码的核酸序列。免疫原性转基因在一些实施例中,通过向患有癌症的受试者施用含有raav载体的raav,包括对与癌症相关的基因产物(例如,肿瘤抑制因子)进行编码的核酸的raav载体可以用于治疗癌症。在一些实施例中,通过向患有癌症的受试者施用含有raav载体的raav,包括对抑制与癌症相关的基因产物(例如,致癌基因)的表达的小干扰核酸(例如,shrna、mirna)进行编码的核酸的raav载体可以用于治疗癌症。在一些实施例中,包括对与癌症相关的基因产物(或抑制与癌症相关的基因表达的功能性rna)进行编码的核酸的raav载体可以用于研究目的,例如研究癌症或鉴定治疗癌症的治疗剂。以下是已知与癌症的发展相关的示例性基因(例如,癌基因和肿瘤抑制因子)的非限制性列表:aars、abcb1、abcc4、abi2、abl1、abl2、ack1、acp2、acy1、adsl、ak1、akr1c2、akt1、alb、anpep、anxa5、anxa7、ap2m1、apc、arhgap5、arhgef5、arid4a、asns、atf4、atm、atp5b、atp5o、axl、bard1、bax、bcl2、bhlhb2、blmh、braf、brca1、brca2、btk、canx、cap1、capn1、capns1、cav1、cbfb、cblb、ccl2、ccnd1、ccnd2、ccnd3、ccne1、cct5、ccyr61、cd24、cd44、cd59、cdc20、cdc25、cdc25a、cdc25b、cdc2l5、cdk10、cdk4、cdk5、cdk9、cdkl1、cdkn1a、cdkn1b、cdkn1c、cdkn2a、cdkn2b、cdkn2d、cebpg、cenpc1、cgrrf1、chaf1a、cib1、ckmt1、clk1、clk2、clk3、clns1a、cltc、col1a1、col6a3、cox6c、cox7a2、crat、crhr1、csf1r、csk、csnk1g2、ctnna1、ctnnb1、ctps、ctsc、ctsd、cul1、cyr61、dcc、dcn、ddx10、dek、dhcr7、dhrs2、dhx8、dlg3、dvl1、dvl3、e2f1、e2f3、e2f5、egfr、egr1、eif5、epha2、erbb2、erbb3、erbb4、ercc3、etv1、etv3、etv6、f2r、fastk、fbn1、fbn2、fes、fgfr1、fgr、fkbp8、fn1、fos、fosl1、fosl2、foxg1a、foxo1a、frap1、frzb、ftl、fzd2、fzd5、fzd9、g22p1、gas6、gcn5l2、gdf15、gna13、gnas、gnb2、gnb2l1、gpr39、grb2、gsk3a、gspt1、gtf2i、hdac1、hdgf、hmmr、hprt1、hrb、hspa4、hspa5、hspa8、hspb1、hsph1、hyal1、hyou1、icam1、id1、id2、idua、ier3、ifitm1、igf1r、igf2r、igfbp3、igfbp4、igfbp5、il1b、ilk、ing1、irf3、itga3、itga6、itgb4、jak1、jarid1a、jun、junb、jund、k-α-1、kit、kitlg、klk10、kpna2、kras2、krt18、krt2a、krt9、lamb1、lamp2、lck、lcn2、lep、litaf、lrpap1、ltf、lyn、lztr1、madh1、map2k2、map3k8、mapk12、mapk13、mapkapk3、mapre1、mars、mas1、mcc、mcm2、mcm4、mdm2、mdm4、met、mgst1、micb、mllt3、mme、mmp1、mmp14、mmp17、mmp2、mnda、msh2、msh6、mt3、myb、mybl1、mybl2、myc、mycl1、mycn、myd88、myl9、mylk、neo1、nf1、nf2、nfkb1、nfkb2、nfsf7、nid、nine、nmbr、nme1、nme2、nme3、notch1、notch2、notch4、npm1、nqo1、nr1d1、nr2f1、nr2f6、nras、nrg1、nsep1、osm、pa2g4、pabpc1、pcna、pctk1、pctk2、pctk3、pdgfa、pdgfb、pdgfra、pdpk1、pea15、pfdn4、pfdn5、pgam1、phb、pik3ca、pik3cb、pik3cg、pim1、pkm2、pkmyt1、plk2、ppard、pparg、ppih、ppp1ca、ppp2r5a、prdx2、prdx4、prkar1a、prkcbp1、prnp、prss15、psma1、ptch、pten、ptgs1、ptma、ptn、ptprn、rab5a、rac1、rad50、raf1、ralbp1、rap1a、rara、rarb、rasgrf1、rb1、rbbp4、rbl2、rea、rel、rela、relb、ret、rfc2、rgs19、rhoa、rhob、rhoc、rhod、ripk1、rpn2、rps6kb1、rrm1、sars、selenbp1、sema3c、sema4d、sepp1、serpinh1、sfn、sfpq、sfrs7、shb、shh、siah2、siva、sivatp53、ski、skil、slc16a1、slc1a4、slc20a1、smo、鞘磷脂磷酸二酯酶1(smpd1)、snai2、snd1、snrpb2、socs1、socs3、sod1、sort1、spint2、spry2、src、srpx、stat1、stat2、stat3、stat5b、stc1、taf1、tbl3、tbrg4、tcf1、tcf7l2、tfap2c、tfdp1、tfdp2、tgfa、tgfb1、tgfbi、tgfbr2、tgfbr3、thbs1、tie、timp1、timp3、tjp1、tk1、tle1、tnf、tnfrsf10a、tnfrsf10b、tnfrsf1a、tnfrsf1b、tnfrsf6、tnfsf7、tnk1、tob1、tp53、tp53bp2、tp5313、tp73、tpbg、tpt1、tradd、tram1、trrap、tsg101、tufm、txnrd1、tyro3、ubc、ube2l6、uchl1、usp7、vdac1、vegf、vhl、vil2、wee1、wnt1、wnt2、wnt2b、wnt3、wnt5a、wt1、xrcc1、yes1、ywhab、ywhaz、zap70和znf9。raav载体可以包括对调节细胞凋亡的蛋白质或功能性rna进行编码的作为转基因的核酸。以下是与细胞凋亡相关的基因的非限制性列表,并且对这些基因的产物及其同源物进行编码的以及对抑制这些基因及其同源物的表达的小干扰核酸(例如,shrna、mirna)进行编码的核酸在本发明的某些实施例中用作转基因:rps27a、abl1、akt1、apaf1、bad、bag1、bag3、bag4、bak1、bax、bcl10、bcl2、bcl2a1、bcl2l1、bcl2l10、bcl2l11、bcl2l12、bcl2l13、bcl2l2、bclaf1、bfar、bid、bik、naip、birc2、birc3、xiap、birc5、birc6、birc7、birc8、bnip1、bnip2、bnip3、bnip3l、bok、braf、card10、card11、nlrc4、card14、nod2、nod1、card6、cards、cards、casp1、casp10、casp14、casp2、casp3、casp4、casp5、casp6、casp7、casp8、casp9、cflar、cidea、cideb、cradd、dapk1、dapk2、dffa、dffb、fadd、gadd45a、gdnf、hrk、igf1r、lta、ltbr、mcl1、nol3、pycard、ripk1、ripk2、tnf、tnfrsf10a、tnfrsf10b、tnfrsf10c、tnfrsf10d、tnfrsf11b、tnfrsf12a、tnfrsf14、tnfrsf19、tnfrsf1a、tnfrsf1b、tnfrsf21、tnfrsf25、cd40、fas、tnfrsf6b、cd27、tnfrsf9、tnfsf10、tnfsf14、tnfsf18、cd40lg、faslg、cd70、tnfsf8、tnfsf9、tp53、tp53bp2、tp73、tp63、tradd、traf1、traf2、traf3、traf4和traf5。有用的转基因产物还包含mirna。mirna和其它小干扰核酸通过靶信使rna(mrna)的靶rna转录物裂解/降解或转译抑制来调节基因表达。mirna是天然表达的,通常作为最终的19-25种非转译的rna产物。mirna通过与靶mrna的3'非转译区(utr)的序列特异性相互作用展现出其活性。这些内源表达的mirna形成发夹前体,所述发夹前体随后被加工成mirna双链体,并且被进一步加工成“成熟的”单链mirna分子。这种成熟的mirna引导多蛋白复合物mirisc,所述多蛋白复合物基于与成熟的mirna的互补性来鉴定靶mrna的靶位点,例如在3'utr区中。在方法的某些实施例中,以下mirna基因及其同源物的非限制性列表可用作转基因或由转基因编码的小干扰核酸(例如,mirna海绵体、反义寡核苷酸、tudrna)的靶标:hsa-let-7a、hsa-let-7a*、hsa-let-7b、hsa-let-7b*、hsa-let-7c、hsa-let-7c*、hsa-let-7d、hsa-let-7d*、hsa-let-7e、hsa-let-7e*、hsa-let-7f、hsa-let-7f-1*、hsa-let-7f-2*、hsa-let-7g、hsa-let-7g*、hsa-let-71、hsa-let-71*、hsa-mir-1、hsa-mir-100、hsa-mir-100*、hsa-mir-101、hsa-mir-101*、hsa-mir-103、hsa-mir-105、hsa-mir-105*、hsa-mir-106a、hsa-mir-106a*、hsa-mir-106b、hsa-mir-106b*、hsa-mir-107、hsa-mir-10a、hsa-mir-10a*、hsa-mir-10b、hsa-mir-10b*、hsa-mir-1178、hsa-mir-1179、hsa-mir-1180、hsa-mir-1181、hsa-mir-1182、hsa-mir-1183、hsa-mir-1184、hsa-mir-1185、hsa-mir-1197、hsa-mir-1200、hsa-mir-1201、hsa-mir-1202、hsa-mir-1203、hsa-mir-1204、hsa-mir-1205、hsa-mir-1206、hsa-mir-1207-3p、hsa-mir-1207-5p、hsa-mir-1208、hsa-mir-122、hsa-mir-122*、hsa-mir-1224-3p、hsa-mir-1224-5p、hsa-mir-1225-3p、hsa-mir-1225-5p、hsa-mir-1226、hsa-mir-1226*、hsa-mir-1227、hsa-mir-1228、hsa-mir-1228*、hsa-mir-1229、hsa-mir-1231、hsa-mir-1233、hsa-mir-1234、hsa-mir-1236、hsa-mir-1237、hsa-mir-1238、hsa-mir-124、hsa-mir-124*、hsa-mir-1243、hsa-mir-1244、hsa-mir-1245、hsa-mir-1246、hsa-mir-1247、hsa-mir-1248、hsa-mir-1249、hsa-mir-1250、hsa-mir-1251、hsa-mir-1252、hsa-mir-1253、hsa-mir-1254、hsa-mir-1255a、hsa-mir-1255b、hsa-mir-1256、hsa-mir-1257、hsa-mir-1258、hsa-mir-1259、hsa-mir-125a-3p、hsa-mir-125a-5p、hsa-mir-125b、hsa-mir-125b-1*、hsa-mir-125b-2*、hsa-mir-126、hsa-mir-126*、hsa-mir-1260、hsa-mir-1261、hsa-mir-1262、hsa-mir-1263、hsa-mir-1264、hsa-mir-1265、hsa-mir-1266、hsa-mir-1267、hsa-mir-1268、hsa-mir-1269、hsa-mir-1270、hsa-mir-1271、hsa-mir-1272、hsa-mir-1273、hsa-mir-127-3p、hsa-mir-1274a、hsa-mir-1274b、hsa-mir-1275、hsa-mir-127-5p、hsa-mir-1276、hsa-mir-1277、hsa-mir-1278、hsa-mir-1279、hsa-mir-128、hsa-mir-1280、hsa-mir-1281、hsa-mir-1282、hsa-mir-1283、hsa-mir-1284、hsa-mir-1285、hsa-mir-1286、hsa-mir-1287、hsa-mir-1288、hsa-mir-1289、hsa-mir-129*、hsa-mir-1290、hsa-mir-1291、hsa-mir-1292、hsa-mir-1293、hsa-mir-129-3p、hsa-mir-1294、hsa-mir-1295、hsa-mir-129-5p、hsa-mir-1296、hsa-mir-1297、hsa-mir-1298、hsa-mir-1299、hsa-mir-1300、hsa-mir-1301、hsa-mir-1302、hsa-mir-1303、hsa-mir-1304、hsa-mir-1305、hsa-mir-1306、hsa-mir-1307、hsa-mir-1308、hsa-mir-130a、hsa-mir-130a*、hsa-mir-130b、hsa-mir-130b*、hsa-mir-132、hsa-mir-132*、hsa-mir-1321、hsa-mir-1322、hsa-mir-1323、hsa-mir-1324、hsa-mir-133a、hsa-mir-133b、hsa-mir-134、hsa-mir-135a、hsa-mir-135a*、hsa-mir-135b、hsa-mir-135b*、hsa-mir-136、hsa-mir-136*、hsa-mir-137、hsa-mir-138、hsa-mir-138-1*、hsa-mir-138-2*、hsa-mir-139-3p、hsa-mir-139-5p、hsa-mir-140-3p、hsa-mir-140-5p、hsa-mir-141、hsa-mir-141*、hsa-mir-142-3p、hsa-mir-142-5p、hsa-mir-143、hsa-mir-143*、hsa-mir-144、hsa-mir-144*、hsa-mir-145、hsa-mir-145*、hsa-mir-146a、hsa-mir-146a*、hsa-mir-146b-3p、hsa-mir-146b-5p、hsa-mir-147、hsa-mir-147b、hsa-mir-148a、hsa-mir-148a*、hsa-mir-148b、hsa-mir-148b*、hsa-mir-149、hsa-mir-149*、hsa-mir-150、hsa-mir-150*、hsa-mir-151-3p、hsa-mir-151-5p、hsa-mir-152、hsa-mir-153、hsa-mir-154、hsa-mir-154*、hsa-mir-155、hsa-mir-155*、hsa-mir-15a、hsa-mir-15a*、hsa-mir-15b、hsa-mir-15b*、hsa-mir-16、hsa-mir-16-1*、hsa-mir-16-2*、hsa-mir-17、hsa-mir-17*、hsa-mir-181a、hsa-mir-181a*、hsa-mir-181a-2*、hsa-mir-181b、hsa-mir-181c、hsa-mir-181c*、hsa-mir-181d、hsa-mir-182、hsa-mir-182*、hsa-mir-1825、hsa-mir-1826、hsa-mir-1827、hsa-mir-183、hsa-mir-183*、hsa-mir-184、hsa-mir-185、hsa-mir-185*、hsa-mir-186、hsa-mir-186*、hsa-mir-187、hsa-mir-187*、hsa-mir-188-3p、hsa-mir-188-5p、hsa-mir-18a、hsa-mir-18a*、hsa-mir-18b、hsa-mir-18b*、hsa-mir-190、hsa-mir-190b、hsa-mir-191、hsa-mir-191*、hsa-mir-192、hsa-mir-192*、hsa-mir-193a-3p、hsa-mir-193a-5p、hsa-mir-193b、hsa-mir-193b*、hsa-mir-194、hsa-mir-194*、hsa-mir-195、hsa-mir-195*、hsa-mir-196a、hsa-mir-196a*、hsa-mir-196b、hsa-mir-197、hsa-mir-198、hsa-mir-199a-3p、hsa-mir-199a-5p、hsa-mir-199b-5p、hsa-mir-19a、hsa-mir-19a*、hsa-mir-19b、hsa-mir-19b-1*、hsa-mir-19b-2*、hsa-mir-200a、hsa-mir-200a*、hsa-mir-200b、hsa-mir-200b*、hsa-mir-200c、hsa-mir-200c*、hsa-mir-202、hsa-mir-202*、hsa-mir-203、hsa-mir-204、hsa-mir-205、hsa-mir-206、hsa-mir-208a、hsa-mir-208b、hsa-mir-20a、hsa-mir-20a*、hsa-mir-20b、hsa-mir-20b*、hsa-mir-21、hsa-mir-21*、hsa-mir-210、hsa-mir-211、hsa-mir-212、hsa-mir-214、hsa-mir-214*、hsa-mir-215、hsa-mir-216a、hsa-mir-216b、hsa-mir-217、hsa-mir-218、hsa-mir-218-1*、hsa-mir-218-2*、hsa-mir-219-1-3p、hsa-mir-219-2-3p、hsa-mir-219-5p、hsa-mir-22、hsa-mir-22*、hsa-mir-220a、hsa-mir-220b、hsa-mir-220c、hsa-mir-221、hsa-mir-221*、hsa-mir-222、hsa-mir-222*、hsa-mir-223、hsa-mir-223*、hsa-mir-224、hsa-mir-23a、hsa-mir-23a*、hsa-mir-23b、hsa-mir-23b*、hsa-mir-24、hsa-mir-24-1*、hsa-mir-24-2*、hsa-mir-25、hsa-mir-25*、hsa-mir-26a、hsa-mir-26a-1*、hsa-mir-26a-2*、hsa-mir-26b、hsa-mir-26b*、hsa-mir-27a、hsa-mir-27a*、hsa-mir-27b、hsa-mir-27b*、hsa-mir-28-3p、hsa-mir-28-5p、hsa-mir-296-3p、hsa-mir-296-5p、hsa-mir-297、hsa-mir-298、hsa-mir-299-3p、hsa-mir-299-5p、hsa-mir-29a、hsa-mir-29a*、hsa-mir-29b、hsa-mir-296-1*、hsa-mir-296-2*、hsa-mir-29c、hsa-mir-29c*、hsa-mir-300、hsa-mir-301a、hsa-mir-301b、hsa-mir-302a、hsa-mir-302a*、hsa-mir-302b、hsa-mir-302b*、hsa-mir-302c、hsa-mir-302c*、hsa-mir-302d、hsa-mir-302d*、hsa-mir-302e、hsa-mir-302f、hsa-mir-30a、hsa-mir-30a*、hsa-mir-30b、hsa-mir-30b*、hsa-mir-30c、hsa-mir-30c-1*、hsa-mir-30c-2*、hsa-mir-30d、hsa-mir-30d*、hsa-mir-30e、hsa-mir-30e*、hsa-mir-31、hsa-mir-31*、hsa-mir-32、hsa-mir-32*、hsa-mir-320a、hsa-mir-320b、hsa-mir-320c、hsa-mir-320d、hsa-mir-323-3p、hsa-mir-323-5p、hsa-mir-324-3p、hsa-mir-324-5p、hsa-mir-325、hsa-mir-326、hsa-mir-328、hsa-mir-329、hsa-mir-330-3p、hsa-mir-330-5p、hsa-mir-331-3p、hsa-mir-331-5p、hsa-mir-335、hsa-mir-335*、hsa-mir-337-3p、hsa-mir-337-5p、hsa-mir-338-3p、hsa-mir-338-5p、hsa-mir-339-3p、hsa-mir-339-5p、hsa-mir-33a、hsa-mir-33a*、hsa-mir-33b、hsa-mir-33b*、hsa-mir-340、hsa-mir-340*、hsa-mir-342-3p、hsa-mir-342-5p、hsa-mir-345、hsa-mir-346、hsa-mir-34a、hsa-mir-34a*、hsa-mir-34b、hsa-mir-34b*、hsa-mir-34c-3p、hsa-mir-34c-5p、hsa-mir-361-3p、hsa-mir-361-5p、hsa-mir-362-3p、hsa-mir-362-5p、hsa-mir-363、hsa-mir-363*、hsa-mir-365、hsa-mir-367、hsa-mir-367*、hsa-mir-369-3p、hsa-mir-369-5p、hsa-mir-370、hsa-mir-371-3p、hsa-mir-371-5p、hsa-mir-372、hsa-mir-373、hsa-mir-373*、hsa-mir-374a、hsa-mir-374a*、hsa-mir-374b、hsa-mir-374b*、hsa-mir-375、hsa-mir-376a、hsa-mir-376a*、hsa-mir-376b、hsa-mir-376c、hsa-mir-377、hsa-mir-377*、hsa-mir-378、hsa-mir-378*、hsa-mir-379、hsa-mir-379*、hsa-mir-380、hsa-mir-380*、hsa-mir-381、hsa-mir-382、hsa-mir-383、hsa-mir-384、hsa-mir-409-3p、hsa-mir-409-5p、hsa-mir-410、hsa-mir-411、hsa-mir-411*、hsa-mir-412、hsa-mir-421、hsa-mir-422a、hsa-mir-423-3p、hsa-mir-423-5p、hsa-mir-424、hsa-mir-424*、hsa-mir-425、hsa-mir-425*、hsa-mir-429、hsa-mir-431、hsa-mir-431*、hsa-mir-432、hsa-mir-432*、hsa-mir-433、hsa-mir-448、hsa-mir-449a、hsa-mir-449b、hsa-mir-450a、hsa-mir-450b-3p、hsa-mir-450b-5p、hsa-mir-451、hsa-mir-452、hsa-mir-452*、hsa-mir-453、hsa-mir-454、hsa-mir-454*、hsa-mir-455-3p、hsa-mir-455-5p、hsa-mir-483-3p、hsa-mir-483-5p、hsa-mir-484、hsa-mir-485-3p、hsa-mir-485-5p、hsa-mir-486-3p、hsa-mir-486-5p、hsa-mir-487a、hsa-mir-487b、hsa-mir-488、hsa-mir-488*、hsa-mir-489、hsa-mir-490-3p、hsa-mir-490-5p、hsa-mir-491-3p、hsa-mir-491-5p、hsa-mir-492、hsa-mir-493、hsa-mir-493*、hsa-mir-494、hsa-mir-495、hsa-mir-496、hsa-mir-497、hsa-mir-497*、hsa-mir-498、hsa-mir-499-3p、hsa-mir-499-5p、hsa-mir-500、hsa-mir-500*、hsa-mir-501-3p、hsa-mir-501-5p、hsa-mir-502-3p、hsa-mir-502-5p、hsa-mir-503、hsa-mir-504、hsa-mir-505、hsa-mir-505*、hsa-mir-506、hsa-mir-507、hsa-mir-508-3p、hsa-mir-508-5p、hsa-mir-509-3-5p、hsa-mir-509-3p、hsa-mir-509-5p、hsa-mir-510、hsa-mir-511、hsa-mir-512-3p、hsa-mir-512-5p、hsa-mir-513a-3p、hsa-mir-513a-5p、hsa-mir-513b、hsa-mir-513c、hsa-mir-514、hsa-mir-515-3p、hsa-mir-515-5p、hsa-mir-516a-3p、hsa-mir-516a-5p、hsa-mir-516b、hsa-mir-517*、hsa-mir-517a、hsa-mir-517b、hsa-mir-517c、hsa-mir-518a-3p、hsa-mir-518a-5p、hsa-mir-518b、hsa-mir-518c、hsa-mir-518c*、hsa-mir-518d-3p、hsa-mir-518d-5p、hsa-mir-518e、hsa-mir-518e*、hsa-mir-518f、hsa-mir-518f*、hsa-mir-519a、hsa-mir-519b-3p、hsa-mir-519c-3p、hsa-mir-519d、hsa-mir-519e、hsa-mir-519e*、hsa-mir-520a-3p、hsa-mir-520a-5p、hsa-mir-520b、hsa-mir-520c-3p、hsa-mir-520d-3p、hsa-mir-520d-5p、hsa-mir-520e、hsa-mir-520f、hsa-mir-520g、hsa-mir-520h、hsa-mir-521、hsa-mir-522、hsa-mir-523、hsa-mir-524-3p、hsa-mir-524-5p、hsa-mir-525-3p、hsa-mir-525-5p、hsa-mir-526b、hsa-mir-526b*、hsa-mir-532-3p、hsa-mir-532-5p、hsa-mir-539、hsa-mir-541、hsa-mir-541*、hsa-mir-542-3p、hsa-mir-542-5p、hsa-mir-543、hsa-mir-544、hsa-mir-545、hsa-mir-545*、hsa-mir-548a-3p、hsa-mir-548a-5p、hsa-mir-548b-3p、hsa-mir-5486-5p、hsa-mir-548c-3p、hsa-mir-548c-5p、hsa-mir-548d-3p、hsa-mir-548d-5p、hsa-mir-548e、hsa-mir-548f、hsa-mir-548g、hsa-mir-548h、hsa-mir-548i、hsa-mir-548j、hsa-mir-548k、hsa-mir-5481、hsa-mir-548m、hsa-mir-548n、hsa-mir-548o、hsa-mir-548p、hsa-mir-549、hsa-mir-550、hsa-mir-550*、hsa-mir-551a、hsa-mir-551b、hsa-mir-551b*、hsa-mir-552、hsa-mir-553、hsa-mir-554、hsa-mir-555、hsa-mir-556-3p、hsa-mir-556-5p、hsa-mir-557、hsa-mir-558、hsa-mir-559、hsa-mir-561、hsa-mir-562、hsa-mir-563、hsa-mir-564、hsa-mir-566、hsa-mir-567、hsa-mir-568、hsa-mir-569、hsa-mir-570、hsa-mir-571、hsa-mir-572、hsa-mir-573、hsa-mir-574-3p、hsa-mir-574-5p、hsa-mir-575、hsa-mir-576-3p、hsa-mir-576-5p、hsa-mir-577、hsa-mir-578、hsa-mir-579、hsa-mir-580、hsa-mir-581、hsa-mir-582-3p、hsa-mir-582-5p、hsa-mir-583、hsa-mir-584、hsa-mir-585、hsa-mir-586、hsa-mir-587、hsa-mir-588、hsa-mir-589、hsa-mir-589*、hsa-mir-590-3p、hsa-mir-590-5p、hsa-mir-591、hsa-mir-592、hsa-mir-593、hsa-mir-593*、hsa-mir-595、hsa-mir-596、hsa-mir-597、hsa-mir-598、hsa-mir-599、hsa-mir-600、hsa-mir-601、hsa-mir-602、hsa-mir-603、hsa-mir-604、hsa-mir-605、hsa-mir-606、hsa-mir-607、hsa-mir-608、hsa-mir-609、hsa-mir-610、hsa-mir-611、hsa-mir-612、hsa-mir-613、hsa-mir-614、hsa-mir-615-3p、hsa-mir-615-5p、hsa-mir-616、hsa-mir-616*、hsa-mir-617、hsa-mir-618、hsa-mir-619、hsa-mir-620、hsa-mir-621、hsa-mir-622、hsa-mir-623、hsa-mir-624、hsa-mir-624*、hsa-mir-625、hsa-mir-625*、hsa-mir-626、hsa-mir-627、hsa-mir-628-3p、hsa-mir-628-5p、hsa-mir-629、hsa-mir-629*、hsa-mir-630、hsa-mir-631、hsa-mir-632、hsa-mir-633、hsa-mir-634、hsa-mir-635、hsa-mir-636、hsa-mir-637、hsa-mir-638、hsa-mir-639、hsa-mir-640、hsa-mir-641、hsa-mir-642、hsa-mir-643、hsa-mir-644、hsa-mir-645、hsa-mir-646、hsa-mir-647、hsa-mir-648、hsa-mir-649、hsa-mir-650、hsa-mir-651、hsa-mir-652、hsa-mir-653、hsa-mir-654-3p、hsa-mir-654-5p、hsa-mir-655、hsa-mir-656、hsa-mir-657、hsa-mir-658、hsa-mir-659、hsa-mir-660、hsa-mir-661、hsa-mir-662、hsa-mir-663、hsa-mir-663b、hsa-mir-664、hsa-mir-664*、hsa-mir-665、hsa-mir-668、hsa-mir-671-3p、hsa-mir-671-5p、hsa-mir-675、hsa-mir-7、hsa-mir-708、hsa-mir-708*、hsa-mir-7-1*、hsa-mir-7-2*、hsa-mir-720、hsa-mir-744、hsa-mir-744*、hsa-mir-758、hsa-mir-760、hsa-mir-765、hsa-mir-766、hsa-mir-767-3p、hsa-mir-767-5p、hsa-mir-768-3p、hsa-mir-768-5p、hsa-mir-769-3p、hsa-mir-769-5p、hsa-mir-770-5p、hsa-mir-802、hsa-mir-873、hsa-mir-874、hsa-mir-875-3p、hsa-mir-875-5p、hsa-mir-876-3p、hsa-mir-876-5p、hsa-mir-877、hsa-mir-877*、hsa-mir-885-3p、hsa-mir-885-5p、hsa-mir-886-3p、hsa-mir-886-5p、hsa-mir-887、hsa-mir-888、hsa-mir-888*、hsa-mir-889、hsa-mir-890、hsa-mir-891a、hsa-mir-891b、hsa-mir-892a、hsa-mir-892b、hsa-mir-9、hsa-mir-9*、hsa-mir-920、hsa-mir-921、hsa-mir-922、hsa-mir-923、hsa-mir-924、hsa-mir-92a、hsa-mir-92a-1*、hsa-mir-92a-2*、hsa-mir-92b、hsa-mir-92b*、hsa-mir-93、hsa-mir-93*、hsa-mir-933、hsa-mir-934、hsa-mir-935、hsa-mir-936、hsa-mir-937、hsa-mir-938、hsa-mir-939、hsa-mir-940、hsa-mir-941、hsa-mir-942、hsa-mir-943、hsa-mir-944、hsa-mir-95、hsa-mir-96、hsa-mir-96*、hsa-mir-98、hsa-mir-99a、hsa-mir-99a*、hsa-mir-99b和hsa-mir-99b*。例如,可能所关注的是靶向染色体8开放阅读框72(c9orf72)的mirna,所述开放阅读框表达与肌萎缩侧索硬化症(als)相关的超氧化物歧化酶(sod1)。mirna抑制其靶向的mrna的功能,并且因此抑制由mrna编码的多肽的表达。因此,(部分地或全部地)阻断mirna的活性(例如,沉默mirna)可以有效地诱导或恢复表达被抑制的多肽的表达(使多肽去抑制)。在一个实施例中,通过多种方法中的任一种抑制细胞中的mirna活性来实现对由mirna的mrna靶标编码的多肽的去抑制。例如,可以通过与和mirna互补或基本上互补的小干扰核酸(例如,反义寡核苷酸、mirna海绵体、tudrna)杂交来实现阻断mirna的活性,由此阻断mirna与其靶mrna的相互作用。如本文所使用的,与mirna基本上互补的小干扰核酸是能够与mirna杂交并阻断mirna活性的小干扰核酸。在一些实施例中,与mirna基本上互补的小干扰核酸是除1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、11个、12个、13个、14个、15个、16个、17个或18个碱基之外完全与mirna互补的小干扰核酸。“mirna抑制剂”是阻断mirna功能、表达和/或加工的药剂。例如,这些分子包含但不限于抑制mirna与drosha复合物相互作用的微小rna特异性反义分子、微小rna海绵体、强诱饵rna(tudrna)和微小rna寡核苷酸(双链、发夹、短寡核苷酸)。仍其它有用的转基因可以包含对赋予病原体被动免疫的免疫球蛋白进行编码的转基因。“免疫球蛋白分子”是含有共价偶联在一起并且能够与抗原特异性组合的免疫球蛋白重链和免疫球蛋白轻链的免疫活性部分的蛋白质。免疫球蛋白分子可以是任何类型(例如,igg、ige、igm、igd、iga和igy)、类别(例如,igg1、igg2、igg3、igg4、iga1和iga2)或子类。术语“抗体”和“免疫球蛋白”在本文中可以互换使用。“免疫球蛋白重链”是含有免疫球蛋白的抗原结合结构域的至少一部分和免疫球蛋白重链的可变区的至少一部分或免疫球蛋白重链的恒定区的至少一部分的多肽。因此,免疫球蛋白源性重链与免疫球蛋白基因超家族的成员具有显著的氨基酸序列同源性区。例如,fab片段中的重链是免疫球蛋白源性重链。“免疫球蛋白轻链”是含有免疫球蛋白的抗原结合结构域的至少一部分和免疫球蛋白轻链的可变区的至少一部分或恒定区的至少一部分的多肽。因此,免疫球蛋白源性轻链与免疫球蛋白基因超家族的成员具有显著的氨基酸同源性区。“免疫粘附素”是嵌合的抗体样分子,所述分子将结合蛋白(通常是受体、配体或细胞粘附分子)的功能结构域与免疫球蛋白恒定结构域组合,所述免疫球蛋白恒定结构域通常包含铰链和fc区。“片段抗原结合(fab)片段”是与抗原结合的抗体上的区。其由重链和轻链中的每一个的一个恒定结构域和一个可变结构域构成。基于寻求针对其进行保护的疾病的病原体(causativeagent/pathogen)选择抗病原体构建体。这些病原体可以是病毒、细菌或真菌来源的,并且可以用于预防人感染人类疾病,或用于非人哺乳动物或其它动物以预防兽医疾病。raav可以包含对抗体以及具体地针对病毒病原体的中和抗体进行编码的基因。这种抗病毒抗体可以包含针对甲型流感、乙型流感和丙型流感中的一种或多种的抗流感抗体。a型病毒是最具毒性的人类病原体。与流行病相关的甲型流感的血清型包含:h1n1,其引起1918年的西班牙流感和2009年的猪流感;h2n2,其引起1957年的亚洲流感;h3n2,其引起1968年的香港流感;h5n1,其引起2004年的禽流感;h7n7;h1n2;h9n2;h7n2;h7n3;以及h10n7。其它靶病原性病毒包含:沙粒病毒(包含funin、马丘波病毒(machupo)和拉沙热病毒(lassa))、丝状病毒(包含马尔堡病毒(marburg)和埃博拉病毒(ebola))、汉坦病毒、小核糖核酸病毒(picornoviridae)(包含鼻病毒、艾柯病毒)、冠状病毒、副粘病毒、麻疹病毒、呼吸道合胞病毒、囊膜病毒、柯萨奇病毒、jc病毒、细小病毒b19、副流感病毒、腺病毒、呼吸道肠道病毒、来自痘病毒家族的天花病毒(variola)(大天花(天花(smallpox))和牛痘(vaccinia/cowpox)以及水痘-带状疱疹病毒(伪狂犬病)。病毒性出血热是由沙粒病毒家族(拉沙热)(此家族也与淋巴细胞性脉络丛脑膜炎病毒(lcm)相关)、丝状病毒(埃博拉病毒)和汉坦病毒(普马拉病毒(puremala))的成员引起的。小核糖核酸病毒(鼻病毒的亚家族)的成员与人普通感冒相关。冠状病毒家族包含多种非人类病毒,如传染性支气管炎病毒(家禽)、猪传染性胃肠炎病毒(猪)、猪血凝素脑脊髓炎病毒(猪)、猫传染性腹膜炎病毒(猫)、猫肠道冠状病毒(猫)、犬冠状病毒(狗)。已经推定人呼吸道冠状病毒与普通感冒、非甲型、乙型或丙型肝炎以及突发性急性呼吸综合症(sars)相关。副粘病毒家族包含副流感病毒1型、副流感病毒3型、牛副流感病毒3型、腮腺炎病毒属(腮腺炎病毒)、副流感病毒2型、副流感病毒4型、新城疫病毒(鸡)、牛瘟、麻疹病毒(包含麻疹和犬瘟热)和肺炎病毒(包含呼吸道合胞病毒(rsv)。细小病毒家族包含猫细小病毒(猫肠炎)、猫泛白细胞减少症病毒、犬细小病毒和猪细小病毒。腺病毒家族包含引起呼吸道疾病的病毒(ex、ad7、ard、o.b.)。因此,在某些实施例中,本文所描述的raav载体可以被工程化以表达抗埃博拉抗体(例如,2g4、4g7、13c6)、抗流感抗体(例如,fi6、cr8033)和抗rsv抗体(例如,帕利珠单抗(palivizumab)、莫维珠单抗(motavizumab))。也可以选择针对细菌病原体的中和抗体构建体用于本发明中。在一个实施例中,中和抗体构建体针对细菌本身。在另一个实施例中,中和抗体构建体针对由细菌产生的毒素。空气传播的细菌病原体的实例包含例如脑膜炎奈瑟氏球菌(neisseriameningitidis)(脑膜炎)、肺炎克雷伯菌(klebsiellapneumonia)(肺炎)、铜绿假单胞菌(pseudomonasaeruginosa)(肺炎)、伪鼻疽假单胞菌(pseudomonaspseudomallei)(肺炎)、鼻疽假单胞菌(pseudomonasmallei)(肺炎)、不动杆菌(肺炎)、卡他莫拉菌(moraxellacatarrhalis)、腔隙莫拉菌(moraxellalacunata)、产碱杆菌属(alkaligenes)、心杆菌属(cardiobacterium)、流感嗜血杆菌(haemophilusinfluenzae)(流感)、副流感嗜血杆菌(haemophilusparainfluenzae)、百日咳博代氏杆菌(bordetellapertussis)(百日咳)、土拉弗朗西斯菌(francisellatularensis)(肺炎/发烧)、肺炎军团菌(legionellapneumonia)(军团病)、鹦鹉热衣原体(chlamydiapsittaci)(肺炎)、肺炎衣原体(chlamydiapneumoniae)(肺炎)、结核分枝杆菌(mycobacteriumtuberculosis)(肺结核(tb))、堪萨斯分枝杆菌(mycobacteriumkansasii)(tb)、鸟分枝杆菌(mycobacteriumavium)(肺炎)、星状诺卡氏菌(nocardiaasteroides)(肺炎)、炭疽杆菌(bacillusanthracis)(炭疽)、金黄色葡萄球菌(staphylococcusaureus)(肺炎)、酿脓链球菌(streptococcuspyogenes)(猩红热)、肺炎链球菌(streptococcuspneumoniae)(肺炎)、白喉棒状杆菌(corynebacteriadiphtheria)(白喉)、肺炎支原体(mycoplasmapneumoniae)(肺炎)。raav可以包含对抗体以及具体地针对细菌病原体(如炭疽的病原体,即由炭疽杆菌产生的毒素)的中和抗体进行编码的基因。已经描述了针对形成类毒素的三种肽之一的保护剂(pa)的中和抗体。其它两种多肽由致死因子(lf)和水肿因子(ef)组成。抗pa中和抗体已经被描述为有效针对炭疽进行被动地免疫。参见例如美国专利第7,442,373号;r.sawada-hirai等人,《基于免疫的疫苗疗法杂志(jimmunebasedthervaccines)》2004;2:5.(2004年5月12日在线)。已经描述和/或可以产生仍其它抗炭疽毒素中和抗体。类似地,针对其它细菌和/或细菌毒素的中和抗体可以用于产生如本文所描述的aav递送性抗病原体构建体。针对传染病的抗体可以由寄生虫或真菌引起,所述真菌包含例如曲霉菌(aspergillusspecies)、伞枝犁头霉(absidiacorymbifera)、匍枝根霉(rhixpusstolonifer)、密丛毛霉(mucorplumbeaus)、新型隐球菌(cryptococcusneoformans)、荚膜组织胞浆菌(histoplasmcapsulatum)、皮炎芽生菌(blastomycesdermatitidis)、粗球孢子菌(coccidioidesimmitis)、青霉菌(penicilliumspecies)、干草小多孢菌(micropolysporafaeni)、普通高温放线菌(thermoactinomycesvulgaris)、互隔交链孢霉(alternariaalternate)、枝孢菌(cladosporiumspecies)、长蠕孢属(helminthosporium)和葡萄穗霉(stachybotrysspecies)。raav可以包含对抗体以及具体地针对以下疾病的致病因子的中和抗体进行编码的基因:如阿尔茨海默氏病(ad)、帕金森氏病(pd)、gba相关的帕金森氏病(gba-pd)、类风湿关节炎(ra)、肠易激综合症(ibs)、慢性阻塞性肺病(copd)、癌症、肿瘤、全身性硬化症、哮喘和其它疾病。此类抗体可以是但不限于例如α-突触核蛋白、抗血管内皮生长因子(vegf)(抗vegf)、抗vegfa、抗pd-1、抗pdl1、抗ctla-4、抗tnf-α、抗il-17、抗il-23、抗il-21、抗il-6、抗il-6受体、抗il-5、抗il-7、抗xii因子、抗il-2、抗hiv、抗ige、抗肿瘤坏死因子受体1(tnfr1)、抗-notch2/3、抗-notch1、抗ox40、抗erb-b2受体酪氨酸激酶3(erbb3)、抗erbb2、抗β细胞成熟抗原、抗b淋巴细胞刺激剂、抗cd20、抗her2、抗粒细胞巨噬细胞集落刺激因子、抗制瘤素m(osm)、抗淋巴细胞活化基因3(lag3)蛋白、抗ccl20、抗血清淀粉样蛋白p组分(sap)、抗脯氨酰羟化酶抑制剂、抗cd38、抗糖蛋白iib/iiia、抗cd52、抗-cd30、抗il-1β、抗表皮生长因子受体、抗cd25、抗rank配体、抗补体系统蛋白c5、抗cd11a、抗cd3受体、抗α-4(α4)整联蛋白、抗rsvf蛋白和抗整联蛋白α4β7。对于本领域的技术人员来说,仍其它病原体和疾病将是显而易见的。其它合适的抗体可以包含可用于治疗阿尔茨海默氏病的抗体,例如抗β淀粉样蛋白(例如,克雷内珠单抗(crenezumab)、索拉珠单抗(solanezumab)、阿杜卡尼单抗(aducanumab))、抗β淀粉样蛋白原纤维、抗β淀粉样蛋白斑块、抗τ、巴匹珠单抗(bapineuzamab)以及其它抗体。用于治疗多种适应症的其它合适的抗体包含描述于例如于2016年10月27日提交的pct/us2016/058968中公开为wo2017/075119a1的抗体。ii.raav载体产生供产生aav病毒载体(例如,重组(r)aav)之用,表达盒可以携带在递送到包装宿主细胞的任何合适的载体(例如,质粒)上。可在本发明使用的质粒可以被工程化,使得其适合于在原核细胞、昆虫细胞、哺乳动物细胞以及其它细胞中进行体外复制和包装。合适的转染技术和包装宿主细胞是已知的和/或可以由本领域的技术人员容易地设计。用于产生和分离适合于用作载体的aav的方法是本领域已知的。通常参见例如grieger和samulski,2005,“腺相关病毒作为基因疗法载体:载体开发、产生和临床应用(adeno-associatedvirusasagenetherapyvector:vectordevelopment,productionandclinicalapplications)”,《生化工程/生物技术进展(adv.biochem.engin/biotechnol)》99:119-145;buning等人,2008,“腺相关病毒载体技术的最新开发(recentdevelopmentsinadeno-associatedvirusvectortechnology),”《基因医学杂志(j.genemed)》10:717-733;以及下文引用的参考文献,这些参考文献中的每个参考文献通过引用整体并入本文中。为了将转基因包装到病毒粒子中,itr是在与含有表达盒的核酸分子相同的构建体中需要的顺式的唯一aav组分。cap和rep基因可以反式供应。在一个实施例中,本文所描述的表达盒被工程化到基因元件(例如,穿梭质粒),所述基因元件将其上携带的转基因构建体序列转移到包装宿主细胞中以产生病毒载体。在一个实施例中,所选基因元件可以通过任何合适的方法递送到aav包装细胞,所述方法包含转染、电穿孔、脂质体递送、膜融合技术、高速dna包被的粒料、病毒感染和原生质体融合。也可以制备稳定的aav包装细胞。可替代地,表达盒可以用于产生除aav之外的病毒载体。用于制备此类构建体的方法对核酸操纵技术人员而言是已知的并且包含基因工程、重组工程以及合成技术。参见例如《分子克隆:实验室手册》,由green和sambrook编辑,冷泉港实验室出版社,冷泉港,纽约(2012)。术语“aav中间体”或“aav载体中间体”是指缺少包装在其中的所期望的基因组序列的组装的raav衣壳。这些也可以被称为“空”衣壳。此类衣壳可以不含有表达盒的可检测基因组序列,或者含有不足以实现基因产物的表达的仅部分包装的基因组序列。这些空衣壳是无功能的以将所关注的基因转移到宿主细胞。可以使用已知的技术产生本文所描述的重组腺相关病毒(aav)。参见例如wo2003/042397;wo2005/033321、wo2006/110689;us7588772b2。此类方法涉及培养含有对aav衣壳蛋白进行编码的核酸序列的宿主细胞;功能性rep基因;至少由aav反向末端重复序列(itr)和转基因构成的表达盒;以及足以允许将表达盒包装到aav衣壳蛋白中的辅助功能。已经描述了产生衣壳的方法、其编码序列以及产生raav病毒载体的方法。参见例如gao等人,《美国国家科学院院刊》100(10):6081-6086(2003)和us2013/0045186a1。在一个实施例中,提供了一种可用于产生重组aav的产生细胞培养物。这种细胞培养物含有在宿主细胞中表达aav衣壳蛋白的核酸;适合于包装到aav衣壳中的核酸分子,例如含有aavitr和对基因产物进行编码的非aav核酸序列的载体基因组,所述非aav核酸序列可操作地连接到引导产物在宿主细胞中的表达的序列;以及足以允许将核酸分子包装到重组aav衣壳中的aavrep功能和腺病毒辅助功能。在一个实施例中,细胞培养物由哺乳动物细胞(例如,人胚肾293细胞以及其它细胞)或昆虫细胞(例如,杆状病毒)构成。任选地,rep功能由除提供衣壳的aav之外的aav提供。例如,rep可以是但不限于aav1rep蛋白、aav2rep蛋白、aav3rep蛋白、aav4rep蛋白、aav5rep蛋白、aav6rep蛋白、aav7rep蛋白、aav8rep蛋白;或rep78、rep68、rep52、rep40、rep68/78和rep40/52;或其片段;或另一种来源。任选地,rep和cap序列在细胞培养物中位于同一基因元件上。rep序列与cap基因之间可以存在间隔子。这些aav或突变aav衣壳序列中的任一种都可以在引导其在宿主细胞中表达的外源性调节控制序列的控制下。在一个实施例中,在合适的细胞培养物(例如,hek293)细胞中制造细胞。用于制造本文所描述的基因疗法载体的方法包含本领域众所周知的方法,如产生用于产生基因疗法载体的质粒dna、产生载体以及纯化载体。在一些实施例中,基因疗法载体是aav载体,并且所产生的质粒是对包含所关注的基因的载体基因组进行编码的aav顺式质粒、含有aavrep和cap基因的aav反式质粒以及腺病毒辅助质粒。载体产生过程可以包含方法步骤,如开始细胞培养、进行细胞传代、接种细胞、用质粒dna转染细胞、将转染后培养基交换为无血清培养基以及采集含载体的细胞和培养基。所采集的含载体的细胞和培养基在本文中被称为粗细胞采集物。在又另一个系统中,通过用基于杆状病毒的载体进行感染来将基因疗法载体引入到昆虫细胞中。关于这些产生系统的综述,通常参见例如zhang等人,2009,“用于大规模重组腺相关病毒产生的腺病毒-腺相关病毒杂合体(adenovirus-adeno-associatedvirushybridforlarge-scalerecombinantadeno-associatedvirusproduction)”,《人类基因疗法(humangenetherapy)》20:922-929,这些参考文献中的每个参考文献的内容通过引用整体并入本文中。在以下美国专利中也描述了制备和使用这些及其它aav产生系统的方法,这些美国专利中的每个美国专利的内容通过引用整体并入本文中:5,139,941;5,741,683;6,057,152;6,204,059;6,268,213;6,491,907;6,660,514;6,951,753;7,094,604;7,172,893;7,201,898;7,229,823;和7,439,065。此后,粗细胞采集物可以是本主题的方法步骤,如浓缩载体采集物、渗滤载体采集物、微流化载体采集物、核酸酶消化载体采集物、过滤经过微流化的中间体、通过色谱粗纯化、通过超速离心法粗纯化、通过切向流过滤进行缓冲液交换和/或调配和过滤以制备大量载体。在高盐浓度下进行两步亲和色谱纯化,然后使用阴离子交换树脂色谱来纯化载体药物产物并去除空衣壳。在国际专利公开第wo2017/160360号中更详细地描述了这些方法,其通过引用并入本文中。对于aav8,公开第wo2017/100676号中描述了纯化方法,并且对于rh10,国际专利公开第wo2017/100704号中描述了纯化方法,并且对于aav1,国际专利公开第wo2017/100674号中描述了纯化方法,这些国际专利公开通过引用全部并入本文中。为了计算空颗粒和完整颗粒的含量,将所选样品(例如,在本文的实例中经过碘克沙醇(iodixanol)梯度纯化的制剂,其中gc#=颗粒#)的vp3带体积相对于加载的gc颗粒进行作图。所得线性等式(y=mx+c)用于计算测试品峰的带状体积中的颗粒的数量。然后将加载的每20μl颗粒数量(pt)乘以50,以得到颗粒(pt)/ml。将pt/ml除以gc/ml得到颗粒与基因组拷贝的比率(pt/gc)。pt/ml-gc/ml得到空pt/ml。空pt/ml除以pt/ml并且×100得到空颗粒的百分比。通常,用于测定具有包装的基因组的空衣壳和aav载体颗粒的方法是本领域已知的。参见例如grimm等人,《基因疗法》(1999)6:1322-1330;sommer等人,《分子疗法(molec.ther.)》(2003)7:122-128。为了测试变性的衣壳,所述方法包含使经过处理的aav原液经受sds-聚丙烯酰胺凝胶电泳(由能够分离三种衣壳蛋白的任何凝胶组成,例如在缓冲液中含有3-8%三乙酸盐的梯度凝胶),然后运行凝胶直到分离出样品材料,并且将凝胶印迹到尼龙或硝酸纤维素膜(优选地是尼龙)上。然后,将抗aav衣壳抗体用作与变性的衣壳蛋白结合的初级抗体,优选地抗aav衣壳单克隆抗体,最优选地b1抗aav2单克隆抗体(wobus等人,《病毒学杂志》(2000)74:9281-9293)。然后使用次级抗体,所述次级抗体与初级抗体结合并且含有一种用于检测与初级抗体的结合的装置,更优选地是含有与其共价结合的检测分子的抗igg抗体,最优选地是与辣根过氧化物酶共价连接的绵羊抗小鼠igg抗体。一种用于检测结合的方法用于半定量地确定初级抗体与次级抗体之间的结合,优选地是能够检测放射性同位素发射、电磁辐射或比色变化的检测方法,最优选地是化学发光检测试剂盒。例如,对于sds-page,可以从柱级分中提取样品并在含有还原剂(例如,dtt)的sds-page上样缓冲液中加热,并且在预制的梯度聚丙烯酰胺凝胶(例如,novex)上解析衣壳蛋白。可以根据制造商的说明使用silverxpress(加利福尼亚州英杰公司)或其它合适的染色方法(即sypro红宝石色或考马斯染色)进行银染色。在一个实施例中,可以通过定量实时pcr(q-pcr)测量柱级分中的aav载体基因组(vg)的浓度。将样品稀释并用dnasei(或另一种合适的核酸酶)消化以去除外源性dna。在核酸酶失活后,使用引物和对引物之间的dna序列具有特异性的taqmantm荧光探针进一步稀释和扩增样品。在appliedbiosystemsprism7700序列检测系统上测量每种样品达到定义的荧光水平所需的周期的数量(阈值周期,ct)。含有与aav载体中所含序列相同的序列的质粒dna用于在q-pcr反应中产生标准曲线。从样品获得的周期阈值(ct)的值用于通过相对于质粒标准曲线的ct值对其进行归一化来确定载体基因组效价。也可以使用基于数字pcr的端点测定。一方面,使用了经过优化的q-pcr方法,所述方法利用了广谱丝氨酸蛋白酶,例如蛋白酶k(如可从凯杰公司(qiagen)商购获得)。更具体地,经过优化的qpcr基因组效价测定与标准测定类似,不同之处在于在dnasei消化之后,将样品用蛋白酶k缓冲液稀释并用蛋白酶k处理,然后进行热失活。合适地,以等于样品大小的量用蛋白酶k缓冲液稀释样品。蛋白酶k缓冲液可以浓缩2倍或更多倍。通常,蛋白酶k处理为约0.2mg/ml,但是可以在0.1g/ml到约1mg/ml之间变化。处理步骤通常在约55℃下进行持续约15分钟,但是可以在较低温度(例如,约37℃到约50℃)下进行持续较长的时间段(例如,约20分钟到约30分钟),或者在较高的温度(例如,最高约60℃)下进行持续较短的时间段(例如,约5到10分钟)。类似地,热失活通常在约95℃下持续约15分钟,但是温度可以降低(例如,约70℃到约90℃)并且时间延长(例如,约20分钟到约30分钟)。然后将样品稀释(例如,1000倍),并如标准测定中所描述的进行taqman分析。另外地或可替代地,可以使用液滴数字pcr(ddpcr)。例如,已经描述了用于通过ddpcr确定单链和自身互补aav载体基因组效价的方法。参见例如,m.lock等人,《人类基因疗法方法》2014年4月;25(2):115-25.doi:10.1089/hgtb.2013.131.电子出版2014年2月14日。简而言之,用于从基因组缺陷型aav中间体中分离具有包装的基因组序列的raav颗粒的方法涉及使包括重组aav病毒颗粒和aav衣壳蛋白中间体的悬浮液经受高效液相色谱,其中aav病毒颗粒和aav中间体与在高ph下平衡的强阴离子交换树脂结合,并经受盐梯度,同时监测洗脱液在约260和约280下的紫外线吸光度。可以基于所选aav调整ph。参见例如wo2017/160360(aav9)、wo2017/100704(aavrh10)、wo2017/100676(例如,aav8)和wo2017/100674(aav1),这些通过引用并入本文中。在此方法中,当a260/a280的比率达到拐点时,从洗脱的级分中收集aav完整衣壳。在一个实例中,对于亲和色谱步骤,可以将经过渗滤的产物应用于有效捕获aav2血清型的captureselecttmporos-aav2/9亲和树脂(生命科技公司(lifetechnologies))上。在这些离子条件下,显著百分比的残留的细胞dna和蛋白质流过柱,而aav颗粒则被有效捕获。iii.组合物和用途本文提供了一种组合物,所述组合物含有至少一种raav原液(例如,raav原液或突变raav原液)和任选的载剂、赋形剂和/或防腐剂。raav原液是指多个raav载体,所述多个raav载体的量与例如在下文关于浓度和剂量单位的讨论中描述的量相同。如本文所使用的,“载剂”包含任何和所有溶剂、分散介质、媒剂、涂层、稀释剂、抗细菌剂和抗真菌剂、等渗剂和吸收延迟剂、缓冲剂、载剂溶液、悬浮液、胶质物等。这种介质和药剂用于药物活性物质的用途在本领域中是熟知的。补充性活性成分也可以并入到组合物中。短语“药学上可接受的”是指当向宿主施用时不会产生过敏或类似不良反应的分子实体和组合物。递送媒剂(如脂质体、纳米胶囊、微颗粒、微球、脂质颗粒、囊泡等)可以用于将本发明的组合物引入到合适的宿主细胞中。特别地,raav载体递送的转基因可以被调配用于递送,所述转基因被包封在脂质颗粒、脂质体、囊泡、纳米球或纳米颗粒等中。在一个实施例中,组合物包含适合于递送到受试者的最终调配物,所述组合物是例如缓冲到生理上相容的ph和盐浓度的水性液体悬浮液。任选地,调配物中存在一种或多种表面活性剂。在另一个实施例中,可以将组合物作为稀释以施用于受试者的浓缩物运输。在其它实施例中,可以在施用时将组合物冻干并重构。可以从无毒的非离子表面活性剂中选择合适的表面活性剂或表面活性剂的组合。在一个实施例中,选择终止于伯羟基的双官能嵌段共聚物表面活性剂,例如f68[basf],也被称为泊洛沙姆(poloxamer)188,其具有中性ph,平均分子量为8400。可以选择其它表面活性剂和其它泊洛沙姆,即非离子型三嵌段共聚物,所述非离子型三嵌段共聚物由与聚氧乙烯(聚(环氧乙烷))的两个亲水链侧接的聚氧丙烯(聚(环氧丙烷))的中心疏水链、solutolhs15(聚乙二醇-15羟基硬脂酸酯)、labrasol(聚氧辛酸甘油酯)、聚氧10油醚、tween(聚氧乙烯山梨聚糖脂肪酸酯)、乙醇和聚乙二醇构成。在一个实施例中,调配物含有泊洛沙姆。这些共聚物通常以字母“p”(对于泊洛沙姆)命名,后跟三个数字:前两位数字×100给出了聚氧丙烯核的近似分子量,并且最后一位数字×10给出了聚氧乙烯含量的百分比。在一个实施例中,选择了泊洛沙姆188。表面活性剂可以以悬浮液的至多约0.0005%到约0.001%的量存在。以足够的量施用载体以转染细胞并提供足够水平的基因转移和表达,以便提供治疗益处,而不会产生过度的副作用或具有医学上可接受的生理作用,这可以由医学领域的技术人员确定。常规和药学上可接受的施用途径包含但不限于直接递送到所期望的器官(例如,肝脏(任选地通过肝动脉)、肺、心脏、眼睛、肾脏)、口服、吸入、鼻内、鞘内、气管内、动脉内、直接递送到眼睛(任选地通过眼部递送、视网膜下注射、视网膜内注射、玻璃体内、局部)、静脉内、肌内、皮下、皮内和其它亲本施用途径。在一个实施例中,施用途径是视网膜下或玻璃体内注射。如果需要的话,可以组合施用途径。病毒载体的剂量将主要取决于如所治疗的病状、患者的年龄、体重和健康状况等因素,并且因此在患者之间可能有所不同。例如,病毒载体的治疗有效人剂量通常在约25到约1000微升到约100ml溶液范围内,所述溶液含有浓度为约1×109到1×1016基因组病毒载体。将调整剂量以平衡治疗益处与任何副作用,并且这种剂量可以根据采用重组载体的治疗应用而变化。可以监测转基因的表达水平以确定所得病毒载体的剂量频率,优选地含有迷你基因的aav载体。任选地,与出于治疗目的描述的剂量方案类似的剂量方案可以用于使用本发明的组合物进行免疫。可以以剂量单位调配复制缺陷型病毒组合物,以使含有的复制缺陷型病毒的量在约1.0×109gc到约1.0×1016gc的范围内(以治疗平均体重70kg的受试者),包含所述范围内的所有整数或分数量,并且对于人类患者而言,优选地为1.0×1012gc到1.0×1014gc。在一个实施例中,将组合物调配成每剂量含有至少1×109、2×109、3×109、4×109、5×109、6×109、7×109、8×109或9×109gc,包含所述范围内的所有整数或分数量。在另一个实施例中,将组合物调配成每剂量含有至少1×1010、2×1010、3×1010、4×1010、5×1010、6×1010、7×1010、8×1010或9×1010gc,包含所述范围内的所有整数或分数量。在另一个实施例中,将组合物调配成每剂量含有至少1×1011、2×1011、3×1011、4×1011、5×1011、6×1011、7×1011、8×1011或9×1011gc,包含所述范围内的所有整数或分数量。在另一个实施例中,将组合物调配成每剂量含有至少1×1012、2×1012、3×1012、4×1012、5×1012、6×1012、7×1012、8×1012或9×1012gc,包含所述范围内的所有整数或分数量。在另一个实施例中,将组合物调配成每剂量含有至少1×1013、2×1013、3×1013、4×1013、5×1013、6×1013、7×1013、8×1013或9×1013gc,包含所述范围内的所有整数或分数量。在另一个实施例中,将组合物调配成每剂量含有至少1×1014、2×1014、3×1014、4×1014、5×1014、6×1014、7×1014、8×1014或9×1014gc,包含所述范围内的所有整数或分数量。在另一个实施例中,将组合物调配成每剂量含有至少1×1015、2×1015、3×1015、4×1015、5×1015、6×1015、7×1015、8×1015或9×1015gc,包含所述范围内的所有整数或分数量。在一个实施例中,对于人类应用,剂量的范围可以为每剂量1×1010到约1×1012gc,包含所述范围内的所有整数或分数量。这些上述剂量可以以各种体积的载剂、赋形剂或缓冲剂调配物施用,范围为约25到约1000微升或更大的体积,包含所述范围内的所有数字,这取决于待治疗区域的大小、所使用的病毒效价、施用途径以及所述方法的所期望的效果。在一个实施例中,载剂、赋形剂或缓冲剂的体积为至少约25μl。在一个实施例中,体积为约50μl。在另一个实施例中,体积为约75μl。在另一个实施例中,体积为约100μl。在另一个实施例中,体积为约125μl。在另一个实施例中,体积为约150μl。在另一个实施例中,体积为约175μl。在又另一个实施例中,体积为约200μl。在另一个实施例中,体积为约225μl。在又另一个实施例中,体积为约250μl。在又另一个实施例中,体积为约275μl。在又另一个实施例中,体积为约300μl。在又另一个实施例中,体积为约325μl。在另一个实施例中,体积为约350μl。在另一个实施例中,体积为约375μl。在另一个实施例中,体积为约400μl。在另一个实施例中,体积为约450μl。在另一个实施例中,体积为约500μl。在另一个实施例中,体积为约550μl。在另一个实施例中,体积为约600μl。在另一个实施例中,体积为约650μl。在另一个实施例中,体积为约700μl。在另一个实施例中,体积介于约700与约1000μl之间。在某些实施例中,剂量可以在约1×109gc/g脑质量到约1×1012gc/g脑质量的范围内。在某些实施例中,剂量可以在约3×1010gc/g脑质量到约3×1011gc/g脑质量的范围内。在某些实施例中,剂量可以在约5×1010gc/g脑质量到约1.85×1011gc/g脑质量的范围内。另一方面,提供了适合施用于受试者的水性悬浮液。在一个实施例中,悬浮液包含水性悬浮液体和约1×109个病毒颗粒到约1×1013gc或病毒颗粒每只眼睛的如本文所描述的重组腺相关病毒(raav),所述raav可用作治疗或预防眼部疾病的治疗剂。在一个实施例中,悬浮液适合于视网膜下或玻璃体内注射。在一个实施例中,病毒构建体可以以至少约1×109gc到约1×1015gc或约1×1011gc到5×1013gc的剂量递送。可以由本领域的技术人员确定用于递送这些剂量和浓度的合适的体积。例如,可以选择约1μl到150ml的体积,其中对于成人而言,选择更大的体积。通常,对于新生婴儿,合适的体积为约0.5ml到约10ml,对于年龄较大的婴儿,可以选择约0.5ml到约15ml。对于幼儿,可以选择约0.5ml到约20ml的体积。对于儿童,可以选择最大约30ml的体积。对于青春期前的少年和青少年,可以选择最大约50ml的体积。在仍其它实施例中,患者可以接受鞘内施用的体积选择为约5ml到约15ml或约7.5ml到约10ml。可以确定其它合适的体积和剂量。将调整剂量以平衡治疗益处与任何副作用,并且这种剂量可以根据采用重组载体的治疗应用而变化。可以根据所公开的方法将上文描述的重组载体递送到宿主细胞。可以将优选地悬浮于生理上相容的载剂中的raav施用于人或非人哺乳动物患者。在某些实施例中,为了施用于人类患者,将raav适当地悬浮于含有盐水、表面活性剂和生理上相容的盐或盐的混合物的水溶液中。合适地,将调配物调整到生理上可接受的ph,例如,在ph6到9、或ph6.5到7.5、ph7.0到7.7或ph7.2到7.8的范围内。由于脑脊液的ph为约7.28到约7.32,对于鞘内递送,可能期望在此范围内的ph;而对于静脉内、视网膜下或玻璃体内注射,可能期望约6.8到约7.2的ph。然而,可以选择最宽范围和这些子范围内的其它ph用于其它递送途径。在另一个实施例中,组合物包含载剂、稀释剂、赋形剂和/或佐剂。鉴于转移病毒所针对的适应症,本领域的技术人员可以容易地选择合适的载剂。例如,一种合适的载剂包含盐水,其可以与多种缓冲溶液(例如,磷酸盐缓冲盐水)一起调配。其它示例性载剂包含无菌盐水、乳糖、蔗糖、磷酸钙、明胶、葡聚糖、琼脂、果胶、花生油、芝麻油和水。缓冲液/载剂应包含防止raav粘附到输液管上但不干扰raav体内结合活性的组分。可以从无毒的非离子表面活性剂中选择合适的表面活性剂或表面活性剂的组合。在一个实施例中,选择终止于伯羟基的双官能嵌段共聚物表面活性剂,例如f68[basf],也被称为泊洛沙姆(poloxamer)188,其具有中性ph,平均分子量为8400。可以选择其它表面活性剂和其它泊洛沙姆,即非离子型三嵌段共聚物,所述非离子型三嵌段共聚物由与聚氧乙烯(聚(环氧乙烷))的两个亲水链侧接的聚氧丙烯(聚(环氧丙烷))的中心疏水链、solutolhs15(聚乙二醇-15羟基硬脂酸酯)、labrasol(聚氧辛酸甘油酯)、聚氧油醚、tween(聚氧乙烯山梨聚糖脂肪酸酯)、乙醇和聚乙二醇构成。在一个实施例中,调配物含有泊洛沙姆。这些共聚物通常以字母“p”(对于泊洛沙姆)命名,后跟三个数字:前两位数字×100给出了聚氧丙烯核的近似分子量,并且最后一位数字×10给出了聚氧乙烯含量的百分比。在一个实施例中,选择了泊洛沙姆188。表面活性剂可以以悬浮液的至多约0.0005%到约0.001%的量存在。在一个实例中,调配物可以含有例如缓冲盐水溶液,所述缓冲盐水溶液包括水中的氯化钠、碳酸氢钠、葡聚糖、硫酸镁(例如,硫酸镁·7h2o)、氯化钾、氯化钙(例如,氯化钙·2h2o)、磷酸氢二钠及其混合物中的一种或多种。合适地,对于鞘内递送,同渗浓摩在与脑脊液相容的范围内(例如,约275到约290);参见例如emedicine.medscape.com/article/2093316-overview。任选地,对于鞘内递送,可以将可商购获得的稀释剂用作悬浮剂,或与另一种悬浮剂和其它任选的赋形剂组合。参见例如elliotts解决方案[lukaremedical]。在其它实施例中,调配物可以含有一种或多种渗透增强剂。合适的渗透增强剂的实例可以包含例如甘露醇、甘氨胆酸钠、牛磺胆酸钠、脱氧胆酸钠、水杨酸钠、辛酸钠、癸酸钠、月桂基硫酸钠、聚氧乙烯-9-月桂基醚或edta。任选地,本发明的组合物除raav和一种或多种载剂之外还可以含有其它常规药物成分,如防腐剂或化学稳定剂。合适的示例性防腐剂包含氯丁醇、山梨酸钾、山梨酸、二氧化硫、没食子酸丙酯、对羟基苯甲酸甲酯、乙基香草醛、甘油、苯酚和对氯苯酚。合适的化学稳定剂包含明胶和白蛋白。根据本发明的组合物可以包括药学上可接受的载剂,如上文所定义的。合适地,本文所描述的组合物包括有效量的一种或多种aav,所述一种或多种aav悬浮于药学上合适的载剂中和/或与合适的赋形剂混合,所述合适的赋形剂被设计成通过注射、渗透泵、鞘内导管递送到受试者或通过另一种装置或途径递送。在一个实例中,组合物被调配用于鞘内递送。如本文所使用的,术语“鞘内递送”或“鞘内施用”是指通过注射到椎管中,更具体地注射到蛛网膜下腔中以便其到达脑脊液(csf)的药物施用途径。鞘内递送可以包含腰椎穿刺、心室内(包含脑室内(icv))、枕骨下/脑池内和/或c1-2穿刺。例如,可以通过腰椎穿刺引入材料以在整个蛛网膜下腔扩散。在另一个实例中,可以向小脑延髓池(cisternamagna)中注射。如本文所使用的,术语“脑池内递送”或“脑池内施用”是指药物直接进入到小脑延髓池(cisternamagnacerebellomedularis)的脑脊液中,更具体地是通过枕骨下穿刺或通过直接注射到小脑延髓池(cisternamagna)中或通过永久定位的管的施用途径。一方面,本文所提供的载体可以通过所述方法和/或装置鞘内施用。参见例如wo2017/181113,其通过引用并入本文中。可替代地,可以选择其它装置和方法。所述方法包括以下步骤:使脊髓针前进到患者的小脑延髓池(cisternamagna)中;将一定长度的柔性管连接到脊髓针的近侧毂,并且将阀的输出端口连接到柔性管的近端,并且在进行了所述前进和连接步骤之后,并在允许管将使用患者的脑脊液进行自身引物后,将含有一定量等渗溶液的第一容器连接到阀的冲洗入口,并且然后将含有一定量的药物组合物的第二容器连接到阀的载体入口。在将第一血管和第二血管连接到阀之后,在阀的载体入口与出口之间打开流体流动路径,并将药物组合物通过脊髓针注射到患者中,并且在注射药物组合物之后,通过阀的冲洗入口和出口打开流体流动路径,并将等渗溶液注射到脊髓针中以将药物组合物冲洗到患者中。此方法和此装置可以各自任选地用于鞘内递送本文所提供的组合物。可替代地,其它方法和装置可以用于此类鞘内递送。应注意的是,术语“一个(a)”或“一个(an)”是指一个或多个。如此,术语“一个(a或an)”、“一个或多个”和“至少一个”在本文中可互换使用。词语“包括(comprise)”、“包括(comprises)”和“包括(comprising)”将被解释为是包含性而非排他性的。词语“由……组成(consist)”、“由……组成(consisting)”及其变体将被解释为是排他性的而非包含性。虽然说明书中的各个实施例是使用“包括”语言来呈现的,但是在其它情况下,也意图使用“由……组成”或“基本上由……构成”的语言来解释和描述相关的实施例。如本文所使用的,除非另有说明,否则术语“约”意指相对于给定参考的10%(±10%)的变化性。如本文所使用的,“疾病”、“病症”和“病状”可互换地用于指示受试者的异常状态。除非在本说明书中另有定义,否则本文所使用的技术术语和科学术语具有与本领域的普通技术人员和参照公开文本所通常理解的相同含义,这为本领域的技术人员提供了本申请中使用的许多术语的通用指南。术语“表达”在本文中以其最广泛的含义使用,并且包括rna或rna和蛋白质的产生。关于rna,术语“表达”或“转译”尤其涉及肽或蛋白质的产生。表达可以是暂时的或可以是稳定的。如本文所使用的,术语“nab效价”是产生多少中和抗体(例如,抗aavnab)的量度,所述中和抗体中和其靶向的表位(例如,aav)的生理作用。抗aavnab效价可以如在calcedo,r等人,“针对腺相关病毒的中和抗体的世界范围的流行病(worldwideepidemiologyofneutralizingantibodiestoadeno-associatedviruses)”《传染病杂志(journalofinfectiousdiseases)》,2009.199(3):第381-390页中所描述的那样进行测量,其通过引用并入本文中。如本文所使用的,“表达盒”是指包括编码序列、启动子并可以包含其的其它调节序列的核酸分子,所述盒可以通过基因元件(例如,质粒)递送到包装宿主细胞,并包装到病毒载体(例如,病毒颗粒)的衣壳中。通常,用于产生病毒载体的这种表达盒含有本文所描述的基因产物的编码序列,所述编码序列侧接病毒基因组的包装信号和其它表达控制序列,如本文描述的序列。缩写“sc”是指自身互补的。“自身互补aav”是指其中由重组aav核酸序列所携带的编码区已经被设计成形成分子内双链dna模板的构建体。感染后,未等待细胞介导的第二条链合成,而是两条互补的半scaav将缔合以形成易于立即复制和转录的一条双链dna(dsdna)。参见例如dmmccarty等人,“自身互补重组腺相关病毒(scaav)载体独立于dna合成而促进高效转导(self-complementaryrecombinantadeno-associatedvirus(scaav)vectorspromoteefficienttransductionindependentlyofdnasynthesis)”,《基因疗法》,(2001年8月),第8卷,第16期,第1248-1254页。自身互补aav在例如美国专利第6,596,535号;第7,125,717号;第7,456,683号中描述,这些美国专利中的每个美国专利通过引用整体并入本文中。如本文所使用的,术语“可操作地连接”是指与所关注的基因邻接的表达控制序列以及以反式或在远处起作用以控制所关注的基因的表达控制序列两者。术语“异源性”当结合蛋白质或核酸使用时表明蛋白质或核酸包含在自然界中未发现彼此间的相同关系的两个或更多个序列或子序列。例如,核酸通常是重组地产生的,具有来自不相关基因的布置成产生新的功能性核酸的两个或更多个序列。例如,在一个实施例中,核酸具有来自一种基因的布置成引导编码序列从不同基因表达的启动子。因此,关于编码序列,启动子是异源的。“复制缺陷型病毒”或“病毒载体”是指其中含有所关注的基因的表达盒包装在病毒衣壳或包膜中的合成或人工病毒粒子,其中也包装在病毒衣壳或包膜内的任何病毒基因组序列均是复制缺陷型的;即,其不能产生子代病毒粒子,但保留了感染靶细胞的能力。在一个实施例中,病毒载体的基因组不包含对复制所需的酶进行编码的基因(基因组可以被工程化成“无肠的(gutless)”-仅含有所关注的转基因,其侧接扩增和包装人工基因组所需的信号),但是这些基因可以在产生期间供应。因此,这被认为可以安全地用于基因疗法,因为除非存在复制所需的病毒酶,否则不会发生通过子代病毒粒子进行的复制和感染。在许多情况下,raav颗粒被称为dnase抗性的。然而,除此核酸内切酶(dnase)之外,其它核酸内切酶和核酸外切酶也可以用于本文所描述的纯化步骤中,以去除污染性核酸。可以选择此类核酸酶以降解单链dna和/或双链dna以及rna。此类步骤可以含有单个核酸酶或针对不同靶标的核酸酶的混合物,并且可以是核酸内切酶或核酸外切酶。术语“抗核酸酶”表示aav衣壳已经在表达盒周围完全组装,所述表达盒被设计成将转基因递送到宿主细胞并保护这些包装的基因组序列在被设计成去除产生过程中可能存在的污染性核酸的核酸酶温育步骤期间免于降解(消化)。在本发明的上下文中,术语“转译”涉及核糖体的过程,其中mrna链控制氨基酸序列的组装以产生蛋白质或肽。如贯穿本说明书和权利要求所使用的,术语“包括”和“包含”包含其它组分、元件、整数、步骤等。相反,术语“由……组成”及其变体不包含其它组分、元件、整数、步骤等。如上文所描述的,除非另有说明,否则术语“约”在用于修改数值时意指±10%的变化。以下实例仅是说明性的,并且不旨在限制本发明。实例以下实例报告了aav8和另外的不同aav血清型的广泛脱酰胺化以及来自结构、生物化学和质谱方法的支持证据。每个位点处的脱酰胺化程度取决于载体的年龄以及多个主要序列和3d结构因素,但在很大程度上不受载体回收和纯化的条件制约。证明脱酰胺化可能影响载体转导活性,并使载体活性的早期时间点丧失与在若干种aav8天冬酰胺处快速进展的自发脱酰胺化相关。探索了稳定侧链酰胺的突变策略,从而改善载体转导并减少批次间分子变异性,所述批次间分子变异性是生物制备中的关键问题。这项研究展示了aav衣壳异质性的先前未知的方面,并突出显示了其在开发这些用于基因疗法的载体中的重要性。实例1中是通过一维和二维凝胶电泳、质谱和从头结构建模提供了对aav8载体衣壳的转译后修饰的表征。在衣壳表面上鉴定了多个推定的脱酰胺化位点之后,评估了其在体内和体外两者对衣壳结构和功能的影响。实例1进一步将此分析扩展到aav9,以确定此现象是否适用于除aav8之外的血清型,从而证实了aav衣壳的脱酰胺化不是血清型特异性的。实例2到5示出了不同的aav中的脱酰胺化。实例1:腺相关病毒衣壳的表面上的氨基酸的脱酰胺化a.材料与方法1.1d和2d凝胶电泳为了进行1dsds聚丙烯酰胺凝胶电泳(sds-page)分析,首先在存在十二烷基硫酸锂和还原剂的情况下在80℃下使aav载体变性持续20分钟。然后,在200v下在4-12%bis-tris凝胶上将其运行90分钟,并用考马斯兰染色。对于图1a-图1d中的数据,肯德里克实验室有限公司(kendricklaboratories,inc.)(威斯康星州麦迪逊)进行了2d凝胶电泳。对于后续实验,在内部进行了2dsds-page。为此,将3×1011gc的aav载体与500uturbonuclease标志物(加利福尼亚州圣地亚哥安诺伦公司(accelagen,sandiego,ca))在150μl含35mmnacl和1mmmgcl2的磷酸盐缓冲盐水(pbs)中组合在一起,并在37℃下温育持续十分钟。接下来,添加九个体积的无水乙醇,将样品涡旋,并且然后在-80℃下将其温育持续至少两小时,然后在冰上温育持续五分钟,并且然后在15℃下以最大速度离心持续30分钟。倒出上清液并风干沉淀物,然后将所述沉淀物重新悬浮于重悬浮缓冲液#1[含0.15%sds,50mm二硫苏糖醇(dtt)、10mmtrisph7.5和1μlph6-9两性电解质(赛默飞世尔科技公司zm0023)的ddh2o,每天添加一次]中,并在室温下不受干扰地温育。在30分钟之后,轻弹样品管以将其混合,添加1μg鸡伴清蛋白标志物(密苏里州圣路易斯西格玛奥德里奇公司(sigmaaldrich,st.louis,mo)),并在37℃下温育样品持续30分钟,轻弹以混合15分钟。然后将样品转移到50℃下持续15-20分钟,进行涡旋,在95℃下温育持续2.5分钟,并且在以最大速度离心一分钟之前允许其冷却,并且进行短暂涡旋。然后将10μl每种样品与140μl重悬浮缓冲液#2(含9.7m脲,2%chaps,0.002%溴酚蓝和0.05%两性电解质的ddh2o,如上文所述的,每天添加一次)混合并在室温下温育持续十分钟。然后,将混合物应用于固定在ph6-10的ph梯度(ipg)条带(马萨诸塞州沃尔瑟姆赛默飞世尔科技公司),并根据制造商的说明在zoomipgrunner系统上运行。使用了以下等电聚焦参数:在100-1,000v下持续120分钟,在1,000-2,000v下持续120分钟,在2,000v下持续120分钟,每个条带运行的极限为0.1w和0.05ma。然后,将ipg条带还原并在单孔4-12%bis-tris凝胶中上样,并如上文所描述的运行1d。通过将内部对照蛋白turbonuclease(安诺伦公司,27kda)与鸡蛋清伴清蛋白(西格玛奥德里奇公司,76kda,pi6.0-6.6)进行比较,确定了aavvp的相对迁移。2.载体产生宾夕法尼亚大学的载体核心公司(vectorcore)生产了用于1d和2d凝胶电泳和质谱实验的重组aav载体,并如先前所描述的通过氯化铯或碘克沙醇梯度对其进行纯化。(lockm等人,《人类基因疗法》2010;21(10):1259-71;gaogp等人,《美国国家科学院院刊》2002;99(18):11854-9)。产生了经过亲和力纯化的载体,如下:使hek293细胞在十个36层超堆叠容器(康宁公司(corning))中生长,用载体基因组质粒(paav-lsp-ivs2.hfixco-wpre-bgh)、含有aav2rep和aav8cap基因的反式质粒和腺病毒辅助质粒的混合物对其进行共转染。使用peipro(polyplus)作为转染试剂。转染后五天,采集上清液,通过sartoguardpesmidicap过滤器(赛多利斯斯泰帝公司(sartoriusstedim))进行澄清,并用苯并酶(benzonase)(密理博公司(millipore))处理,然后添加盐以使其达到0.6m。通过切向流过滤(tff)将澄清的块状采集材料浓缩十倍,并且然后针对四个体积的亲和柱上样缓冲液进行渗滤。在poroscaptureselect(赛默飞世尔科技公司)亲和柱上捕获载体,并在低ph下将载体峰直接洗脱到中和缓冲液中。将中和的洗脱液稀释到高ph的结合缓冲液中,并且将其上样到阴离子交换精制柱(cimultusqa-8;bia分离)上,其中制剂富集了含基因组的(完整)颗粒。用浅盐洗脱梯度洗脱完整的载体颗粒,并立即进行中和。最后,使载体经受第二轮tff,以进行最终浓缩,并将缓冲液交换成调配物缓冲液(pbs+0.001%普朗尼克(pluronic)f-68)。通过六孔板中的hek293细胞的小规模三重转染产生用于体外测定的突变载体。将5.6μl1mg/ml聚乙烯亚胺溶液在90μl无血清培养基中与质粒dna(在90μl无血清培养基中,0.091μg顺式质粒,0.91μg反式质粒,1.82μgδf6ad-辅助质粒)混合,将其在室温下温育持续15分钟,并且将其添加到细胞中以及另外0.8ml新鲜的无血清培养基中。第二天,用完全血清培养基代替0.5ml的顶部培养基。转染后三天,通过三个冷冻/融化循环采集载体,然后离心以去除细胞碎片并收集上清液。顺式质粒含有转基因盒,所述转基因盒在鸡β-肌动蛋白(cb7)启动子以及普洛麦格(promega)嵌合内含子和兔β-珠蛋白(rbg)聚腺苷酸化信号的控制下对萤火虫荧光素酶转基因进行编码。反式质粒对wtaav8cap基因进行编码;为了产生突变aav8cap变体,使用了quikchangelightningmutagenesis试剂盒(特拉华州威明顿市安捷伦科技公司(agilenttechnologies,wilmington,de)。如先前所描述的滴定载体。(lockm等人,《人类基因疗法》2010;21(10):1259-71)。对于时程载体产生实验,通过hek293细胞在15cm组织培养皿中的中等规模的三重转染产生载体。对于每个板,将36μl1mg/ml聚乙烯亚胺溶液在2ml无血清的培养基中与质粒dna(0.6μg顺式质粒,5.8μg反式质粒,11.6μgδf6ad-辅助质粒)混合,在室温下温育持续15分钟,并且在用14ml无血清培养基更新的板上将其以约60%汇合度添加到细胞中。接下来的一天,用新鲜的全血清培养基替代8ml的顶部培养基。通过收集所有顶部培养基,从培养皿中刮下细胞并将其冷冻在-80℃下来采集载体。通过应用3次冷冻/融化循环并通过离心澄清裂解物,从上清液/细胞混合物中回收粗载体。通过向澄清的裂解物中添加苯并酶、1mtrisph7.5和5mnacl来纯化和浓缩用于质谱分析的载体,使其最终浓度为20mmtris和360mmnacl。在1mlporoscaptureselect亲和柱上捕获载体,并在低ph下将载体峰直接洗脱到中和缓冲液中。通过在280nm处的吸光度分析级分,并且使最浓缩的级分经受质谱分析。对于体内实验,产生了如先前所描述的具有wtaav8衣壳或具有6个脱酰胺化突变体之一的载体;转基因盒包含cb7启动子、pi内含子、萤火虫荧光素酶转基因和rbg聚腺苷酸化信号(lockm等人,《人类基因疗法》2010;21(10):1259-71)。3.质谱运行/消化/分析材料:从西格玛公司(密苏里州圣路易斯市)购买了碳酸氢铵、dtt、碘乙酰胺(iam)和富集18o的水(纯度为97.1%);并且从赛默飞世尔科技公司(伊利诺伊州罗克福德市)购买了乙腈、甲酸、三氟乙酸(tfa)、8m盐酸胍(gndhcl)和胰蛋白酶。胰蛋白酶消化:制备了1mdtt和1.0m碘乙酰胺的储备溶液。使衣壳蛋白变性,并在存在10mmdtt和2mgndhcl的情况下在90℃下还原十分钟。允许样品冷却到室温,并且然后在黑暗中在室温下用30mmiam使其烷基化30分钟。通过添加1mldtt淬灭烷基化反应。将20mm碳酸氢铵(ph7.5-8)以将最终的gndhcl浓度稀释到200mm的一定体积添加到变性的蛋白质溶液中。添加胰蛋白酶溶液,使胰蛋白酶与蛋白质的比率达到1:20,并在37℃下温育过夜。在消化之后,将tfa添加到最终浓度0.5%,以淬灭消化反应。对于18o水实验,首先使用zeba旋转脱盐柱(伊利诺伊州罗克福德市赛默飞世尔科技公司)将衣壳样品缓冲液交换成在18o水中制备的100mm碳酸氢铵。为了确保完全去除样品中的水,进行了两次缓冲液交换。在18o水中制备了1mdtt和1miam的储备溶液。使用18o水试剂和缓冲液进行与上述相同的变性、烷基化和消化步骤。液相色谱串联-质谱:使用acclaimpepmap柱(长15cm,内径300μm)和与具有nanoflex源的qexactivehf(赛默飞世尔科技公司)联接的thermoultimate3000rslc系统(赛默飞世尔科技公司)进行在线色谱。在线分析期间,柱温保持在35℃下。用流动相a(含0.1%甲酸的milliq水)和流动相b(含0.1%甲酸的乙腈)的梯度分离肽。梯度在15分钟内从4%b运行到6%b,在25分钟内运行到10%b(共40分钟),并且然后在46分钟内运行到30%b(共86分钟)。将样品直接上样到柱上。柱尺寸为75cm×15umi.d.,并装有2微米的c18培养基(acclaimpepmap)。由于上样、导入和洗涤步骤,每个液相色谱串联-质谱的总时间为约两小时。在qexactivehf质谱仪上使用数据依赖性前20种方法获取质谱数据,所述质谱仪从调查扫描(200-2000m/z)中动态选择最丰富的尚未测序的前体离子。通过高能碰撞解离片段化进行测序,其中通过预测性自动增益控制确定的靶值为1e5离子;并且以4m/z的窗口进行前体分离。在200m/z下以120,000的分辨率获取调查扫描。hcd光谱的分辨率在m/z200下设置为30,000,其中最大离子注入时间为50毫秒,并且归一化碰撞能量为30。s-透镜rf水平设置为50,以使来自消化的肽所占据的m/z区达到最佳传输。从片段化选择中排除具有单个、未分配或六个和更高电荷状态的前体离子。数据处理:使用biopharmafinder1.0软件(赛默飞世尔科技公司)分析所获取的所有数据。对于肽作图,使用单进入蛋白fasta数据库进行搜索,其中脲基甲基化设置为固定的修饰,并将氧化、脱酰胺化和磷酸化设置为可变修饰。对于串联质谱,使用了10ppm的质量准确度、高蛋白酶特异性和0.8的置信水平。脱酰胺化的肽的质谱鉴定相对简单,因为脱酰胺化向完整分子的质量添加了+0.984da(-oh基团与-nh2基团之间的质量差)。通过将脱酰胺化的肽的质量面积除以脱酰胺化的和天然的肽的面积之和来确定特定肽的脱酰胺化百分比。考虑到可能的脱酰胺化位点的数量,在不同位点处被脱酰胺化的同量异位物种可以在单个峰处共迁移。因此,源自具有多个潜在的脱酰胺化位点的肽的片段离子可以用于定位或区分多个脱酰胺化位点。在这些情况下,观察到的同位素图案内的相对强度可以用于特异性确定不同的脱酰胺化的肽异构体的相对丰度。此方法假设所有异构物种的片段化效率是相同的,并且脱酰胺化位点是独立的。此方法允许定义脱酰胺化涉及的特定位点以及脱酰胺化涉及的潜在组合。次级数据处理:使用以下方法在马里兰州巴尔的摩市的大学进行了原始质谱的次级分析。所有质谱分析均使用peaksstudiov5.3软件(生物信息学解决方案有限公司)。使用以下参数对原始数据文件进行数据精化:前体的m/z公差≤10ppm,并且前体电荷状态最小为2,最大为4。使用峰算法对输入光谱进行从头测序,其中前体离子误差公差为10ppm,并且产物离子误差公差为0.1da。将消化酶设置为胰蛋白酶,可变修饰设置为氧化、磷酸化和脱酰胺化,并且固定修饰设置为半胱氨酸的脲基甲基化。4.aav衣壳的结构分析从rcsb蛋白质数据库(pdbid:3ra8)获得了aav8原子坐标、结构因子和相关的衣壳模型。进行了结构精修,并产生了独立于aav8vp3的初级氨基酸序列的电子密度,以用于衣壳的三维(3d)结构分析。执行此分析以观察aav8衣壳中的不受预期的aav8vp3的初级序列偏置的异天冬氨酸电子密度。使用所得结构,用n+1个甘氨酸作为异天冬氨酸对aav8vp3初级序列中的四个天冬酰胺进行建模,并且然后使用crystallography和nmr系统(cns)软件通过使用标准精修方案严格施加二十面体非结晶矩阵来精修aav8衣壳结构(brungerat等人,《晶体学学报d区—生物晶体学(actacrystallogrdbiolcrystallogr)》1998;54(pt5):905-21)。从hic-up数据库中获得异天冬氨酸的结构模型,然后在prodrg中生成分子词典以进行结构精修(kleywegtgj,《晶体学学报d区—生物晶体学》2007;63(pt1):94-100)。然后,计算aav8衣壳的平均电子密度图(也在cns中),并使用coot软件使其可视化,然后对所得模型进行较小的调整,以将建模的异天冬氨酸残基拟合到电子密度图中(emsleyp和cowtank,《晶体学学报d区—生物晶体学》2004;60(pt12pt1):2126-32)。重复此方案,以使用n+1个甘氨酸(pdbid:3ux1)在aav9vp3初级序列中另外对n512进行建模。使用coot、pymol和ucsfchimera生成了所有图(emsleyp和cowtank等人,《晶体学学报d区—生物晶体学》2004;60(pt12pt1):2126-32;delanowlpymol:开源分子图形工具(delanowlpymol:anopen-sourcemoleculargraphicstool)》第40卷,2002:82-92;pettersenef等人,《计算化学杂志(jcomputchem)》2004;25(13):1605-12)。获得了多种先前鉴定的脱酰胺化的蛋白质(pdbid:1dy5、4e7g、1rtu、1w9v、4e7d和1c9d)的结构,以用于将其脱酰胺化的异天冬氨酸残基的电子密度图与来自aav8和aav9的建模的异天冬氨酸残基进行比较(raofv等人,《化学生物(chembiol)》2005;12(1):65-76;noguchis等人,《生物化学》1995;34(47):15583-91;elliott等人,《分子医学杂志(jmolbiol)》2000;297(3):713-32)。通过求aav8或aav9晶体结构原子坐标(pdbid:3ra8、3ux1)中报告的每个天冬酰胺残基的每个原子的温度系数的平均值来确定脱酰胺化的残基的温度系数。5.动物研究宾夕法尼亚大学的机构动物护理和使用委员会批准了所有动物程序。为了评估载体性能,通过尾静脉注射向八周龄的c57bl/6小鼠静脉内注射3e10gcwtaav8或衣壳突变载体,体积为100μl。在第14天处死所有小鼠。为了体内评估荧光素酶表达,将小鼠(约20g)麻醉并腹膜内注射200μl或15mg/ml萤光素底物(马萨诸塞州沃尔瑟姆市珀金埃尔默公司(perkinelmer,waltham,ma))。在萤光素施用之后五分钟对小鼠进行成像,并通过ivisxenogen体内成像系统进行成像。使用livingimage3.0软件对所描述的所关注的区中的信号进行定量。在第7天和第14天进行测量。6.评估突变载体效价和体外转导效率通过抗dnasei基因组的qpcr确定载体效价。使qpcr引物与包装的转基因的聚腺苷酸化序列粘接。为了通过荧光素酶表达体外评估载体转导效率,将0.9e5huh7细胞/孔接种在完整dmem(10%胎牛血清,1%青霉素/链霉素)中的黑壁96孔板中。第二天,去除培养基,并用在完全培养基中稀释的50μl粗制或经过纯化的载体将其替代。对于每种粗载体样品,以3倍稀释系列测试4种稀释液。在48小时之后,以0.3μg/μl在完全培养基中制备荧光素(威斯康星州麦迪逊市普洛麦格公司),并以50μl的体积将其添加到经过转导的细胞中。在biotekclarity光度计上读取结果。发现添加到靶细胞的荧光素酶活性/gc在宽gc范围内是恒定的,但在高moi时可能变得饱和。因此,检查了针对线性度的稀释系列数据(发光单位对比gc),如果饱和度明显,则排除最高点,并针对每种变体的每个测定的线性范围中的值计算平均荧光素酶/gc。这产生转导效率值。通过将wt对照物的值设置为1,对数据进行归一化以简化比较。7.生物分布使用qiaampdna迷你试剂盒(德国希尔登凯杰公司)从肝脏样品中提取dna,并且然后使用针对转基因盒的rbg聚腺苷酸化信号设计的引物/探针组如先前所描述的通过实时pcr分析用于载体gc的dna(chensj等人,《人类基因疗法临床开发(humgenetherclindev)》2013;24(4):154-60)。aav8突变体的引物序列b.结果aav8示出其衣壳蛋白中的大量电荷异质性为了定量评估可能影响载体性能的aav8载体衣壳上的转译后修饰的存在,通过1d和2d凝胶电泳两者分析了通过碘克沙醇梯度纯化的aav8总衣壳蛋白。在1d还原十二烷基硫酸钠sds凝胶中,vp1、vp2和vp3以单个带的形式在合适的分子量下解析(图1b)(roseja等人,《病毒学杂志》1971;8(5):766-70)。当通过基于电荷分离蛋白质(图1c)的2d凝胶电泳进一步评估时,取决于vp同种型,衣壳蛋白中的每种衣壳蛋白另外解析为一系列具有不同等电点(pi)的不同点,其范围从ph6.3到>7.0(图1d)。每个vp上的单独点通过如所测量的作为相对于碳酸酐酶同种型内部等电点标准的迁移的0.1pi单位的离散间隔分开,这表明单个残基电荷发生变化。这些同种型的存在表明每个vp都有可能经历许多修饰,由此导致其在等电聚焦下不同地迁移。其中一部分(通常为天冬酰胺)侧链酰胺基团转化为羧酸的脱酰胺化(图1a)是蛋白质制剂中电荷异质性的常见来源。为了确定脱酰胺化是否可能负责vp电荷同种型的不同群体,将两个aav8天冬酰胺残基分别突变为天冬氨酸。这些衣壳突变应使电荷偏移,其偏移量等于使单个另外的天冬酰胺残基完全脱酰胺化的量。突变体的2d凝胶分析表明,与野生型(wt)aav8中的等效点相比,vp1、vp2和vp3的主要点偏移更酸性的一个点位置(0.1ph单位)(图1e-图1g)。此偏移的幅度等于wtvp电荷同种型之间观察到的间隔。因此,aav衣壳蛋白的2d凝胶图案化与多位点脱酰胺化一致。在aav8载体衣壳上发生自发脱酰胺化为了鉴定负责每个衣壳蛋白的离散点样图案的修饰,通过质谱分析一组aav8载体。aav8衣壳蛋白的平均覆盖>总vp1序列的95%(数据未示出)。通过质谱检测到天冬酰胺和谷氨酰胺残基的子集的广泛脱酰胺化,这示出与基于由dna编码序列的预测值相比,观察到的单独肽质量增加了约1da;在aav8载体的所有制剂中都观察到脱酰胺化的此模式(图2a-图2d)。为了评估常用纯化方法之间脱酰胺化的总体异质性并检查vp1和vp2独特区中的脱酰胺化,选择了通过三重转染在293个细胞中产生的九批aav8,并通过氯化铯梯度、碘克沙醇梯度或亲和色谱对其进行纯化。载体在启动子和转基因盒方面也不同。为了确定载体基因组的存在是否对脱酰胺化有影响,还评估了在不存在顺式质粒(仅产生空衣壳)的情况下通过三重转染在293个细胞中产生的aav8制剂,并通过碘克沙醇梯度进行纯化。跨aav8衣壳的天冬酰胺和谷氨酰胺残基存在广泛范围的脱酰胺化,范围从不可检测到超过99%的被脱酰胺化的单独氨基酸(图2e)。在天冬酰胺残基处发生最高水平的脱酰胺化(>75%),其中n+1残基为甘氨酸(即,ng对)(表1)。在n+1不是甘氨酸的另外的天冬酰胺残基处检测到较低水平的脱酰胺化(即,最高17%)。制剂之间的天冬酰胺的平均脱酰胺化在很大程度上一致。还检测到在谷氨酰胺残基处的脱酰胺化,但频率比在天冬酰胺处的频率低;在q467处观察到的最高百分比<2%(图7)。此观察结果在所有制剂中均不一致(数据未示出)。在残基n499(n+1残基是天冬酰胺)处观察到最大的制备间差异,其值范围为<1%到50%以上的脱酰胺化。无论如何,在载体制剂之间的脱酰胺化中观察到的变化似乎与纯化方法、转基因同一性或载体基因组的存在不相关,这表明这些因素不会影响脱酰胺化率。表1:所关注的aav8脱酰胺化的残基的特性。星号表示所选的用于进一步分析的残基。接下来,进行了一系列实验,以确定样品处理是否有助于aav8中的观察到的脱酰胺化水平。极端温度(70℃持续7天)或ph(ph2或ph10持续7天)没有显著诱导aav8衣壳中的另外的脱酰胺化(图4a和图4b)。考虑到这种抗性,认为观察到的脱酰胺化不太可能仅发生在相对较短且较温和的纯化阶段。试图对未经纯化的载体进行质谱分析,以确定纯化前后的脱酰胺化程度,但均未成功。同样,重水对照物表明特定于质谱工作流程的处理不会有助于另外的脱酰胺化事件(图4c)。为了验证质谱工作流程,检查了两种先前已经评估脱酰胺化的重组蛋白;所述发现(图5a和图5b)与已经公开的结果一致[henderson,le、henriksson,d和nyman,po(1976),“人碳酸酐酶c的初级结构(primarystructureofhumancarbonicanhydrasec)”《生物化学杂志》251:5457-5463以及carvalho,rn、solstad,t、bjorgo,e、barroso,jf和flatmark,t(2003),重组人苯丙氨酸羟化酶中的脱酰胺化(deamidationsinrecombinanthumanphenylalaninehydroxylase)不稳定的天冬酰胺残基的鉴定和asn-->asp突变体形式的功能表征(identificationoflabileasparagineresiduesandfunctionalcharacterizationofasn-->aspmutantforms)《生物化学杂志》278:15142-1515]。另外,雇用了次级机构来评估来自aav8的原始数据。此独立分析鉴定了与被脱酰胺化的位点相同的位点,其中在每个位点处的修饰程度变化最小,这归因于峰检测和面积计算中软件间的变化(图6)。n+1氨基酸的结构拓扑、温度系数和同一性有助于脱酰胺化频率由于aav8的结构已经解决并公开(pdb标识符:2qa0)(namhj等人,《病毒学杂志》2011;85(22):11791-99),接下来检查aav8衣壳结构,以为非酶促脱酰胺化的有利条件提供证据,并使脱酰胺化百分比与已建立的结构特征相关(namhj等人,《病毒学杂志》2007;81(22):12260-71)。仅关注天冬酰胺残基,因为影响天冬酰胺脱酰胺化的因素在文献中被更好地表征,并且天冬酰胺脱酰胺化事件远比谷氨酰胺脱酰胺化事件更为常见(robinson,ne和robinson,ab(2001),“分子时钟(molecularclocks)”《美国国家科学院院刊》98:944-949)。还确定了来自aav8晶体结构的这些残基中的每个残基的温度(或b)系数;温度系数是对原子从其平均位置开始的位移的量度,值越大表示位移越大、热振动越大,并且因此柔性越高(parthasarathys和murphymr,《蛋白质科学:蛋白质协会的出版物(proteinscience:apublicationoftheproteinsociety)》1997;6:2561-7))。大多数所关注的天冬酰胺都定位在表面暴露的hvr中或其附近(表1),这在结构上有利于脱酰胺化,并提供了溶剂暴露的环境(govindasamyl等人,《病毒学杂志》2013;87(20):11187-99)。发现平均而言定位在这些柔性环区中的残基比如β链和α螺旋等柔性较差的区中的残基更频繁地被脱酰胺。例如,在位置n263处的ng残基是hvri的一部分,具有高温系数,并且平均被脱酰胺化>98%(图7a和图6,表1)。在所述时间内被脱酰胺化约85%的n514(图3和图6,表1)也存在于具有n+1甘氨酸的hvr(hvrv)中;然而,局部温度系数与n263的温度系数相比相对低,这是由于其与在三重轴处的其它vp单体上的残基相互作用。不利的+1残基和较低的局部温度系数与较低的脱酰胺化相关,即使对于hvr残基也是如此。例如,n517平均仅4%脱酰胺化(表1);此残基具有与高度脱酰胺化的n514相当的温度系数,但其n+1残基为丝氨酸,这降低了由于空间位阻而引起的脱酰胺化事件的可能性。这表明,尽管+1残基的同一性显然是最有影响力的因素,但许多因素会累积地确定在给定衣壳位置处的脱酰胺化程度。为了测试+1残基在天冬酰胺脱酰胺化中的作用,产生了突变载体,其中aav8ng位点在+1位置处分别突变为丙氨酸或丝氨酸。模型肽研究表明,ng肽的脱酰胺化半衰期短至1天,而na肽或ns肽的脱酰胺化通常分别慢25倍或16倍(robinsonne和robinsonab,《美国国家科学院院刊》2001;98(8):4367-72)。载体突变体的质谱分析证实了+1位点在确定载体脱酰胺化程度中起着核心作用。当+1位点变为丙氨酸(<5%脱酰胺化)或丝氨酸(<14%脱酰胺化)时,此集合中的ng位点(>80%脱酰胺化,在wt中)示出相邻天冬酰胺的选择性稳定(表2)。表2:在wt中五个aav8ng位点处和六个+1位点突变体的脱酰胺化程度(%)位置/变体wt(平均值)g58sg58ag264ag386sg386ag515an5781.88.41.989.789.791.693.6n26399.398.298.94.8100.094.597.2n38589.196.394.897.113.52.597.0n51485.2100.098.098.8100.0100.02.2n54084.595.092.697.996.986.189.5与定位在更有利的环境中的残基相比,在完整的完全组装的aav8衣壳中,至少部分被掩埋且不易暴露于溶剂和/或定位在局部柔性低的区中的残基的脱酰胺化频率更低(表1)。尽管如此,在不利条件下的少数残基仍被脱酰胺化。例如,n630被至少部分掩埋,但仍具有可检测的脱酰胺化程度。对于此残基,苯丙氨酸作为n+1残基存在表明此区可能是aav8vp3蛋白内的非酶促自蛋白水解裂解的新颖位点。aav8vp3的结构建模证实了脱酰胺化事件为了提供在组装的衣壳的背景下的脱酰胺化的直接证据,评估了aav8的晶体结构(namh-j等人,《病毒学杂志》2011;85(22):11791-9)。此血清型的可用晶体结构(即,)的分辨率不足以鉴定r基团中的末端原子,并且因此不足以在天冬酰胺残基、天冬氨酸残基与异天冬氨酸残基之间进行直接区分。在这些条件下形成的天冬氨酸的异构体的结构的其它方面提供了从结构确定脱酰胺化的机会。此分析基于两个假设:1)天冬酰胺的自发脱酰胺化的主要产物是异天冬氨酸而不是天冬氨酸,其以3:1的比率产生(geigert和clarkes,《生物化学杂志》1987;262(2):785-94);以及2)由于对应于异天冬氨酸的r基团的电子密度图长度较短,因此可以将天冬酰胺或天冬氨酸与异天冬氨酸区分开。在脱酰胺化反应期间解析琥珀酰亚胺基中间体之后,当将来自异天冬氨酸的r基团的β碳并入到aav8vp3衣壳蛋白骨架的主链中而丢失时,这会产生较短的r基团。首先精修了aav8结构本身,从而产生不受已知aav8vp3序列偏置的aav8衣壳电子密度。然后,基于与异天冬氨酸相关的较短的r基团的存在,检查了经过精修的aav8晶体结构以证明脱酰胺化(图3a-图3e)。电子密度图证实,与在410处通过质谱没有检测到脱酰胺化的天冬酰胺相比(图3b),在位置263处(图3c)、385处(未示出)、514处(图3d)和540处(图3e)高度脱酰胺化的n+1甘氨酸残基的r基团更短。因此,由电子密度图指示的脱酰胺化与通过质谱在这些位点处进行的>75%脱酰胺化的数据一致。所得异天冬氨酸模型与在其它已知的脱酰胺化的蛋白质的晶体结构中观察到的异天冬氨酸残基相当,这支持aav8的分析的有效性(raofv等人,《化学生物》2005;12(1):65-76;noguchis等人,《生物化学》1995;34(47):15583-91;elliott等人,《分子医学杂志》2000;297(3):713-32)。此结构分析用作对通过质谱分析aav8衣壳时观察到的脱酰胺化现象的独立证实。aav衣壳的脱酰胺化不是血清型特异性的调查了aav8以外的血清型,以证明衣壳脱酰胺化。使用2d凝胶电泳(图11a)和质谱(图11b)检查aav9载体制剂,包含针对潜在载体处理效果的对照物(图11d-图11f)。aav9脱酰胺化的模式和程度与aav8脱酰胺化的模式和程度类似。所有四个aav9ng位点>85%脱酰胺化;13个非ng位点的脱酰胺化程度较小,其中少数位点示出的脱酰胺化%的批次间变异性很大。接下来,应用了结构分析工作流程,并对现有的aav9晶体学数据进行了重新拟合(图11c,表3)。与aav8一样,异天冬氨酸更好地拟合到aav9晶体结构中若干个ng位点的电子密度。将2d凝胶分析(数据未示出)和质谱(总结在表4中)扩展到了五种另外的进化上不同的血清型(rh32.33、aav7、aav5、aav4、aav3b和aav1)。所有检查的衣壳均含有类似的脱酰胺化模式和程度,这表明此修饰在临床上相关的aav载体中是广泛的,并由类似的基础初级序列和结构因素确定。表3.所关注的aav9脱酰胺的残基的特性。具有同源n+1残基(与aav8相比)的保守天冬酰胺残基以斜体表示(由aav8和aav9vp1的全长氨基酸序列的比对确定)。表4.观察到不同血清型的脱酰胺化程度脱酰胺化事件可能影响衣壳组装和转导效率测试脱酰胺化的功能影响的一种方法是通过基因突变用天冬氨酸取代天冬酰胺。通过对293个细胞进行小规模三重转染为每个脱酰胺化的aav8天冬酰胺产生对荧光素酶报告基因进行编码的天冬氨酸突变载体,并通过抗dnasei基因组拷贝的qpcr对载体进行滴定(图8a)。相对于wtaav8,突变极少影响衣壳组装,并且影响仅限于wt载体中总体上具有低脱酰胺化的大部分掩埋的非ng位点。接下来,评估人肝源性huh7细胞的突变组体外转导效率(图8b)。若干个突变体示出受损的转导效率,其中位置n57、n94、n263、n305、q467、n479和n653表现出>10倍的转导损失。观察到类似数量的aav9的敏感位点(图11g和图11h)。通常,在给定位置处只有一部分残基是内源性脱酰胺化的,因此此方法可能会高估蛋白质(如衣壳)的功能丧失,其中功能单元是同单体组装;在一个衣壳位点处的内源性修饰可以被具有完整残基的相邻亚基补偿。尽管如此,认为所述方法可以帮助脱酰胺化的残基进行优先级排序,以便在制备或突变稳定期间进行进一步监测。将需要来自内源性脱酰胺化载体的群体的功能数据,以将这种功能丧失诱变数据置于适当的背景下。随着时间的推移的载体活性丧失与进行性脱酰胺化相关考虑到ng脱酰胺化的半衰期很短,认为年龄相差仅1天的载体样品可能示出不同的脱酰胺化谱,从而提供使内源性脱酰胺化与功能相关的机会。大规模载体制备方案要求对293个细胞进行三重转染,然后温育5天以产生载体,并需要1-2天进行载体纯化。为了近似此过程,用wtaav8制备293个细胞的中等规模三重转染(每个细胞培养皿10×15cm)。以1天的间隔收集载体(2×15cm细胞培养皿/天)持续5天,从而通过将载体冷冻在-80℃下保存到5天结束的时间点。接下来,如上文所描述的评估粗载体效价和体外转导效率。如所期望的,组装的抗dnasei基因组拷贝的数量随时间推移而增加(图9a)。然后,通过亲和纯化快速处理早期(第1天和第2天)和晚期(第5天)时间点的粗载体,并测量huh7细胞的体外转导效率。载体的相对转导效率随时间推移而逐渐降低(图9b)。就添加到靶细胞的每个gc的转基因表达而言,第5天的载体的效率为第1天的材料的效率的仅40%。也观察到粗材料的此活性下降,这表明纯化前分子组合物的变化(图)。观察到5天内aav9的活性丧失趋势类似,其中载体效力降低了约40%(图11i-11k)。接下来,通过质谱测量时程样品的脱酰胺化。ng位点脱酰胺化基本上在每个间隔内进行,其中在第1天平均25%脱酰胺化,并且到第5天转化的位点>60%(图9c)。非ng位点脱酰胺化通常在5天内进行,尽管在第2天与第5天之间的水平要低得多并且一致性较低(图9d)。数据使内源性载体脱酰胺化与特异性活性的早期时间点衰减相关,并通过缩短生产周期或寻找稳定天冬酰胺的衣壳突变突出显示捕获更多活性载体的潜在机会。应注意的是,图2a-图2e中的用于质谱分析的材料是转染后至少7天的,这是由于另外2天用于纯化。这些样品中的较高ng位点脱酰胺化(>80%)表明,脱酰胺化可能会在表达时间段之后以及在回收和纯化过程期间以约相同的速率继续进行,直到ng位点被完全脱酰胺化或载体样品被冷冻。因此,脱酰胺化很大程度上由载体的年龄确定,而不是回收和纯化过程所独有或引起的过程。第1天的材料对比第5天的材料(均经过亲和纯化)的低得多的脱酰胺化值说明了这一点。稳定ng天冬酰胺可以改善载体性能考虑到载体ng脱酰胺化与转导效率损失之间的相关性,认为通过+1位点诱变稳定ng酰胺可以改善载体功能。以小规模为aav8ng位点突变体产生载体,其中每个+1残基分别转化为丙氨酸或丝氨酸。就载体组装(图10a)和转导效率(图10b)而言,单个+1突变体具有良好的耐受性。定位在衣壳表面上的先前定义的“死区”附近的g386取代(aydemirf等人,《病毒学杂志》2016年7月;90(16):7196-204)存在体外转导缺陷。g386突变体的功能丧失可能表明对在n385处的脱酰胺化的天冬酰胺的偏好。可替代地,+1位置处的另外的侧链本体可能对独立于酰胺基团稳定化的功能具有负面影响。尽管其相邻的天冬酰胺具有显著的稳定性,但单个位点突变体没有显著改善体外转导(表2)。因为体外和体内的转导活性可能不一致,所以在c57bl/6小鼠中测试了用于肝脏转导的单个位点+1突变体的子集。进行静脉内尾静脉注射(n=3到5),并通过持续2周的每周成像来检查荧光素酶表达(图10c)。体内和体外转导数据与每个测定的相关误差范围相一致(即,在误差范围内)。g386取代在转导方面是有缺陷的,而在其它位置处的+1位点突变在很大程度上被耐受,从而以等于但不超过wtaav8的水平转导肝脏。由于在任何一个ng位点处稳定酰胺可能是必要的,但不足以恢复功能,因此接下来评估了具有+1位点丙氨酸取代的组合的载体变体。重组+1丙氨酸功能强大的所有3个aav8ng位点(n263、n514和n540)。一些包含三重突变体g264a/g515a/g541a的组合组装不佳,并且对转导而言具有功能障碍。然而,涉及n263的两种成对组合(g246a/g515a和g264a/g541a)均改善了体外转导效率(分别是wtaav8的2.0倍和2.6倍),而没有效价损失(图10d)。因为这些突变会引入至少两种变化(n-酰胺稳定化和+1残基侧链取代),所以这些数据不能最终将ng脱酰胺化与功能丧失联系起来。然而,数据与在时程研究中建立的模型一致,在所述模型中,ng位点脱酰胺化可能影响体外转导效率。功能性天冬酰胺取代改进了载体制备的批次间重现性报告的载体脱酰胺化谱的另一个潜在问题方面是在一些位置处的脱酰胺化具有高批次间变异性。对于wtaav8,此变异性对于n459(观察到的脱酰胺化范围为0%到31%)和n499(观察到的脱酰胺化范围为0%到53%)最为显著。通常在生物制剂开发期间实际上要避免转译后修饰中的变异性,这通过完全避免表现出此变异性的克隆,仔细监测和控制生产菌株和条件,或者对受影响的候选物进行蛋白质工程化。由于无法确定导致n459和n499脱酰胺化变异性的生产或加工因素(图2e),因此寻求在这些位置处进行功能性氨基酸取代。首先评估了小规模载体制剂在各个位置处分别被谷氨酰胺保守取代的可能性。n459q和n499q两者都高效地组装到载体中,并且与wtaav8的体外转导效率参考相当(图7a)。接下来,大规模产生突变体并进行质谱。与对极为罕见的谷氨酰胺脱酰胺化的观察一致,观察到这些突变体中在位置459或499处的谷氨酰胺具有选择性和完全的稳定性(数据未示出)。如上所述在c57bl/6小鼠的尾静脉注射后体内评估了这些突变体批次的肝脏转导(图7b和图7c)。在此实验中用作对照物的wtaav8载体批次在n499处脱酰胺化16.8%,但是在n459处未检测到脱酰胺化(数据未示出)。两个突变体在第14天的肝脏转导等同于wtaav8的肝脏转导。此数据证明了蛋白质工程化方法解决制备的aav载体中与脱酰胺化相关的分子变异性的潜力。c.讨论分别通过2d凝胶电泳、质谱、从头蛋白质建模以及体外和体内两者的功能研究来鉴定和评估aav8衣壳上天冬酰胺和谷氨酰胺残基的非酶促脱酰胺化。已经示出脱酰胺化发生在广泛多种蛋白质中,并显著影响生物制剂的活性,包含基于抗体的治疗剂(nebijad等人,《国际分子科学学报(intjmolsci)》2014;15(4):6399-411)和基于肽的疫苗(vermaa等人,《临床与疫苗免疫学》2016;23(5):396-402)。已经通过质谱示出其它病毒蛋白(如轮状病毒的vp6蛋白)会经历脱酰胺化事件(emsliekr等人,《功能与整合基因组学(functintegrgenomics)》2000;1(1):12-24)。在aav8中发生这些脱酰胺化的情况表明它们是自发的非酶促事件的结果。已知天冬酰胺残基比谷氨酰胺残基更广泛地被脱酰胺化;天冬酰胺下游的氨基酸基本上影响脱酰胺化的速率,其中n+1甘氨酸(即,ng)被最高效地脱酰胺化。观察到n+1氨基酸在aav衣壳的脱酰胺化中的作用得到明显证实,因为vp1中存在的每个ng均以>75%的水平被脱酰胺化,而衣壳中的其它天冬酰胺或谷氨酰胺中的任一种的脱酰胺化始终从未>20%。实际上,aav8和aav9衣壳(即,7/9)中的所有ng基序也存在于与构象柔性和热振动的高速率相关的hvr区中含有的衣壳的表面上。这与其它蛋白质的ng基序的先前报告是一致的,所述其它蛋白质定位在可能需要柔性来实现适当蛋白质功能的区中,而不是在更有序的结构中,如α螺旋或β折叠(yanbx和sunyq,《生物化学杂志》1997;272(6):3190-4)。通过提供溶剂可及性和构象柔性,对表面暴露的hvr中ng基序的偏好进一步增强了脱酰胺化速率,由此促进了琥珀酰亚胺基中间体的形成。如所预期的,不利的环境导致脱酰胺化速率低得多。关于aav的生物学及其作为载体的用途的重要问题是这些脱酰胺化的功能后果。将天冬酰胺转化为天冬氨酸的衣壳dna的诱变允许评估衣壳,其中在特定位点处的所有氨基酸均表示为天冬氨酸。然而,除潜在地突变n+1残基之外,不存在使用诱变防止脱酰胺化的任何容易的策略,这与第二位点突变的直接后果相混淆。研究了有限数量的变体,其中通过诱变将天冬酰胺残基转化为天冬氨酸。功能分析包含衣壳组装以及体外和体内转导。诱变对载体功能的最基本影响是涉及在基线处没有被完全地脱酰胺化并且没有暴露的表面的天冬酰胺的影响。然而,令人惊讶的是,在514处将高度脱酰胺化的天冬酰胺诱变为天冬氨酸确实对功能有一些影响。此结果表明对应酰胺的残留量的存在可能影响功能。这可能部分地由于另一三倍相关的vp3单体(在wtaav8晶体结构中鉴定)的n514与d531之间存在氢键相互作用,所述氢键相互作用在脱酰胺化之后此残基转化为天冬氨酸时失去。在评估这些脱酰胺化对新颖治疗剂开发的影响时,更好地了解影响aav载体中脱酰胺化程度的因素很重要。在极端条件(已知能显著加速脱酰胺化动力学)下温育载体几乎没有影响。结合同位素并入研究,此结果表明脱酰胺化发生在衣壳组装期间,并且不是载体处理或质谱分析的伪像。在ng位点处的脱酰胺化不太可能对载体性能具有实质性影响,因为在评估的每个样品中,反应实际上都已经完成。然而,最初的功能研究表明未脱酰胺化的天冬酰胺的残留量可以有助于功能。更加关注其中脱酰胺化不完全的位点,在大多数情况下,这也与样品间的差异相关。实例是在位置499处的天冬酰胺示出的脱酰胺化范围为0%到53%,平均为17%。载体产生条件的细微差异可能会导致这种异质性。aav8和aav9中脱酰胺化的惊人相似性表明这是这种整个病毒家族的性质。总之,在aav8和aav9衣壳蛋白的初级氨基酸结构中发现了大量异质性。这些研究可能以若干种方式影响作为载体的aav的发展。首先,vp蛋白的实际氨基酸序列不是由对应dna序列所预测的。其次,生产方法的各个方面可能导致脱酰胺化的差异以及载体功能的对应变化。直到掌握影响在非ng位点处的脱酰胺化速率的因素并更好地了解其功能后果之前,可能有必要在临床级aav载体的表征中包含脱酰胺化。2d凝胶电泳可以提供净脱酰胺化的总体评估,尽管质谱对于评估在特定残基处的脱酰胺化是必不可少的。实例2:脱酰胺化aav5.5.9seqidno:9和10中分别提供了aav5.5.9的新颖序列。如实例1中针对aav9所描述的,评估aav5.5.9载体的脱酰胺化。在n57、n319、n442、n502处可以看到高度脱酰胺化的残基。实例3:脱酰胺化aavrh79(进化枝e)aavrh79是从恒河猴的小肠组织提取的dna中分离出的。其在系统发育上已经被表征为在进化枝e内(图14a-14d)。本文提供了其序列,其中核苷酸序列在seqidno:1中,并且氨基酸序列在seqidno:2中。图14a中提供了aavrh79、aavrh.10和aavhu.37的氨基酸序列的比对。图14b-14d中提供了aavrh79、aavrh.10和aavhu.37的核酸序列的比对。aavrh79在其初级序列中具有三种氨基酸差异。而aavhu37具有定位在其初级vp1序列的位置67处的ala和在位置169处的lys,aavrh79具有在位置67处的谷氨酸(e)和在位置169处的arg。图11b中示出了rh.79、hu.37和hu.40之间的vp1的dna序列的差异。制备基于各种进化枝e变体的载体表达性egfp,并评估其对huh7细胞的相对感染性(图11c)。c57bl/6小鼠注射有两个剂量水平(3×1010和3×1011gc/小鼠)的表达egfp的aav8或aavrh.79载体,并且通过荧光显微镜评估感染性(数据未示出)。使用已知的产生技术使用用于产生cap的aavrh79核苷酸序列来制备基于aavrh79的载体,如先前针对aav8载体所描述的。图15a-15b和图16中分别提供了产生产率和产生纯度评估的结果。为了使用含有标志物基因(萤火虫荧光素酶)的aavrh79评估表达水平,使用汉密尔顿注射器向6-8周龄的雄性ragko小鼠肌内注射3×1011gc/小鼠的载体。如先前所描述的,通过全身生物发光成像使ffluc表达可视化[greigja、pengh、ohlsteinj、medina-jaszekca、ahonkhaio、mentzingera等人,(2014),“小鼠和猕猴肌内注射aav8与大量肝靶向和转基因表达相关(intramuscularinjectionofaav8inmiceandmacaquesisassociatedwithsubstantialhepatictargetingandtransgeneexpression)”公共科学图书馆综合(plosone)9(11):e112268.doi.org/-10.1371/journal.pone.-0112268.]图17a-17d中提供了结果。在向雄性和雌性食蟹猴肌内施用1013gc/kgaavrh79后,比较avv8三重、aavhu68、aav9、aav8和aavrh79载体的表达(图17e)。将表达分泌的转基因(201igia)的载体肌内注射到雄性ragko小鼠(n=5/组)的腓肠肌中(3×1010或3×1011gc/小鼠)。结果表明,im注射后,aav8三重表达更好,并且在较低的测试剂量下,来自aav8三重的表达的差异相当大。在较高的较高剂量下,avvrh79以与所测试的其它载体相当的水平表达(图17f)。预筛选雌性食蟹猴(ra2362)的nab(图18a),并注射aavg2.tbg.egfp.wpre.bgh(1×1013ddgc/kg,静脉内)。处理后7天对动物实施安乐死,并进行尸检以分离肝脏和其它组织以供分析。在第7天评估肝脏和脾中的gfp表达(图18b和18c)。在图18d中示出在各种组织中检测到的载体水平。在接受aavg2(ra2362)或aav8和aavg3载体的动物的肝脏中评估gfp表达水平(图18e-图ij)。图18k示出了在来自这些动物的各种组织中检测到的载体水平。产生各种aav8和aavrh79载体,并且在一些情况下,产生了许多批次。比较这些aav9和aavrh79载体批次的产率(图19)。如实例1中针对aav8和aav9所描述的,评估aavrh79载体的脱酰胺化。结果示出,载体含有高度脱酰胺化的四种氨基酸(n57、n263、n385、n514),所述四种氨基酸基于aavrh79(seqidno:1)的编号对应于天冬酰胺-甘氨酸对中的天冬酰胺。在残基n94、n254、n410中始终观察到较低的脱酰胺化百分比。实例4:aav8.2.08的制备如在wo2017/180854(通过引用并入本文)中所讨论的,产生了若干个aav8突变体c41、c42、c46、g110、g113、g115和g117,其在hvr.viii区中具有突变。如gurda等人所讨论的,主要的adk8表位位于hvr.viii区(使用aav8vp1编号的氨基酸586到591)。在体外测试这些突变体的adk8抗性,并且在体内测试其中一些的adk8抗性。参见例如上文引用的lochrie2006。随机挑选ar2.1-9。基于频率选择ar2.25-61。就6孔板产率和huh7转导而言,随机挑选的变体以及频率较高的变体都是活的。图24示出在aav8.ar2.08的各种组织中的表达(最左边的一组条)。实例5:aav8.ar2.08的脱酰胺化seqidno:17和18中分别提供了aav8.ar2.08的新颖序列,其如实例4中进行设计。a.如实例1中针对aav8所描述的,产生修饰aav8.ar2.08载体,并评估其修饰。结果示出,载体含有高度脱酰胺化的五种氨基酸(n57、n263、n385、n514和n540),所述五种氨基酸基于aav8.ar2.08(seqidno:18)的编号对应于天冬酰胺-甘氨酸对中的天冬酰胺。在残基n94、n254、n410中始终观察到较低的脱酰胺化百分比。与aav8相反,在位置n459(aav8中的平均值为7%)或n499(在aav8中的平均值为17%)处未观察到脱酰胺化。b.单细胞rna-seq揭示了用aav8处理后从非人灵长类动物中分离出的转导肝细胞的组织定位和转录特征。单细胞rna测序已经被证明是用前所未有的单细胞分辨率表征细胞转录组的强大技术。在当前的工作中,利用单细胞rna-seq研究在用表达gfp的aav8载体处理后从恒河猕猴分离出的初级肝细胞的转录情况。facs分选的gfp+和gfp-细胞的转录组分析揭示了肝小叶内转导细胞的组织定位以及涉及肝细胞转导和转基因表达调节的基因和调节途径。对于研究设计,恒河猴用1×1013ddgc/kgaav8.tbg.egfp.wpre(n=1)或1×1013ddgc/kgaav8.2.08.tbg.egfp.wpre(aav8变体,n=1)处理。在处理后7天对动物实施安乐死,并进行尸检以从两只动物中分离出肝脏。在用胶原酶处理并进行梯度离心后,通过gfp转基因表达将分离的肝细胞facs分选到bdprecisetm96孔板上。从每只动物中分离出192个单细胞(96个gfp+和96个gfp-),并且然后按照标准bdprecisetm方案用于制备单细胞rna-seq库。使用r中的seurat、scran和scater包分析数据,以确定gfp-与gfp+分选的细胞之间差异表达的转录物,并使用建立的转录表达特征对肝小叶内分离的细胞进行空间重构。发现与aav8相比,aav8.ar2.08具有增加的肝向性,并且表现出转导效率增加1.5倍。对分选的肝细胞进行单细胞转录组分析揭示,表达转基因的细胞均匀分布在整个肝小叶中,示出对门静脉区的轻微偏好,这也在组织病理学中观察到。感兴趣的是,发现分选的gfp-细胞的亚群体以与分选的gfp+细胞相当的水平表达转基因转录物,这表明尽管没有可检测水平的转译蛋白,但实际上这些细胞已经被转导并表达转基因mrna。比较gfp-和gfp+细胞的转录谱揭示了参与病毒mrna转译的差异表达的转录物,从而阐明了涉及转导细胞中的转基因蛋白表达的调节的可能途径。c.通过人肝异种移植模型用定向进化对非人灵长类动物中具有更好肝脏转导和更高肝脏特异性的腺相关病毒8变体的分离为了得到具有更好转导和更高特异性的aav变体,使用靶向aav8衣壳上的表面暴露的位点的饱和诱变执行aav定向进化(即肝基因疗法的基准),随后在人肝异种移植小鼠模型中进行两轮体内富集,并分离出被称为aav8.2.08的aav8变体。在以1e13基因组拷贝(gc)/kg体重的剂量向非人灵长类动物静脉内注射后,与aav8相比,由aav8.ar2.08在各个器官(包含肺、心脏、胃、胰腺、肾脏和肠系膜淋巴结)中递送的载体基因组拷贝减少,而在肝脏中递送的载体基因组拷贝增加,这意味着更好的肝脏转导和更高的组织特异性。下一代测序表明,在体内选择期间,aav8.ar2.08的显著富集证明了用新的和改良的向性分离衣壳的方法的潜力。图21到27示出了其它比较。图28示出了aav8.ar2.08和aav8的生物分布。d.加条形码6只黑色小鼠静脉内注射有2×1012gc/小鼠的源自12种制剂的raavg3的混合物(图18b)。每种制剂含有载体基因组内单独的条形码,从而允许鉴定特异性制剂(图18a)。两周后,对动物实施安乐死,并采集组织。如所预测的,raavg3在肝脏中的表达高于在心脏或肌肉中的表达。图18c组织分布实验表明,注射的载体混合物中条形码的实际频率与理论频率相匹配(图18d:总计;图19a、19b:肌肉;图19c、19d:心脏;以及图19e、19f:肝脏),其中bc02和bc06中有轻微的异常(图20a-20c)。本说明书中引用的所有文件通过引用并入本文中。均于2018年8月24日提交的美国临时专利申请第62/722,388号和第62/722,382号、均于2018年7月26日提交的美国临时专利申请第62/703,670号和第62/703,673号、均于2018年5月29日提交的美国临时专利申请第62/677,471号和第62/677,474号、于2018年5月29日提交的美国临时专利申请第62/667,585号和于2018年2月27日提交的美国临时专利申请第62/635,964号通过引用并入本文中。于2018年5月7日提交的美国临时专利申请第62/667,881号、于2018年5月7日提交的美国临时专利申请第62/667,888号、于2018年5月6日提交的美国临时专利申请号第62/667,587号、于2018年4月27日提交的美国临时专利申请第62/663,797号、于2018年4月27日提交的美国临时专利申请第62/663,788号、于2018年2月27日提交的美国临时专利申请第62/635,968号通过引用并入本文。本文引用的并出现在所附序列表中的seqidno通过引用并入。尽管已经参考特定实施例描述了本发明,但是应当理解的是,可以在不脱离本发明的精神的情况下进行修改。这种修改旨在落入所附权利要求的范围内。当前第1页1 2 3
起点商标作为专业知识产权交易平台,可以帮助大家解决很多问题,如果大家想要了解更多知产交易信息请点击 【在线咨询】或添加微信 【19522093243】 与客服一对一沟通,为大家解决相关问题。
与客服一对一沟通,为大家解决相关问题。
此文章来源于网络,如有侵权,请联系删除
相关标签:

 热门咨询
热门咨询




tips