
 商标分类
商标分类  商标转让
商标转让 
与抗锈病相关的蛋白及其编码基因与应用的制作方法
 2021-02-02 16:02:33|
2021-02-02 16:02:33| 415|
415| 起点商标网
起点商标网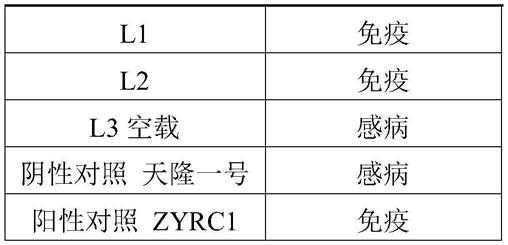
[0001]
本发明涉及植物基因工程领域,具体涉及一种与抗锈病相关的蛋白及其编码基因 与应用。
背景技术:
[0002]
大豆(glycine max)是全球四大油料作物之一,也是最重要的蛋白质源作物之一。 锈病是世界大豆生产的主要病害,通过使用叶面杀真菌剂是控制该病的主要方法。目 前尚没有对锈病具有完全抗性的商业化品种。锈病的病原菌为豆薯层锈菌phakopsora pachyrhizi,其寄主为广泛的豆科植物(分别为17个属中至少31种;slaminko等人, (2008)plantdis.,92:797-771;和19个属中至少42种;frederick等人,(2002)mycology, 92:217-227)。此外,另有152种已被认是大豆锈菌的潜在宿主(bonde等人, (2008)plantdis.,92:30-38;goellner等人,(2010)molecular plant pathology,11:169-177; ono等人,(1992)mycol.res.,96(10):825-850;和slaminko等人,(2008)plant dis., 92:797-771)。杀真菌剂应用是当前缓解锈病的唯一可用方法。
[0003]
对大豆锈菌具有抗性的大豆资源很少;由于大豆资源对大豆锈菌的抗性存在着生 理小种的特异性;在利用抗性资源进行育种时,如果忽视这些抗性特异性,可能会因 寄主抗性与生理小种的不相容而导致抗病资源所具有的抗性丧失,不利于抗性资源持 久性的利用。
[0004]
培育抗锈品种是防治锈病危害的最经济有效的途径。利用现代生物技术使不同物 种来源的抗性基因聚合到同一个品种中已成为可能。通过发掘大豆种质及其他豆科植 物中新的抗病基因,将其作为单个基因、多个基因,或以多基因盒的形式转移到大豆 时,这些抗性基因可以经由同源或异源表达提供对大豆锈菌的抗性。
技术实现要素:
[0005]
本发明的目的是提供一种与抗锈病相关的蛋白及其编码基因与应用。
[0006]
第一方面,本发明要求保护一种蛋白质。
[0007]
本发明所要求保护的蛋白质来源于大豆,命名为rpprc1,具体可为如下任一所 示蛋白质:
[0008]
(a1)氨基酸序列如seq id no.1所示的蛋白质;
[0009]
(a2)将seq id no.1所示的氨基酸序列经过一个或几个氨基酸残基的取代和/ 或缺失和/或添加且具有相同功能的蛋白质;
[0010]
(a3)与(a1)-(a2)中任一所限定的氨基酸序列具有99%以上、95%以上、 90%以上、85%以上或者80%以上同源性且具有相同功能的蛋白质;
[0011]
(a4)在(a1)-(a3)中任一所限定的蛋白质的n端和/或c端连接标签后得 到的融合蛋白。
[0012]
上述蛋白质中,所述标签是指利用dna体外重组技术,与目的蛋白一起融合表 达
j.4:3047-3053)。它们可单独使用或与其它的植 物启动子结合使用。。合适的转录终止子包括但不限于:农杆菌胭脂碱合成酶终止子 (nos终止子)、花椰菜花叶病毒camv 35s终止子、tml终止子、豌豆rbcs e9终止 子和胭脂氨酸和章鱼氨酸合酶终止子(参见,例如:odell等人(i
985
)nature 313:810; rosenberg等人(1987)gene,56:125;guerineau等人(1991)mol.gen.genet,262:141; proudfoot(1991)cell,64:671;sanfacon等人genes dev.,5:141;mogen等人(1990) plant cell,2:1261;munroe等人(1990)gene,91:151;ballad等人(1989)nucleic acids res.17:7891;joshi等人(1987)nucleic acid res.,15:9627)。
[0024]
构建含有所述核酸分子的重组表达载体。所利用的植物表达载体可为gateway系 统载体或双元农杆菌载体等,如pgwb411、pgwb412、pgwb405、pbin438、 pcambia1302、pcambia2301、pcambia1301、pcambia1300、pbi121、 pcambia1391-xa或pcambia1391-xb。使用rpprc1构建重组表达载体时,在其转 录起始核苷酸前可加上任何一种增强型、组成型、组织特异型或诱导型启动子,如花 椰菜花叶病毒(camv)35s启动子、泛生素基因ubiqutin启动子(pubi)等,它们 可单独使用或与其它的植物启动子结合使用;此外,使用本发明的基因构建植物表达 载体时,还可使用增强子,包括翻译增强子或转录增强子,这些增强子区域可以是atg 起始密码子或邻接区域起始密码子等,但必需与编码序列的阅读框相同,以保证整个 序列的正确翻译。所述翻译控制信号和起始密码子的来源是广泛的,可以是天然的, 也可以是合成的。翻译起始区域可以来自转录起始区域或结构基因。
[0025]
为了便于对转基因植物细胞或植物进行鉴定及筛选,可对所用植物表达载体进行 加工,如加入可在植物中表达的编码可产生颜色变化的酶或发光化合物的基因(gus 基因、萤光素酶基因等)、具有抗性的抗生素标记物(庆大霉素标记物、卡那霉素标记 物等)或是抗化学试剂标记基因(如抗除莠剂基因)等。
[0026]
所述转基因细胞系既可为繁殖材料,也可为非繁殖材料。
[0027]
在本发明的具体实施方式中,所述表达盒中启动所述蛋白质的编码基因转录的启 动子具体为所述编码基因的原始内源启动子,所述编码基因的原始内源启动子的核苷 酸序列如seq id no.7所示。
[0028]
在本发明的具体实施方式中,所述重组载体具体为将前文第二方面所述核酸分子 (seq id no.2)克隆到pb2gw7载体的attr1和attr2位点间,并且将saci和spei 酶切位点之间的35s启动子替换为seq id no.7所示rpprc1基因内源性启动子后得 到的重组质粒。
[0029]
第四方面,本发明要求保护前文第一方面中所述蛋白质或第二方面所述核酸分子 或第三方面所述重组载体、重组菌或转基因细胞系在调控植物对锈病抗性中的应用。
[0030]
在所述应用中,所述蛋白质或其编码基因在所述植物中的表达量和/或活性提高, 其对锈病的抗性增强。
[0031]
第五方面,本发明要求保护一种提高植物对锈病抗性的方法。
[0032]
本发明所要求保护的提高植物对锈病抗性的方法,可包括如下步骤:提高植物中 前文所述蛋白质的表达量和/或活性。
[0033]
其中,所述提高植物对锈病抗性,既可以指使原本对锈病无抗性的植物赋予锈病 抗性;也可以指使原本对锈病有抗性的植物对锈病的抗性得到进一步增强。所述提高 植物中所述蛋白质的表达量和/或活性,既可以指使原本不表达所述蛋白质的植物表达 所述蛋白质,也可以指使原本表达所述蛋白质的植物中所述蛋白质的表达量和/或活性 得到进一
面中所述核酸分子中的应用。
[0053]
(d6)所述引物对或所述探针或所述试剂盒在鉴定待测植物是否具有由前文第二 方面中所述核酸分子所带来的锈病抗性中的应用;
[0054]
(d7)dna分子,为seq id no.7所示dna分子,或者将seq id no.7进行一 个或多个核苷酸的添加、缺失和/或替换后得到的具有启动子功能的dna分子;
[0055]
(d8)所述dna分子在启动目的基因表达中的应用。进一步地,所述目的基因 可为前文第二方面中所述核酸分子。
[0056]
在上述各方面中,所述锈病可为豆科植物锈病。
[0057]
进一步地,所述豆科植物锈病可为大豆锈病。
[0058]
更进一步地,所述大豆锈病的病原菌可为大豆锈菌(phakopsora pachyrhizi)或山蚂 蟥层锈菌(phakopsora meibomiae)。
[0059]
在本发明的具体实施方式中,所述大豆锈病的病原菌具体为大豆锈菌(phakopsora pachyrhizi)生理小种ss4。
[0060]
在上述各方面中,所述植物可为豆科植物。
[0061]
进一步地,所述豆科植物可为大豆属植物、鹰嘴豆属植物、木豆属植物、扁豆属 植物、苜蓿属植物、菜豆属植物、豌豆属植物、葛属植物、三叶草属植物或豇豆属植 物。
[0062]
其中,所述大豆属植物可为沙生大豆(glycine arenaria)、银毛大豆(glycine argyrea)、弯裂片大豆(glycine cyrtoloba)、灰毛大豆(glycine canescens)、澎湖大豆 (glycine clandestine)、弯荚大豆(glycine curvata)、镰荚大豆(glycine falcata)、宽叶大豆 (glycine latifolia)、小叶大豆(glycine microphylla)、glycine pescadrensis、glycine stenophita、glycine syndetica、野生大豆(glycine soja seib.et zucc.)、栽培大豆(glycine max(l.)merrill.)、烟豆(glycine tabacina)或短绒野大豆(glycine tomentella)。
[0063]
所述鹰嘴豆属植物可为鹰嘴豆(cicerarietinum)、cicer echinospermum、cicer reticulatum或cicer pinnatifidum。
[0064]
所述扁豆属植物可为扁豆(lablab purpureus)。
[0065]
所述苜蓿属植物可为蒺藜苜蓿(medicago truncatula)或紫花苜蓿(medicago sativa)。
[0066]
所述菜豆属植物可为菜豆(phaseolus vulgaris)、棉豆(phaseolus lunatus)、宽叶菜豆 (phaseolus acutifolius)或红花菜豆(phaseolus coccineus)。
[0067]
所述豌豆属植物可为pisum abyssinicum、菜豌豆(pisum sativum)、pisum elatius、 pisum fulvum、pisum transcaucasium或pisumhumile。
[0068]
所述葛属植物可为葛根(pueraria lobata)。
[0069]
所述三叶草属植物可为黄车轴草(trifoliumaureum)或trifolium occidentale。
[0070]
所述豇豆属植物可为豇豆(vigna unguiculata)、vigna dalzelliana、长圆叶豇豆(vigna oblongifolia)、长虹豆(vigna parkeri)、vignafilicaulis、vigna kirkii、长叶豇豆(vigna luteola)、绿豆(vigna radiata)、三裂叶豇豆(vigna trilobata)、长叶豇豆(vigna luteola)或 黑吉豆(vigna mungo)。
[0071]
更进一步地,所述豆科植物可为如下任一:大豆、苜蓿、三叶草、豌豆、菜豆兵 豆、
羽扇豆、牧豆树、角豆、大豆、花生或罗望子。
[0072]
在本发明的具体实施方式中,所述植物具体为大豆品种天隆一号。
[0073]
本发明所公开的蛋白质及其编码基因与方法可用于保护植物免受锈病病原体的 侵害。宿主和病原体之间的相互作用可以“免疫性”到“部分抗性”到“易感性”的 连续体来描述。术语“免疫性”或“免疫”在本发明中用于表示不存在任何宏观可见 的病害症状。术语“部分抗性”在本发明中用于表示不具有或具有有限孢子形成的宏 观可见病变的存在和/或任何病害症状的范围或程度的减少和/或进展延缓,并且可以例 如表现为病变数减少或具有孢子形成减少的病变。如本发明所用,对锈病的术语“易 感性”或短语“缺乏抗性”是指在孢子形成水平等于或高于参考标准(诸如例如品种 williams或peking)中观察到的孢子形成水平的情况下,出现病变。
[0074]
术语“抗性”在本文中用于表示在植物中不存在由植物病原体所致的一种或多种 病害症状或所述病害症状减少。抗性可以是指当与对病害易感的植物或不包含减少一 种或多种病害症状的有效抗性基因的植物相比时,病害症状,例如发病植株的数量、 脱叶、和相关的产量损失降低、最小化或减少。另外,抗性可包括病原体增殖的预防 或延缓。广义上来讲,术语“抗性”包括如上文所定义的免疫性和部分抗性。
[0075]“植物病原体”在本文中可用于表示例如担子菌纲层锈菌属(phakopsora)(包括大 豆锈菌和山蚂蟥层锈菌)的真菌病原体。植物病害或豆科作物物种病害例如可以为锈 病。
[0076]
本发明公开的方法可以提高、增强或改善大豆对专性活体寄生真菌大豆锈菌(锈病 的主要病原菌)或对山蚂蟥层锈菌的抗性。例如,对锈菌病原增加或增强的抗性可与病 原在感病植株上的反应进行比较。抗性增加或增强的表现可以是不同层面的,但与在 植物或植物组织(例如,叶)上观察到的病害症状(如病斑颜色)、发病率相关。可以给出 免疫性、抗性和易感性的数值。例如,抗性的数值表示植物对植物病害(例如,锈病) 表现出的抗性程度。数值还可用于在例如所关注的植物(例如,转基因豆科植物)与易 感植物(例如天隆一号或williams)或参考标准之间进行抗性程度的比较。
[0077]
本发明所公开的蛋白质及其编码基因与方法涉及从豆科物种中分离一种抗性基 因,并且随后将该抗性基因转移到受体植物,例如大豆中,以提供或提高对豆薯层锈 菌的抗性。术语“抗病性基因”或“抗性基因”在本发明中用于表示编码能够增强或 改善植物中的防御或免疫系统应答蛋白质的基因。本发明公开的一个方面包括将抗性 基因移到性相容或不相容物种以产生抗病性。本发明所述的抗性基因可单独使用或与 其它抗性基因叠加使用或与非抗性基因聚合使用以提供或提高受体物种对锈病的抗 性。
[0078]
因此,本发明公开的转基因方法可单独使用或与其它策略组合使用以在植物中产 生或赋予锈病抗性。其它可用的策略包括但不限于阻断效应子的功能活性,抑制病原 体或病原体因子(例如真菌)摄入宿主细胞(例如植物细胞)中和/或常规的抗性育种。
[0079]
本发明公开的方法可提供或增强植物的锈病抗性,使得锈病的病原菌不能再繁殖 或繁殖系数显著下降。术语“增强”是指改善、增加、扩增、繁殖、升高和/或提升, 从而减少一种或多种病害症状。因此,当与对层锈菌属易感或耐受的植物相比时,本 发明所述的方法可减轻豆科植物锈病的一种或多种症状(即,病害症状)。本发明所指 的植物亦包括将采用基因工程方法将抗病基因或蛋白质导入豆科植物(例如,大豆), 从而使其在暴露于豆科植物锈病时,对病害增强抗性的转基因豆科植物。
[0080]
术语“植物”在本文中用于包括任何植物、组织或器官(例如,植物部分)。植物 部分包括但不限于可被培养成整个植株的细胞、茎、根、花、胚珠、雄蕊、种子、叶。 植物细胞是植物的细胞,其直接从种子或植株中获取,或者来源于由植物获取的细胞 培养。再生植物的子代、变体和突变体在本发明的范围内,前提条件是这些部分包含 被引入的抗性基因。
[0081]
本发明所述植物、植物部分或植物细胞来源于植物,所述植物包括但不限于大豆、 苜蓿、三叶草、豌豆、菜豆、兵豆、羽扇豆、牧豆树、角豆树、花生和罗望子。
[0082]
本发明所述植物为豆科。豆科的示例包括但不限于菜豆属(例如,法国菜豆、四 季豆、菜豆(phaseolus vulgaris)、棉豆(phaseolus lunatus)、宽叶菜豆 (phaseolusacutifolius)、红花菜豆(phaseolus coccineus));大豆属(例如,野大豆(glycine soja)、大豆(glycine max(l.)));豌豆(pisum)(例如,脱荚豌豆(有时被称为光皮豌豆或 圆粒豌豆;菜豌豆(pisum sativum));皱粒豌豆(pisum sativum)、糖荚豌豆(pisum sativum),其也被称为糖荚豌豆(snow pea)、食荚豌豆(edible-podded pea)或嫩豌豆 (pisumgranda));花生(arachis hypogaea)、三叶草(三叶草属(trifolium spp.))、苜蓿 (medicago)、葛藤(pueraria lobata)、常见的苜蓿、苜蓿(紫花苜蓿(medicago sativa))、 鹰嘴豆(鹰嘴豆属(cicer))、兵豆(lens culinaris)、羽扇豆(lupinus);野豌豆(vicia)、蚕 豆(vicia faba)、连理草(山黧豆属(lathyrus))(例如家山黧豆(lathyrussativus)、玫红山 黧豆(lathyrus tuberosus));豇豆属(例如,乌头叶菜豆(vignaaconitifolia)、赤豆(vigna angularis)、黑吉豆(vigna mungo)、绿豆(vigna radiata)、班巴拉花生(vigna subterrane)、 赤小豆(vigna umbellata)、野豇豆(vigna vexillata)、豇豆(vigna unguiculata)(也称为长 豇豆、豇豆));木豆(cajanus cajan)、硬皮豆属(macrotyloma)(例如,落花生(macrotyloma geocarpum)、马蚕豆(macrotylomauniflorum);四棱豆(psophocarpus tetragonolobus)、 非洲豆薯(sphenostylisstenocarpa)、埃及黑豆、鹊豆(lablab purpureus)、豆薯 (pachyrhizus erosus)、瓜尔豆(cyamopsis tetragonolobus);和/或刀豆属(例如,矮刀豆 (canavalia ensiformis))、刀豆(canavalia gladiata)。
[0083]
在本发明中,“核酸”是指单链或双链形式的脱氧核糖核苷酸或核糖核苷酸聚合 物,并且除非另外限制,否则涵盖在以下方面具有天然核苷酸的基本性质的已知类似 物(例如肽核酸):其以与天然存在的核苷酸相似的方式杂交至单链核酸。
[0084]
术语“蛋白质”是指氨基酸残基的聚合物。该术语适用于其中一个或多个氨基酸 残基为相应的天然氨基酸的人工化学类似物的氨基酸聚合物,并且适用于天然氨基酸 聚合物。本发明的蛋白质可以由本发明所公开的核酸分子产生,或者通过使用标准的 分子生物学技术产生。
[0085]
术语“编码”在本发明中用于表示核酸包含所需信息,其通过使用密码子以指导 核苷酸序列(例如,豆科序列)翻译成特定蛋白质来指定。编码蛋白质的核酸可以包含 在所述核酸的翻译区内的非翻译序列(例如内含子)或可以缺少这种居间的非翻译序列 (例如,如在cdna中)。
[0086]
本发明涵盖分离的或实质上纯化的核酸分子或蛋白质。“分离的”或“纯化的
”ꢀ
核酸分子或蛋白质或其生物活性部分,实质上或基本上不含通常在该核酸分子或蛋白 质的天然环境中存在的、与该核酸分子或蛋白质相伴随或相互作用的成分。因此,分 离的或纯
化的核酸分子或蛋白质,当通过重组技术(例如pcr扩增)生产时实质上不含 其它细胞物质或培养基,或者当通过化学法合成时实质上不含化学前体或其它化学品。 最佳地,“分离的”核酸分子不含在衍生该核酸分子的生物体的基因组dna中天然位 于该核酸分子的旁侧的序列(即,位于该核酸分子的5
′
和3
′
末端的序列)(例如,蛋白 质编码序列)。例如,在本发明的一些实施方案中,分离的核酸分子可含有少于约5kb、 约4kb、约3kb、约2kb、约1kb、约0.5kb或约0.1kb的在衍生该核酸分子的细胞的基 因组dna中天然位于该核酸分子的旁侧的核苷酸序列。基本上不含细胞物质的蛋白 质包括具有小于约30%、约20%、约10%、约5%或约1%(以干重计)的污染性蛋白 质的蛋白质制剂。当实施方案的蛋白质或其生物活性部分用重组法产生时,最佳地, 培养基具有少于约30%、约20%、约10%、约5%或约1%(以干重计)的化学前体或 非受关注蛋白的化学品。
[0087]
与编码的核苷酸序列和蛋白质相关的片段和变体在本发明公开的范围内。“片 段”是指核苷酸序列的一部分或由其编码的氨基酸序列以及蛋白质的一部分。核苷酸 序列的片段可编码蛋白质片段,所述蛋白质片段保留天然蛋白质的生物学活性并具有 赋予对植物的抗性(即,抗真菌性)的能力。作为另一选择,可用作杂交探针的核苷酸 序列片段不一定编码保持生物活性的片段蛋白质。因此,核苷酸序列的片段可在至少 约15个核苷酸、约50个核苷酸、约100个核苷酸并且至多编码本发明公开的蛋白质 的全长核苷酸序列的范围内。
[0088]
编码本发明公开的蛋白质的生物活性部分的核苷酸序列的片段可编码至少约15、 约25、约30、约40、或45、约50个连续氨基酸或者至多该实施方案的全长蛋白中存 在的氨基酸的总数(例如对于seq id no.1而言,857个氨基酸)。可用作杂交探针或 pcr引物的核苷酸序列的片段通常不必编码蛋白质的生物活性部分。
[0089]
当涉及指定的核酸分子时,术语“全长序列”是指具有天然序列的整个核酸序列。
ꢀ“
天然序列”在本发明中用于表示内源性序列,即存在于生物体基因组中的非工程化 序列。
[0090]
因此,本发明公开的核苷酸序列的片段可编码蛋白质的生物活性部分,或者它可 以是用作杂交探针或pcr引物的片段。本发明的核酸分子包含至少约15、约20、约 50、约75、约100、或约150个核苷酸或者至多本发明所公开的全长核苷酸序列中存 在的核苷酸的数目(例如对于seq id no.2而言,2574个核苷酸)。
[0091]
术语“变体”表示基本上类似的序列。对于核酸分子,变体包含在天然核酸分子 中的一个或多个位点处的一个或多个核苷酸的缺失和/或添加,和/或在天然核酸分子中 的一个或多个位点处的一个或多个核苷酸的置换。如本发明所用,“天然”核酸分子或 蛋白质分别包含天然存在的核苷酸序列或氨基酸序列。本领域技术人员可认识到本发 明中的核酸变体将被构造成使得开放阅读框得以保持。对于核酸分子,保守变体包括 那些这样的序列,其由于遗传密码的简并性而编码本发明蛋白质中的氨基酸序列。天 然等位基因变体可用公知的分子生物学技术进行鉴定,例如用聚合酶链反应(pcr)和杂 交技术来鉴定。变体核酸分子还包括通过合成来源的核酸分子,例如通过使用定点诱 变产生但仍编码本发明蛋白质的那些核酸分子。通常,如通过本领域熟知的序列比对 程序所测定的,本发明公开的特定核酸分子的变体可与该特定核酸分子具有至少约40 %、约45%、约50%、约55%、约60%、约65%、约70%、约75%、约80%、约 85%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、 约98%、约99%或更高的序列同一性。
[0092]
本发明的特定核酸分子(即参考核酸分子)的变体还可通过比较变体核酸分子所编 码的蛋白质与该参考核酸分子所编码的蛋白质之间的序列同一性百分比来进行评价。 任何两个蛋白质之间的序列同一性百分比可用本领域已知的序列比对程序来计算。在 任何给定的本发明核酸分子对是通过比较它们编码的两个蛋白质所共享序列同一性百 分比进行评价的情况中,其中两个被编码的蛋白质之间的序列同一性百分比为至少约 40%、约45%、约50%、约55%、约60%、约65%、约70%、约75%、约80%、 约85%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、 约98%、约99%或更高的序列同一性。
[0093]“变体蛋白质”是指通过在天然蛋白质中的一个或多个位点处缺失或添加一个或 多个氨基酸和/或在天然蛋白质的一个或多个位点处置换一个或多个氨基酸而从该天 然蛋白质衍生的蛋白质。本发明所涵盖的变体蛋白是具有生物活性的,也就是说它们 继续拥有天然蛋白质的所需生物活性,即如本发明所述增强植物抗性(即,植物真菌病 原体抗性)的能力。此类变体可例如由遗传多态性或由人类操纵而得到。如通过本领域 已知的序列比对程序所测定的,本发明的天然蛋白质的生物活性变体可与该天然蛋白 质的氨基酸序列具有至少约40%、约45%、约50%、约55%、约60%、约65%、约 70%、约75%、约80%、约85%、约90%、约91%、约92%、约93%、约94%、 约95%、约96%、约97%、约98%、约99%或更高的序列同一性。本发明公开的蛋 白质的生物活性变体与该蛋白可相差少至约1-15个氨基酸残基,少至约1-10个(如约 6-10个)、少至约5个、少至4个、3个、2个或甚至1个氨基酸残基。
[0094]
本发明所公开的蛋白质可例如通过包括氨基酸置换、缺失、截短和插入来进行变 更。此类操纵的方法在本领域中是已知的。例如,抗性蛋白的氨基酸序列变体和片段 可通过在dna中作出突变来制备。诱变和多核苷酸变更的方法是本领域已知的。
[0095]
变体核酸分子和蛋白质还涵盖由诱变和重组程序,包括但不限于诸如dna改组 的程序获得的序列和蛋白质。可从一群包含具有实质序列同一性并可在体外或体内进 行同源重组的序列区的相关序列多核苷酸产生重组多核苷酸的文库。例如,使用该方 法,可使编码受关注结构域的序列基序在本发明公开的蛋白质基因和其它已知的蛋白 质基因之间发生改组,以获得编码具有改善的受关注性质(诸如增加的赋予或增强植物 对真菌病原体的抗性的能力)的蛋白质的新基因。这种dna改组的策略是本领域已知 的。
[0096]
本本发明公开涵盖基于其与本发明示出的整个序列或与其变体和片段的序列同 一性而分离的序列。这类序列包括为所公开的序列的直系同源物的序列。术语“直系 同源物”是指衍生自共同祖先基因并且由于物种形成而存在于不同物种中的基因。存 在于不同物种中的基因,当它们的核苷酸序列和/或它们的编码蛋白质序列共享至少约 60%、约70%、约75%、约80%、约85%、约90%、约91%、约92%、约93%、 约94%、约95%、约96%、约97%、约98%、约99%或更大的序列同一性时,被认 为是直系同源物。直系同源物的功能在各种物种中常常是高度保守的。因此,本发明 公开涵盖编码赋予或增强真菌植物病原体抗性的蛋白质并且与本发明所公开的序列杂 交,或与其变体或片段杂交的分离的核酸分子。
[0097]
在pcr方法中,可设计寡核苷酸引物用于pcr反应,以从提取自任何受关注生 物体的cdna或基因组dna扩增相应的dna序列。设计pcr引物和pcr克隆的方 法是本领域已知的,在以下文献中公开:sambrook等人,(1989)molecular cloning:a laboratory manual(第2
形式。“稳定转化”或“稳定地转化”是指被引入植物中的核苷酸构建体整合进植物的 基因组中,并能够由其后代继承。如本文所用,“瞬时转化”意指将核酸分子引入植物 中但它没有整合到植物的基因组中,或者将蛋白质引入植物中。
[0112]
转化方法以及将核酸分子序列引入植物中的方法,可取决于要进行转化的植物或 植物细胞的类型。将蛋白质或核酸分子引入植物细胞中的合适方法包括但不限于微注 射、电穿孔、直接基因转移、lec1转化和弹道粒子加速。随着更新的方法变得可用, 其也可施用于本发明,因为转化或转染的方法并不是关键。
[0113]
可根据常规方式将已转化的细胞培育成植株。然后可使这些植株生长,并用同一 转化株系或不同株系授粉,再鉴定出具有所需表型特征的组成型表达的子代。可培育 两代或更多代植株,以确保所需表型特征的表达得到稳定保持和遗传。然后收获种子, 以确保已实现所需表型特征的表达。在本发明的一些方面,将具有核苷酸构建体或表 达盒的转化的种子或转基因种子稳定地并入其基因组中。
[0114]
本发明涵盖包含本发明所公开的核酸分子序列的种子,其可发育成或可用于使一 种或多种植物发育,与例如野生型品种的植物种子比,所述植物具有对病原体(例如真 菌)或由病原体形成的感染的增强的抗性。本发明的特征在于来自转基因豆科植物的种 子,其中所述种子包含本发明所公开的核酸分子。
[0115]
受关注的植物包括豆科作物物种,其包括但不限于苜蓿(medicagosativa);三叶草 或车轴草(三叶草属物种);豌豆,包括(豌豆(pisum satinum))、木豆(gajanus cajan)、豇 豆(vigna unguiculata)和山黧豆属(lathyrus spp.);菜豆(fabaceae或leguminosae);兵豆 (lens culinaris);羽扇豆(lupinus spp.);牧豆树(prosopis spp.);长豆角(ceratonia siliqua)、 大豆(glycine max)、花生(arachishypogaea)或罗望子(tamarindus indica)。术语“豆科物 种”和“豆科作物物种”在本文中用于是指植物,并且可以例如为受关注的植物豆科 物种或豆科作物物种可以是植物、植物部分或植物细胞。
[0116]
术语“转基因”在本文中用于是指植物,包括来源于植物的任何部分,诸如细胞、 组织或器官,其中通过基因工程方法,诸如农杆菌属转化,将外源性核酸(例如,包括 一种或多种核酸的重组构建体、载体或表达盒)整合进基因组中。通过进行基因技术方 法,将外源性核酸稳定整合进染色体中,使得下一代也可以是转基因的。如本文所用,
ꢀ“
转基因”还涵盖生物处理,其包括植物杂交和/或天然重组。
[0117]
构建的载体或表达盒不存在于初始植物的基因组中或转基因植物的基因组中,也 不在其初始植物的基因组的天然基因座处。
[0118]
本发明所公开的组合物可通过基因掺入的方法来生成或维持。当所述方法重复两 次或更多次时,基因掺入有时被称为“回交”。在基因掺入或回交时,“供体”亲本是 指具有待渗入的所需基因或基因座的亲本植物。“受体”亲本(使用一次或多次)或“轮 回”亲本(使用两次或更多次)是指基因或基因座被基因掺入到其中的亲本植物。初始 杂交产生f1代;术语“bc1”则是指轮回亲本的第二次使用,并且“bc2”是指轮回 亲本的第三次使用,等等。
[0119]
因此,本发明公开的是增强植物对植物病害诸如锈病的抗性的方法。所述方法可 包括通过在育种程序(即,抗锈病的育种程序)中将豆科植物来源的抗性基因掺入种质 中来赋予对病原体(如锈病)的抗性。
[0120]
术语“种质”在本发明中用于表示来自个体(例如植物)、一组个体(例如,植物种 系、品种或家族)的遗传物质,或源自品系、品种、物种或培养物的克隆体。种质可以 为生物体或细胞的部分,或可以从生物体或细胞分离。种质提供具有特定分子构成的 遗传物质,所述特定分子构成提供有机体或细胞培养物的一些或全部遗传性质的物理 基础。在本发明的上下文中,种质包括新植物可由此生成的细胞、种子或组织,或者 可被培育成整株植物的植物部分诸如叶、茎、花粉、或细胞。
[0121]
本发明还可包括所述的序列,其可由受关注植物中表达的表达盒或dna构建体 中提供。该表达盒可包括与本发明所公开的序列可操作地连接的5
′
和3
′
异源调控序 列。术语“可操作地连接”在本发明中用于表示以允许核酸表达的方式(即,当将载体 引入宿主植物细胞中时,在宿主植物细胞中)将待表达的核酸连接至调控序列,包括启 动子、终止子、增强子和/或其它表达控制元件(例如,聚腺苷酸化信号)。此类调控序 列是本领域中所熟知的并且包括在多种宿主细胞中可直接组成型表达的,和在特定宿 主细胞中或在特定条件下可直接表达的那些核苷酸序列。载体的设计可取决于例如待 转化的宿主细胞的类型,或期望的核酸表达水平。所述表达盒可含有一个或多个待共 转化到植物中的附加基因。并且,可在多个表达盒上提供任何附加基因。
[0122]
本发明的表达盒可包括多个限制性酶切位点用于插入该核苷酸序列以便处于调 节区的转录调节之下。该表达盒还可含有选择性标记基因。
[0123]
表达盒还可在5
′-
3
′
转录方向上包括在植物中起作用的转录和翻译起始区、本 公开的dna序列、以及转录和翻译终止区。转录起始区,启动子,相对于植物宿主, 可以是天然的或类似的,或是外来或异源的。另外,该启动子可以是天然序列或另选 地是合成序列。术语“异源的”是指转录初始区不存在于所述转录初始区被引入其中 的天然植物中。如本发明所用,嵌合基因包含与转录起始区可操作连接的编码序列, 该转录起始区对于该编码序列是异源的。启动子的示例包括但不限于,花椰菜花叶病 毒35s和大豆泛素6。
[0124]
虽然可优选使用异源启动子来表达序列,但可以使用同源启动子或天然启动子序 列。此类构建体将改变宿主细胞(即,植物或植物细胞)中的表达水平。因此,改变宿 主细胞(即,植物或植物细胞)的表型。
[0125]
终止区可以天然具有转录起始区,天然具有可操作地连接的受关注的dna序列, 或源自另一个源。易得的终止区可获自根瘤农杆菌(agrobacterium tumefaciens)的ti质 粒,如章鱼碱合酶和胭脂碱合酶终止区。
[0126]
内源性或源基因抗性直系同源物可通过同源或非同源重组方法,诸如例如通过基 因组编辑来更改。当与其未修饰的序列相比时,此类更改是指核苷酸序列具有至少一 种修饰,并且包括例如:(i)置换至少一个核苷酸,(ii)缺失至少一个核苷酸,(iii)插入 至少一个核苷酸,或者(iv)(i)-(iii)的任何组合。
[0127]
在一些实施方案中,可使用基因组编辑技术将本发明所公开抗性基因引入植物的 基因组中,或者可以使用基因组编辑技术来编辑先前引入植物基因组中的抗性基因。
[0128]
基因组编辑可使用任何可用的基因编辑方法来实现。例如,基因编辑可通过将多 核苷酸修饰模板(有时被称为基因修复寡核苷酸)引入宿主细胞中来实现,其中所述多 核苷酸修饰模板包括对宿主细胞基因组内的基因的靶向修饰。多核苷酸修饰模板可以 为单链或双链。
[0129]
根据需要,可优化一种或多种基因以增加在转化的植物中的表达。如使用植物优 选的密码子来合成基因以改善表达。用于合成植物优选的基因的方法是本领域已知的。
[0130]
已知有另外的序列修饰能增强细胞宿主中的基因表达。这些序列修饰包括消除以 下序列:编码假多腺苷酸化信号、外显子-内含子剪接位点信号、转座子样重复序列, 以及其它此类得到充分表征的可能对基因表达有害的序列。可将序列的g-c含量调整 到给定的细胞宿主的平均水平,该平均水平可通过在宿主细胞中表达的已知基因计算 得到。必要时可修饰该序列以避免可能产生的发夹二级mrna结构。
[0131]
表达盒可以在表达盒构建体中额外含有5
′
前导序列。此类前导序列可以起到增 强翻译的作用。翻译前导序列是本领域已知的,它包括:小核糖核酸病毒前导序列, 如emcv前导序列(脑心肌炎5
′
非编码区);马铃薯y病毒组前导序列,如tev前导 序列(烟草蚀纹病毒),和人免疫球蛋白重链结合蛋白(bip);来自苜蓿花叶病毒的外壳 蛋白mrna(amvrna 4)的非翻译前导序列;烟草花叶病毒前导序列(tmv);以及玉 蜀黍枯黄斑点病毒前导序列(mcmv)(lommel等人,(1991)virology 81:382385)。也 可利用其它已知的用于增强翻译的方法,诸如内含子。
[0132]
可根据需要定,在适当的阅读框中,操纵表达盒中的各种dna片段,以确保dna 序列处于正确方向。为此,可以使用衔接子或接头来连接dna片段。另外,也可使 用其它操纵来提供便利的限制位点、移除多余的dna、或移除限制位点。出于这个目 的,可能涉及体外诱变、引物修复、限制、退火、再置换(例如转换和颠换)。
[0133]
一般来讲,表达盒可包含用于选择转化的细胞的选择性标记基因。选择性标记基 因用于选择转化的细胞或组织。标记基因包括编码抗生素抗性的基因,诸如那些编码 新霉素磷酸转移酶ii(neo)和潮霉素磷酸转移酶(hpt)的基因,以及赋予对除草化合物 的抗性的基因,所述除草化合物为诸如草铵膦、草丁膦、溴草腈、咪唑啉酮和2,4
-ꢀ
二氯苯氧基乙酸(2,4-d)。以上选择性标记基因的列表并不意指是限制性的。任何选 择性标记基因均可用于本公开。
[0134]
为了在植物或植物细胞中表达本发明的靶基因和/或蛋白质,本发明所述的方法包 括用编码靶标蛋白的核酸分子转化植物或植物细胞。本发明所述核酸分子可以被操作 连接至启动子,所述启动子驱动植物细胞中的表达。本领域中已知的任何启动子均可 用于本发明的方法中,其包括但不限于组成型启动子、病原体诱导型启动子、伤口诱 导型启动子、组织优选的启动子和化学调节启动子。启动子的选择可取决于转化的植 物中期望的表达时间和位置,以及本领域技术人员已知的其它因素。可种植或培育转 化的细胞或植物以形成一种植物,所述植物包含被引入例如编码r蛋白的细胞或植物 中的多核苷酸中的一种或多种。
[0135]
多种启动子可用于实施本发明。可根据期望的结果来选择启动子。即,可将核酸 与组成型启动子、组织优选的启动子、或其它启动子进行组合,在受关注的宿主细胞 中表达。此类组成型启动子包括(例如)rsyn7启动子的核心启动子和wo 99/43838和 美国专利6,072,050中公开的其他组成型启动子;camv 35s启动子;水稻肌动蛋白; 泛素;pemu;mas;als等。其在本领域中是已知的,并且可设想用于本公开中。
[0136]
一般来讲,采用诱导型启动子特别是来自病原体的诱导型启动子表达本基因是有 利于本基因的应用。这类启动子包括来自发病相关蛋白(pr蛋白)的启动子,所述蛋白 受病
原体感染诱导形成,例如pr蛋白、sar蛋白、β-1,3-葡聚糖酶、壳多糖酶等等。
[0137]
在病原体感染位点处或附近局部表达的启动子值得关注。另外,因为病原体可通 过伤口或昆虫病变进入植物中,伤口诱导型启动子也可用于本发明的载体构建。此类 伤口诱导型启动子包括马铃薯蛋白酶抑制剂(pinii)基因、wun1和wun2、win1和win2、 系统素(systemin)、wip1、mpi基因等。
[0138]
化学调控启动子可通过施用外源化学调控剂来调节基因在植物中的表达。根据利 用目的,启动子可以是化学诱导型启动子,如通过施用化学物质诱导基因表达;或者 是化学阻遏型启动子,如通过施用化学物质抑制了基因表达。化学诱导型启动子是本 领域已知的,包括但不限于玉蜀黍in2-2启动子(其通过苯磺酰胺除草剂安全剂激活)、 玉蜀黍gst启动子(其通过用作前突生除草剂的疏水性亲电化合物激活)和烟草pr-1a 启动子(其通过水杨酸激活)。受关注的其它化学调节的启动子包括甾类化合物响应性 启动子(例如,糖皮质素诱导型启动子、和四环素诱导型启动子和四环素阻遏型启动 子)。
[0139]
组织优选的启动子可用于靶基因或蛋白质(例如,编码豆科植物来源的nb-lrr 多肽的多核苷酸序列)在特定植物组织内的靶向增强的表达。此类组织优选的启动子包 括但不限于叶片特异性启动子、根系特异性启动子、种子特异性启动子和茎部特异性 启动子。组织特异性启动子包括yamamoto等人,(1997)plant j.12(2):255-265(yamamoto 等人,1997年,《植物杂志》,第12卷,第2期,第255-265页);kawamata等人, (1997)plant cell physiol.38(7):792-803;hansen等人,(1997)mol.gen genet.254(3): 337-343;russell等人,(1997)transgenic res.6(2):157-168;rinehart等人,(1996)plant physiol.112(3):1331-1341;van camp等人,(1996)plant physiol.112(2):525-535; canevascini等人,(1996)plant physiol.112(2):513-524;yamamoto等人,(1994)plant cell physiol.35(5):773-778;lam,(1994)results probl.cell differ.20:181-196;orozco等 人,(1993)plant molbiol.23(6):1129-1138;matsuoka等人,(1993)proc natl.acad.sci.usa 90(20):9586-9590;以及guevara-garcia等人,(1993)plant j.4(3):495-505。此类启动 子可被用于修饰本公开的核苷酸序列。
[0140]
叶特异性启动子是本领域已知的。参见例如,yamamoto等人,(1997)plant j.12(2): 255-265;kwon等人,(1994)plant physiol.105:357-67;yamamoto等人,(1994)plantcell physiol.35(5):773-778;gotor等人,(1993)plant j.3:509-18;orozco等人,(1993)plant mol.biol.23(6):1129-1138;以及matsuoka等人,(1993)proc.natl.acad.sci.usa90(20): 9586-9590。
[0141]“种子优选的”启动子包括“种子特异性”启动子(那些启动子在种子发育期间活 跃,诸如种子贮藏蛋白的启动子)和“种子萌发”启动子(那些启动子在种子萌发期间 活跃)。此类种子优选的启动子包括但不限于cim1(细胞分裂素诱导信息)、mi1ps(肌醇
ꢀ-
1-磷酸合酶)和cela(纤维素合酶)。球蛋白-1(glob-1)是优选的胚特异性启动子。对于 双子叶植物而言,种子特异性启动子包括但不限于菜豆β-菜豆蛋白基因启动子、油菜 籽蛋白(napin)基因启动子、β-伴球蛋白基因启动子、大豆凝集素基因启动子、十字花 科蛋白基因启动子等。
[0142]
本发明的核酸分子的表达可涉及使用完整的天然抗性基因,其中所述表达由同源 5
′
上游启动子序列驱动或其他异源启动子。本领域技术人员将能够鉴定抗性基因,以 评价
其表达水平,并且选择可用于表达受关注的抗性基因的优选的启动子序列。使用 同源或异源抗性基因启动子序列提供调节蛋白质表达的选择,以避免或最小化那些与 植物防御活化相关的任何潜在的不适当或不期望的结果。
[0143]
特异性大豆启动子包括但不限于大豆泛素(subi-1)、延长因子1a和用于组成型表 达的s-腺苷甲硫氨酸合酶和rpp4、rpg1-b以及包含在基因模型中的启动子,例如本 领域技术人员已知用于更紧密地调节由nb-lrr基因启动子提供的表达的glyma启动 子。
[0144]
本发明还包括用于本文所述的测定的试剂盒。蛋白质或核酸分子或含有所述核酸 分子的表达盒、重组载体或重组菌或转基因细胞系还可作为试剂盒的组成与说明书一 起包装,用于完成本发明所公开的测定。本发明的试剂盒可包括本发明所述的蛋白质 或核酸分子或含有所述核酸分子的表达盒、重组载体或重组菌或转基因细胞系和适宜 的说明书(书面和/或作为音频、视觉或视听材料提供)的任何组合。例如,试剂盒还可 包括特定的探针,所述特定的探针具有的序列对应于以下序列或与以下序列互补,所 述序列与转基因事件的特定区域具有介于80%和100%之间的序列同一性。试剂盒可 包括进行测定或检测方法所需的任何试剂和材料。
[0145]
除非上下文另外明确规定,如本发明所用的单数形式“一个”、“一种”和“该
”ꢀ
包括多个指代物。因此,例如,提及“一个细胞”包括多个此类细胞,并且提及“该 蛋白”包括提及一种或多种蛋白以及本领域技术人员已知的其等同物,等等。除非另 外明确指明,否则本文所用的所有技术和科学术语均具有本发明所属领域的普通技术 人员通常所理解的相同含义。
[0146]
如说明书和权利要求中所用的,术语“包含”可包括“由...组成”和“基本上由... 组成”的方面。“包含”还可表示“包括但不限于”。
[0147]
如本发明所用,词语“或”是指具体列表中的任一个成员,并且还包括该列表中 成员的任何组合。
[0148]
如本发明所用,术语“大豆”是指大豆和利用大豆育种或培养的任何植物品种。
[0149]
范围在本发明中可表示为从“约”一个具体值,和/或至“约”另一个具体值。当 表达此类范围时,其它方面包括从一个具体值和/或至其它具体值。类似地,当值被表 示为近似值时,应当理解,通过使用先行词“约”,该具体值形成另一方面。还应当理 解,所述范围中每一个的端点既与另一端点显著相关又独立于另一端点。还应当理解, 存在本发明所公开的多个值并且除了该值本身之外,每个值在本文中也以“约”该具 体值的形式公开。例如,如果公开了值“10”,则也公开了“约10”。还应当理解,还 公开了两个特定单元之间的每个单元。例如,如果公开了10和15,则也公开了11、 12、13和14。
[0150]
本发明的实验证明,将rpprc1基因转入感病大豆品种天隆一号中得到的转基因 大豆对锈病的抗性显著高于受体亲本天隆一号,说明rpprc1及其编码基因可以调控 豆科植物对锈病的抗性,过表达后提高了植物的锈病抗性。rpprc1及其编码基因可 用于提高豆科作物的抗病性,对于培育抗病新品种具有重要意义。
[0151]
保藏说明
[0152]
分类命名:大豆
[0153]
拉丁名:glycine max
[0154]
参椐的生物材料:zyrc1
[0155]
保藏机构:中国微生物菌种保藏管理委员会普通微生物中心
[0156]
保藏机构简称:cgmcc
[0157]
地址:北京市朝阳区北辰西路1号院3号
[0158]
保藏日期:2019年6月28日
[0159]
保藏中心登记入册编号:cgmcc no.17575
附图说明
[0160]
图1为抗锈基因rpprc1重组载体pb2gw7-rpprc1的质粒图谱。
[0161]
图2为t1代rpprc1转基因植株pcr检测图。m:marker。l1-1、l1-2、l1-3 为转化事件l1的部分t1代单株,l2-1、l2-2、l2-3为转化事件l2的部分t1代单株, 阴:阴性对照天隆一号,阳:阳性对照zyrc1。
[0162]
图3为rpprc1基因在t1代转基因植株中表达的rt-pcr鉴定。l1-2为转化事 件l1的一个t1代单株,l2-1为转化事件l2的一个t1代单株。
[0163]
图4为转基因植株southern分析图。a:rpprc1转基因植株southern分析图。 l1-1、l1-2、l1-3、l1-4、l1-5分别为转化事件l1的部分t1单株。l2-1、l2-2为转 化事件l2的部分t1代单株,天隆一号为阴性对照。b:空载转基因植株southern分 析图。l3-1、l3-2、l3-3、l3-4、l3-5、l3-6为空载转化事件l3的部分t1代单株。 ck为天隆一号。
[0164]
图5为t0代转基因植株接种12天后抗性鉴定表现。zyrc1为抗性对照,表现 免疫;rpprc1转化事件l2表现免疫;空载转化事件l3表现感病;非转基因天隆一 号表现感病。
[0165]
图6为t1代转基因植株接种12天后抗性鉴定表现。zyrc1为抗性对照,表现 免疫;rpprc1转化事件l2的t1代单株l2-1表现免疫;阴性对照天隆一号表现感病。
[0166]
注:各图中,同一转化事件标注相同,同一单株标注相同。
具体实施方式
[0167]
下述实施例中所使用的实验方法如无特殊说明,均为常规方法。
[0168]
下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
[0169]
实施例1、rpprc1基因的克隆
[0170]
大豆是古四倍体豆科植物,自花授粉,其基因组大小为约1.1gbp。不同抗性的大 豆种质对大豆锈菌的反应有着明显的差异。大豆(glycine max(l.)merrill)zyrc1 是中国农业科学院油料作物研究所从中国大豆种质筛选得到的一个抗锈资源,该品种 现由中国微生物菌种保藏管理委员会普通微生物中心保存,保藏编号为cgmcc no. 17575,其对大豆锈菌的反应为免疫。
[0171]
大豆zyrc1(rpprc1基因的来源)的基因组为模板,采用引物f和引物r(序 列如下)进行pcr扩增。
[0172]
f:5
’-
atggcagatagtgttgttgcttttctgc-3
’
(seq id no.3);
[0173]
r:5
’-
tcacagttcattagagattttgagcttacagc-3
’
(seq id no.4)。
[0174]
所得扩增产物为seq id no.2所示序列。seq id no.2即为rpprc1基因的核苷酸 序列,其编码seq id no.1所示蛋白质,该蛋白质命名为rpprc1。
[0175]
实施例2、用大豆rpprc1基因转化大豆
100mg/l,草铵膦8mg/l。倒入直径9cm培养皿中备用。
[0190]
生根液:吲哚丁酸(indolebutyric acid,iba)30mg,溶于10ml清水中。4c保存, 使用时稀释1000倍。
[0191]
转化流程如下:
[0192]
种子消毒:取干净种子,平铺于9cm培养皿内1-2层,将培养皿置于300ml干燥 器内,干燥器内另放一个烧杯,向烧杯内加入3%次氯酸钠溶液10ml,再加入15%盐 酸溶液10ml,盖上干燥器盖,封闭16-20小时,取出装有种子的培养皿,放置超净台 上约30分钟吹去氯气,培养皿中加入约40ml无菌水,8-12小时后即可进行转化。本 步骤以及下面所述操作如无强调均在无菌条件下操作。
[0193]
农杆菌制备:取单克隆农杆菌(转入pb2gw7-rpprc1),于含有相应抗生素的农 杆菌培养基培养皿上划线,加入3ml液体农杆菌培养基,轻轻旋转培养皿使之铺满培 养皿,28℃条件下培养过夜。第二天用共培养培养基洗下菌苔,制成od值约0.8-1.2 的菌悬液备用。
[0194]
外植体制备:垂直切种子的下胚轴,沿下胚轴中线平均分离两片子叶。子叶和下 胚轴结合处划伤,并移除子叶节处的真叶,每个种子可制成两个外植体;
[0195]
侵染及共培养:外植体放置于菌悬液中,确保所有外植体都浸泡在共培养培养基 中,20-40min后,用移液器移去菌液。共培养平皿(15cm直径)中放置2张无菌圆 形滤纸,滤纸直径略小于培养皿直径(约13-14cm),每皿加入共培养培养基10ml。 将侵染后的外植体平铺于滤纸上,切口向上。培养皿封口,置于22℃18h光周期下培 养5d。
[0196]
再生芽诱导:共培后的外植体切去伸长的下胚轴,放置于再生芽诱导培养基中, 下胚轴区域没入培养基中,每皿(9cm直径)6-7个外植体。在24℃18h光周期下培 养。两周后切去下胚轴部分长出的愈伤,转入新鲜的再生芽诱导培养基中,继续培养 两周。
[0197]
再生芽伸长:将子叶从外植体移除,切除下胚轴新生的愈伤,外植体转入再生芽 伸长培养基中,每两周继代一次,每次转移时都去除下胚轴处新生的愈伤。当芽伸长 到3cm以上时,将伸长的芽(>3cm)从外植体上切除,剩余外植体继续在再生芽伸长 培养基中进行培养。
[0198]
再生芽生根(此步骤可在开放条件下操作):取一空平皿(15cm直径),放一张 直径略小于平皿直径滤纸,加水将纸彻底浸润,将伸长的芽末端浸入3mg/l iba溶液 中10-20秒,取出后用吸水纸包裹好末端,平铺于已浸润的滤纸上,盖上平皿,在24 ℃18h光周期下培养,每天打开盖子换气,并适当补水保持滤纸湿润,待新生根长至 2-3cm时,转入土壤中在温室培养至结实。
[0199]
实验同时设置向大豆品种天隆一号中导入pb2gw7空载体的对照(简称空载对 照)。
[0200]
三、转基因大豆鉴定
[0201]
1、pcr鉴定
[0202]
随机选取2个t0代转基因rpprc1大豆(称为转化事件l1和转化事件l2)的部 分t1代单株(6株,分别记为l1-1、l1-2、l1-3、l2-1、l2-2和l2-3,l1-1、l1-2、 l1-3为转化事件l1的t1单株,l2-1、l2-2和l2-3为转化事件l2的t1代单株), 以及非转基因天隆一号的植株(阴)与zyrc1植株(阳),分别提取基因组dna, 用实施例1的引物f和引物r对目的基因rpprc1进行pcr扩增,目的产物大小为 2574bp,将扩增产物进行1%琼脂糖凝胶电泳,将获
得2574bp条带的植株记为阳性。
[0203]
结果:l1-1、l1-2、l1-3、l2-1、l2-2和l2-3植株全部为阳性,非转基因植株为 阴性,zyrc1植株为阳性,见图2。
[0204]
2、rt-pcr鉴定
[0205]
取步骤二获得的t
1
代转基因rpprc1的大豆(即步骤1中的l1-2和l2-1),空载 转化体的大豆株系(ck)和非转基因植株天隆一号,分别提取总rna,反转录获得 cdna,以该cdna为模板,用特异引物f1和r1对基因rpprc1的cdna进行实时 荧光定量pcr扩增,以大豆β-actin为内参,引物为fc和rc。实时荧光定量pcr在 cfx connect
tm
实时荧光定量pcr仪上进行,一次平行试验设3次重复。利用livak kj 和schmittgen td(2001)报道的方法,即2-δδct
计算相对表达量。
[0206]
δδc
t
=(c
t.target
-c
t.actin
)
time x
-(c
t.target
-c
t.actin
)
time 0
[0207]
time x表示任意时间点,time 0
表示经β-actin校正后1倍量的目标基因表达。
[0208]
上述引物的序列如下:
[0209]
f1:5
’-
tcggcaaagttggttttcatct-3
’
(seq id no.5);
[0210]
r1:5
’-
ccattcctgggctccacatt-3
’
(seq id no.6)。
[0211]
fc:5
’-
attggactctggtgatggtg-3
’
;
[0212]
rc:5
’-
tcagcagaggtggtgaacatt-3
’
。
[0213]
结果如图3显示,非转基因天隆一号和空载植株中基本不表达目的基因rpprc1; 而转基因rpprc1的天隆一号l1-2和l2-1中高表达目的基因rpprc1。
[0214]
3、southern检测
[0215]
取步骤二获得的t1代转基因rpprc1的大豆植株(l1-1、l1-2、l1-3、l1-4、l1-5、 l2-1和l2-1)、非转基因天隆一号的植株(阴),以及转空载的t1代植株l3-1、l3-2、 l3-3、l3-4和l3-5,分别提取基因组dna,内切酶hindiii对基因组dna进行酶切, 采用地高辛杂交检测试剂盒ii(化学发光法)进行southern检测,采用bar基因为探 针,探针引物如下:
[0216]
f2:5'-agaaacccacgtcatgccagtt-3';
[0217]
r2:5'-atcgtcaaccactacatcg-3'(421bp)。
[0218]
结果如图4显示,植株l1-1、l1-2、l1-3、l1-4和l1-5为双拷贝,l2-1和l2-1 为单拷贝,空载植株l3-1、l3-2、l3-3、l3-4和l3-5为单拷贝。对于rpprc1转基 因大豆而言,其可代表rpprc1基因拷贝数。
[0219]
实施例3、检测转基因大豆的抗锈病功效
[0220]
本实施例将用大豆锈菌生理小种ss4(记载于“单志慧等.大豆锈菌在不同豆科 寄主上的致病反应.中国油料作物学报,2008,30(4):497-500”一文,公众可从申请人处 获得,仅可用于重复本发明实验使用,不得他用)接种实施例2获得的转基因植物, 并且对植物病害症状进行评分来测试rpprc1基因的抗锈病功效。
[0221]
首先,对t0转化事件l1和l2进行初步测试,以评估rpprc1转基因对锈病感 染的效应。具体操作如下:取t0植物上完全展开的新叶片,用1
×
10
5
个孢子/ml的大 豆锈菌生理小种ss4菌悬液进行喷雾接种,接种量为每平方厘米为10μl,以未转化的 受体基因型天隆一号(阴性对照)以及来自相同事件的空载植物(即空载对照)为感 病对照,未转化的抗病品种zyrc1为抗病对照(阳性对照)。接种后植株于25℃温室, 光周期为16日照/8小时黑暗,相
对湿度在65-85%下培养,接种后12-15天对病害症 状进行评分。根据病斑性质以及孢子堆破裂情况判断植株的抗病性。将植物定性评级 为免疫(im;没有病变)、高抗(r;产生红黑色病斑,少量孢子形成)和感病(s; 产生黄褐色病斑,大量孢子形成)。参考“bromfield,kr.,melching,js.,kingsolver,ch.. 1980,virulence and aggressiveness of phakopsora pachyrhizi isolates causing soybean rust. phytopathology.70:17-21”一文。结果显示:阳性对照与转化植株(t0转化事件l1和 l2)均表现为免疫,空载与阴性对照表现为感病,如图5和表1。
[0222]
接着,对rpprc1基因抗大豆锈菌的功效进行t1代转基因植株l1-1、l1-2、l1-3、 l2-1和l2-1,空载转化植株l3-1测试。在生长室条件下种植t1代种子,待植株长至 两片真叶完全展开时进行接种鉴定。用大豆锈菌生理小种ss4的孢子悬浮液进行接种。 接种方法同上,未转化品种天隆一号植株为感病对照,未转化抗病品种zyrc1为抗 病对照。12天后观察症状。结果显示:在转基因植株植株l1-1、l1-2、l1-3、l2-1 和l2-1的叶片上没有观察到病斑形成和孢子堆形成,抗性等级为免疫,分子检测其包 含rpprc1基因全长,且具有高水平表达。未转化植株叶片和空载转化植株l3-1的叶 片产生包含黄褐色病斑,产生大量孢子,表现为感病反应(图6和表2)。
[0223]
这些锈病感染测定结果表明,rpprc1基因能够通过转基因的方式提供对大豆锈 菌的抗性。
[0224]
表1.携带rpprc1基因的t0代转化植株对锈病的抗性表现
[0225][0226][0227]
表2.携带rpprc1基因的t1代转化植株对锈病的抗性表现
[0228]
转化植株抗性l1-1免疫l1-2免疫l1-3免疫l2-1免疫l2-2免疫l3-1空载感病阴性对照天隆一号感病阳性对照zyrc1免疫
起点商标作为专业知识产权交易平台,可以帮助大家解决很多问题,如果大家想要了解更多知产交易信息请点击 【在线咨询】或添加微信 【19522093243】 与客服一对一沟通,为大家解决相关问题。
与客服一对一沟通,为大家解决相关问题。
此文章来源于网络,如有侵权,请联系删除

 热门咨询
热门咨询




tips