
 商标分类
商标分类  商标转让
商标转让 
一种海栖热袍菌纤维寡糖转运蛋白、其编码基因及应用的制作方法
 2021-02-02 12:02:11|
2021-02-02 12:02:11| 315|
315| 起点商标网
起点商标网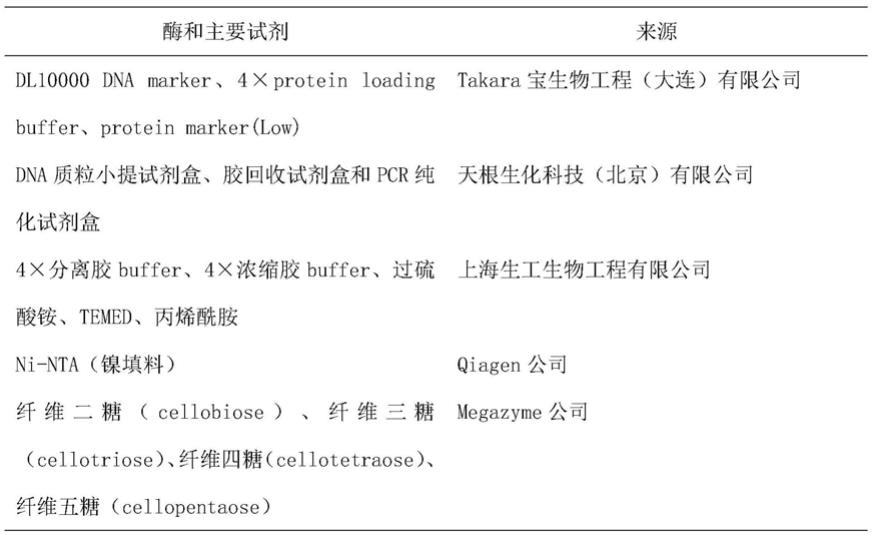
[0001]
本发明涉及生物技术领域,具体涉及一种海栖热袍菌纤维寡糖转运蛋白、其编码基因及应用。
背景技术:
[0002]
随着化石能源的日趋枯竭以及人们对环境污染、能源危机等问题关注度的提高,寻找清洁的可再生能源成为当前的发明热点。木质纤维素是由多种有机高分子化合物组成的具有高结晶度和聚合度的复合体,主要由纤维素、半纤维素和木质素通过共价键和非共价键相互连接形成,因其储量丰富、成本低廉、清洁环保而受到各国科学家的青睐。
[0003]
纤维素酶,包含外切-1,4-β-d-葡聚糖酶(exo-β-glμcanases,cbh)、内切-1,4-β-d-葡聚糖酶(endo-β-glμcanases,eg)、β-葡萄糖苷酶(endo-β-glμcosidases,ge),可以将纤维素、半纤维素水解为单糖,其中外切-1,4-β-d-葡聚糖酶从纤维素的两端水解结晶纤维素,释放纤维二糖;内切-1,4-β-d-葡聚糖酶作用于纤维素内部的β-1,4糖苷键,随机水解无定形纤维素,释放各种纤维寡糖;β-葡萄糖苷酶也称纤维二糖酶,主要水解纤维二糖、纤维寡糖,释放葡萄糖。纤维二糖对外切-1,4-β-d-葡聚糖酶和内切-1,4-β-d-葡聚糖酶有抑制作用,而葡萄糖对β-葡萄糖苷酶有抑制作用,当水解液中的葡萄糖浓度升高时,纤维素的水解效率下降。因此,较低的木质纤维素酶解效率,较高的纤维素酶使用成本是制约木质纤维素这一可再生能源开发利用的瓶颈。
[0004]
直接利用木质纤维素预处理液中的纤维寡糖是有效降低纤维素酶解和发酵成本,实现木质纤维素高效发酵转化的途径之一。但是,因为纤维素结构复杂以及纤维寡糖是大分子物质,不能以自由扩散等简单的扩散方式通过细胞膜,必需通过转运蛋白的协助。在目前的工业发酵微生物菌株中,没有纤维寡糖转运蛋白,不具有直接利用纤维寡糖进行发酵的能力。
技术实现要素:
[0005]
本发明公开了一种海栖热袍菌纤维寡糖转运蛋白、其编码基因及应用,以解决上述现有技术存在的问题。
[0006]
为实现上述目的,本发明提供了如下方案:
[0007]
本发明提供一种海栖热袍菌纤维寡糖转运蛋白,所述海栖热袍菌纤维寡糖转运蛋白的氨基酸序列如seq id no.1所示。
[0008]
本发明提供一种海栖热袍菌纤维寡糖转运蛋白的编码基因,所述编码基因的dna序列如seq id no.2所示。
[0009]
本发明提供一种制备海栖热袍菌纤维寡糖转运蛋白的方法,具体包括如下步骤:
[0010]
(1)pet-tma-cbta重组表达质粒的构建;
[0011]
(2)重组底物结合蛋白的表达:将步骤(1)所述重组表达质粒转化至大肠杆菌中,培养基因工程菌、诱导重组底物结合蛋白表达;
[0012]
(3)纯化步骤(2)所得的重组底物结合蛋白;
[0013]
(4)将步骤(3)所得纯化融合蛋白的洗脱液透析除盐,得到纯的重组底物结合蛋白。
[0014]
进一步地,步骤(2)所述基因工程菌在含卡那霉素以及氯霉素的培养基中培养,采用的诱导剂为iptg。
[0015]
进一步地,所述基因工程菌的转化方法为氯化钙-氯化镁法。
[0016]
进一步地,所述纯化重组底物结合蛋白的方法为亲和层析法。
[0017]
本发明还提供一种海栖热袍菌纤维寡糖转运蛋白在纤维素发酵中的应用。
[0018]
进一步地,所述的纤维素为纤维寡糖。
[0019]
进一步地,所述的纤维寡糖包括纤维二糖、纤维三糖、纤维四糖、纤维五糖中的一种或多种。
[0020]
进一步地,所述的纤维寡糖为纤维五糖。
[0021]
本发明公开了以下技术效果:
[0022]
本发明通过将海栖热袍菌纤维寡糖转运蛋白基因转入感受态目的菌株并进行诱导表达培养,成功构建了高效表达海栖热袍菌纤维寡糖转运蛋白的工程菌株,并通过热力学实验证明了其能够高效结合纤维寡糖,尤其是纤维五糖,为产业上进一步利用纤维素提供了工具。
附图说明
[0023]
图1是牛血清蛋白标准曲线;
[0024]
图2是纯化的tma-cbta底物结合蛋白进行的sds-聚丙烯酰胺凝胶电泳分析;
[0025]
图3是tma-cbta底物结合蛋白与葡萄糖以及各种纤维寡糖相互作用的微量热差示扫描量结果;图3-a是tma-cbta的dsc分析,图3-b是tma-cbta与葡萄糖的dsc分析,图3-c是tma-cbta与纤维二糖的dsc分析,图3-d是tma-cbta与纤维三糖的dsc分析,图3-e是tma-cbta与纤维四糖的dsc分析,图3-f是tma-cbta及其与纤维五糖的dsc分析;
[0026]
图4是纤维寡糖和底物结合蛋白tma-cbta在50℃等温滴定的曲线;图4-a是纤维二糖滴定tma-cbta,图4-b是纤维三糖滴定tma-cbta,图4-c是纤维四糖滴定tma-cbta,其中图4-d纤维五糖滴定tma-cbta。
具体实施方式
[0027]
下面结合附图进一步说明本发明的实施例,该详细说明不应认为是对本发明的限制,而应理解为是对本发明的某些方面、特性和实施方案的更详细的描述。下述实施例中所使用的实验方法,如无特别说明,均为常规方法。
[0028]
实验材料
[0029]
1、菌株和质粒
[0030]
克隆宿主escherichia coli top10、表达宿主escherichia coli rosetta(de3)、和表达载体pet-28a均由本实验室保存。
[0031]
2、酶和主要试剂
[0032]
本发明使用的酶和主要试剂见表1。
[0033]
表1酶和主要试剂
[0034][0035][0036]
3、培养基和抗生素的配制
[0037]
各培养基的配制成分为:
[0038]
lb培养基:1%tryptone(w/v)、0.5%yeast extract(w/v)、1%nacl(w/v),用去离子水定容,分装后于121℃灭菌20min;
[0039]
la培养基:在lb培养基中加入1.8%(w/v)的琼脂粉,121℃灭菌20min;
[0040]
sob培养基:2%tryptone(w/v)、0.5%yeast extract(w/v)、0.05%nacl(w/v)、2.5mmol/lkcl、10mmol/lmgcl,121℃灭菌20min,另外,配制1mol/l的葡萄糖溶液,115℃灭菌15min;
[0041]
soc培养基:在超净工作台里,向sob培养基中加入灭菌后的葡萄糖溶液,使葡萄糖终浓度为10mmol/l,混匀后,分装到1.5ml的ep管,1ml/管,放-20℃冷藏;
[0042]
m9盐溶液:0.0855%na
2
hpo
4
·
12h
2
o(w/v)、0.015%kh2po4(w/v)、0.0025%nacl(w/v)、0.005%nh
4
cl(w/v);
[0043]
另外,单独配制1mol/l硫酸镁溶液、0.1mol/l氯化钙溶液,2%(w/v)纤维二/三/四/五糖溶液,121℃灭菌20min;
[0044]
m9培养基:20%5
×
m9盐溶液(v/v)、0.2%1mol/l硫酸镁溶液(v/v)、0.1%0.1mol/l氯化钙溶液(v/v);
[0045]
在超净工作台里,将上述溶液混匀,加硫酸镁和氯化钙溶液时,及时混匀,以免形成沉淀。
[0046]
本发明所使用的培养基均采用121℃20min湿热高压灭菌,各培养基根据需要,在使用前加入相应的抗生素,固体培养基含1.8%(w/v)琼脂粉。本发明所用抗生素见表2,将抗生素贮存液分装后放-20℃冷藏。
[0047]
表2抗生素溶液
[0048][0049]
4、仪器和设备
[0050]
本发明所用仪器和设备见表3。
[0051]
表3仪器设备
[0052][0053]
实施例1重组底物结合蛋白tma-cbta的制备
[0054]
1、pet-tma-cbta重组表达质粒构建
[0055]
对来自海栖热袍菌(thermotoga maritima)的基因序列进行blastx比对分析,并对其编码的氨基酸序列进行蛋白质结构域预测和信号肽分析,将其命名为tma-cbta(如seq id no.2所示)。根据对基因序列的分析,将编码信号肽的序列去掉以后,由科雷生物科技有限责任公司进行密码子优化并构建到pet-28a表达载体上。
[0056]
2、从e.coli中小量制备质粒
[0057]
(1)取1.5ml过夜培养的菌液加入容积为2ml的ep管,12000rpm离心1min,弃去上清。
[0058]
(2)向ep管中加入250μl预冷的p1溶液(使用前加入rnase a),使用涡旋振荡器重悬菌体。
[0059]
(3)向ep管中加入250μl预冷的p2溶液,温和地上下翻转6-8次,使菌体充分裂解。
[0060]
(4)向ep管中加入350μl预冷的p3溶液,立刻温和地上下翻转6-8次,充分混匀,12000rpm离心10min。
[0061]
(5)将上清转移到吸附柱cp3中,12000rpm离心1min,弃去收集管中的废液,将吸附柱cp3放回收集管中。
[0062]
(6)向吸附柱cp3中加入500μl去蛋白液pd,12000rpm离心1min,弃去收集管中的废液,将吸附柱cp3放回收集管中。
[0063]
(7)向吸附柱cp3中加入600μl去漂洗液pw,12000rpm离心1min,弃去收集管中的废液,将吸附柱cp3放回收集管中。
[0064]
(8)重复步骤7。
[0065]
(9)将吸附柱cp3放回收集管中,12000rpm离心2min,将吸附柱cp3转移到一个干净的ep管,向吸附膜中间部位加入50μl无菌水,室温静置2min,12000rpm离心2min,将质粒溶液收集到ep管,放-20℃存储备用。
[0066]
3、氯化钙-氯化镁法制备大肠杆菌感受态细胞
[0067]
(1)从-40℃取出甘油保藏的e.coli菌种,在超净工作台里用接种环蘸取菌液,在无抗生素的la平板上划线,放37℃培养过夜。
[0068]
(2)从la平板上挑取生长良好的单菌落,接种到内含5ml lb培养基的直形瓶,37℃,200rpm培养过夜。
[0069]
(3)按1%的接种量(v/v)转接到内含100ml lb培养基的锥形瓶里,37℃,200rpm培养2-3h,菌液od
600
至0.4-0.6。
[0070]
(4)将培养好的菌液使用冰浴进行冷却,冰浴时间约为30min,中间偶有混匀。
[0071]
(5)在超净台里将充分冷却的菌液分装到预冷的、无菌的容积为50ml的聚丙烯管,在4℃下,5000rpm离心10min,弃去上清。
[0072]
(6)向聚丙烯管中加入35ml预冷的氯化钙-氯化镁溶液,轻轻敲打重悬菌体,在4℃下,5000rpm离心5min,弃去上清。
[0073]
(7)重复步骤6两次。
[0074]
(8)向聚丙烯管中加入4ml预冷的甘油-氯化钙溶液,轻轻敲打重悬菌体,在冰上将感受态细胞分装到无菌的、预冷的ep管,每管100μl,放-80℃冷藏。
[0075]
4、dna分子转化大肠杆菌感受态细胞
[0076]
(1)从-80℃取出大肠杆菌感受态细胞,迅速插入冰里,让大肠杆菌感受态细胞在冰里自然融化。
[0077]
(2)用微量移液器吸取50-100ng的dna分子溶液加入内含100μl大肠杆菌感受态细胞的ep管中,混匀后冰浴静置30min。
[0078]
(3)将ep管快速地放入42℃水浴,在静置的条件下,热激90s。
[0079]
(4)热激结束后立即取出ep管,冰浴静置3min。
[0080]
(5)向ep管内加入200μl soc培养基,37℃,200rpm培养1h。
[0081]
(6)取适量体积的培养物涂布到含有100μg/ml氨苄青霉素的la平板上,放37℃恒
温培养箱倒置培养12-16h。
[0082]
5、重组底物结合蛋白的诱导表达
[0083]
(1)从转化平板上挑取生长良好的单菌落到5ml的lb培养基(含50μg/ml卡那霉素)的直形瓶中,37℃、200rpm培养10h。
[0084]
(2)按1%(v/v)的接种量分别转接到内含65ml的lb培养基(含50μg/ml卡那霉素)的锥形瓶中,37℃、200rpm培养过夜。
[0085]
(3)按1%(v/v)的接种量分别转接到内含1l lb培养基(含50μg/ml卡那霉素)的锥形瓶中,37℃、200rpm培养2-3h,菌液od
600
至0.4-0.6时加入iptg至终浓度为1mmol/l。
[0086]
(4)将重组菌株e.colirosetta(de3)/pet-tma-cbta放37℃、200rpm培养10-12h进行目的蛋白的诱导表达。
[0087]
6、重组底物结合蛋白tma-cbta的纯化
[0088]
(1)诱导菌液表达后,8000rpm离心10min,收集菌体。
[0089]
(2)用300ml lysis buffer(ph8.0,咪唑10mmol/l,氯化钠300mmol/l,磷酸二氢钠50mmol/l)重悬菌体,加入3ml 25mg/ml的溶菌酶,混匀后冰上静置20min。
[0090]
(3)在冰水混合物中使用超声波细胞破碎仪进行破胞,破胞总时间20min,工作时间5s,间歇时间5s,破胞功率30%。
[0091]
(4)破胞结束后,重组菌株e.colirosetta(de3)/pet-tma-cbta的破胞液放70℃水浴40min,水浴结束后于4℃,12000rpm离心30min,离心后的上清液为粗蛋白液。
[0092]
(5)吸取2ml ni-nta填料至层析柱中,用lysis buffer(ph8.0,咪唑10mmol/l,氯化钠300mmol/l,磷酸二氢钠50mmol/l)充分平衡层析柱中的ni-nta填料。
[0093]
(6)将粗蛋白液与用lysis buffer平衡的ni-nta填料混合,放10℃,150rpm作用1h,使含有组氨酸标签的重组底物结合蛋白与ni-nta填料充分结合。
[0094]
(7)将混合液转移到层析柱中并排空液体,向层析柱中加入10ml预冷的wash buffer(ph8.0,氯化钠300mmol/l,磷酸二氢钠50mmol/l,咪唑浓度为40mmol/l),轻轻重悬ni-nta,排空层析柱内的液体,重复4次。
[0095]
(8)充分洗涤以后向层析柱中加入10ml预冷的elution buffe(ph8.0,咪唑250mmol/l,氯化钠300mmol/l,磷酸二氢钠50mmol/l)洗脱底物结合蛋白,收集洗脱液,重复3次,4℃保存含有底物结合蛋白的洗脱液。
[0096]
(9)底物结合蛋白洗脱结束以后,使用elution buffer多次冲洗ni-nta填料,冲洗结束后用20%的乙醇溶液保存。纯化同一种底物结合蛋白时,ni-nta填料可重复使用3-5次,结合率下降以后可对ni-nta填料进行再生。
[0097]
7、底物结合蛋白tma-cbta的脱盐
[0098]
(1)将含有底物结合蛋白tma-cbta的洗脱液转移到30kda的超滤管,洗脱液较多时可分多次转移,在4℃条件下,5000rpm离心30min,弃去收集管中的废液。
[0099]
(2)超滤管中的洗脱液约为500μl时,向超滤管中加入buffer a溶液(ph7.0,50mm tris-hcl,100mm nacl)在4℃条件下,5000rpm离心30min,弃去收集管中的废液。
[0100]
(3)重复步骤2两次。
[0101]
(4)将超滤管中含有底物结合蛋白的溶液和收集管中的超滤液转移到干净的ep管和直形瓶,放4℃保存。
[0102]
8、底物结合蛋白tma-cbta标准曲线的绘制和定量分析
[0103]
吸取100μl牛血清蛋白bsa标准品(2mg/ml),加入1900μl ddh
2
0后充分混匀,按照表4在1.5ml的ep管内用稀释液(ddh
2
o、0.9%nacl或者pbs)稀释bsa标准品,每个浓度三个平行。
[0104]
表4牛血清蛋白标准曲线测定
[0105][0106]
分别吸取20μl稀释后的bsa标准品溶液加入1.5ml的ep管,每个浓度做三个平行,分别向每管加入1ml复温的broadford dye reagent,混匀后室温反应5min,各取200μl加入96孔板,使用酶标仪测定600nm处的吸光度值,计算各浓度的平均值后减去blank的平均值,以蛋白质含量为横坐标,od
600
的读数为纵坐标,绘制bsa标准品溶液的标准曲线,如图1所示。
[0107]
纯化的底物结合蛋白tma-cbta定量:分别取20μl不同稀释倍数的目的蛋白溶液加入1ml复温的broadford dye reagent,混匀后室温反应5min,各取200μl加到酶标板,读取od
600
的吸光度值,代入标准曲线计算目的蛋白的质量浓度,结果得浓度为8.55469mg/ml。
[0108]
9、底物结合蛋白tma-cbta的sds-page分析
[0109]
按照标准sds-page方法(分离胶浓度10%,浓缩胶浓度5%)分析底物结合蛋白的表达情况与纯化的底物结合蛋白的分子量和纯化程度,经考马斯亮蓝染色以及聚丙烯酰胺凝胶充分脱色以后,使用成像仪gel doc
tm xr+获得理想的sds-聚丙烯酰胺凝胶电泳图。如图2所示,对诱导后的重组菌株分别进行超声破胞纯化,在相应分子量大小处出现了明显的蛋白质特征性条带,tma-cbta在70℃水浴1h后进行纯化,杂蛋白已经明显减少,说明我们已经成功表达了该底物结合蛋白tma-cbta(如seq id no.1所示)。
[0110]
实施例2底物结合蛋白的热力学和底物亲和力分析方法
[0111]
1、微量热差示扫描量热法(dsc)
[0112]
本发明对底物结合蛋白tma-cbta分别做了与葡萄糖、纤维二糖、纤维三糖、纤维四糖、纤维五糖的微量热差示扫描实验。
[0113]
(1)用最后一次收集管中的超滤液将底物结合蛋白溶液稀释至2mg/ml并配制浓度为6mmol/l或者8mmol/l的各种纤维寡糖溶液。
[0114]
(2)按1:1(v/v)分别将底物结合蛋白溶液和纤维寡糖溶液混匀,则底物结合蛋白的终浓度为1mg/ml,各种纤维寡糖的终浓度为3mmol/l或者4mmol/l,于4℃真空脱气10—
20min。
[0115]
(3)分别各吸取400μl混合液加入96孔板的偶数孔,每个样品两个平行,每个样品前面的奇数孔和最前面的两个偶数孔为400μl真空脱气的buffer a,随后紧跟的两个偶数孔为空白对照,即400μl真空脱气的超滤液。检测结束后用5%decon 90清洗仪器。
[0116]
dsc检测条件:
[0117]
底物结合蛋白tma-cbta:样品槽温度10℃,检测起始温度30℃,终止温度125℃,温度变化率90℃/h,蛋白浓度1mg/ml,各种纤维寡糖浓度4mmol/l,反馈模为none。
[0118]
利用分析软件对底物结合蛋白tma-cbta与纤维寡糖的热力学作用进行分析并计算其变性过程中在溶解温度tm值时的量热焓变(
△
h)、范特霍夫焓变(
△
hv)、熵变(
△
s)、热容变化量(
△
cp)及在tm值时的结合常数(kb)。
[0119]
2、等温滴定量热法(itc)
[0120]
等温滴定量热法可以直接测定底物结合蛋白和纤维寡糖结合时产生的结合焓变化量(
△
h)、熵变化量(
△
s)、结合常数(kb)和化学计量数(n),同时,根据实验测得的热力学参数可以计算出结合反应的吉布斯自由能变化量(
△
g)和转变温度(t转)。使用纤维二糖、纤维三糖、纤维四糖、纤维五糖滴定底物结合蛋白tma-cbta,滴定结果使用一个结合位点模式进行曲线拟合运算分析;
[0121]
(1)用最后一次收集管中的超滤液将底物结合蛋白溶液稀释至0.1-0.2mmol/l并配制浓度为底物结合蛋白浓度10倍左右的各种纤维寡糖溶液。
[0122]
(2)于4℃真空脱气10—20min,设置等温滴定的各个参数。
[0123]
各个底物结合蛋白的滴定条件:
[0124]
tma-cbta底物结合蛋白的滴定浓度为0.1mmol/l,各种纤维寡糖的滴定浓度为1mmol/l,进样针体积为40μl,进样针抽滤时间为5s,每次滴定持续时间6s,第1滴体积为0.1μl,第2-20滴体积为1.5μl,共20滴,每两滴的间隔时间为120s,样品池温度为50℃,参考能量值10μcal/sec,起始延长时间1min,进样针搅拌速度500rpm,反馈模式low。
[0125]
底物结合蛋白的热力学参数
[0126]
使用origin itc 200软件进行数据分析和曲线拟合,在一次等温滴定实验中,itc可以测定亲和力常数kb,化学计量数n,结合的焓变
△
h和熵变
△
s,根据实验测得的热力学参数和吉布斯—亥姆霍兹方程可以计算出吉布斯自由能
△
g。
[0127]
3、底物结合蛋白tma-cbta的热力学特性和特异性底物亲和能力分析结果
[0128]
3.1微量热差示扫描量热法对tma-cbta的热力学分析
[0129]
按照步骤1的方法,使用终浓度为0.015mmol/l的底物结合蛋白tma-cbta以及其与葡萄糖、纤维二糖、纤维三糖、纤维四糖、纤维五糖进行的dsc热力学分析,结果如图3所示,图3是tma-cbta底物结合蛋白与葡萄糖以及各种纤维寡糖相互作用的微量热差示扫描量结果;图3-a是tma-cbta的dsc分析,图3-b是tma-cbta与葡萄糖的dsc分析,图3-c是tma-cbta与纤维二糖的dsc分析,图3-d是tma-cbta与纤维三糖的dsc分析,图3-e是tma-cbta与纤维四糖的dsc分析,图3-f是tma-cbta及其与纤维五糖的dsc分析;
[0130]
如图3所示,结果表明:tma-cbta不结合葡萄糖。而底物结合蛋白tma-cbta与纤维二糖、纤维三糖、纤维四糖、纤维五糖结合以后,tm值比未结合纤维寡糖的tma-cbta要高,说明纤维二糖、纤维三糖、纤维四糖、纤维五糖与tma-cbta结合以后,诱导了tma-cbta内部构
象改变。应用非二态模式对tma-cbta及其与各种纤维寡糖形成复合物以后的微量热差示扫描结果进行分析,热力学参数:溶解温度tm、量热焓变
△
h、范特霍夫焓变
△
h
v
见表5。
[0131]
表5 tma-cbta的dsc热力学参数
[0132][0133]
从表5可知,底物结合蛋白tma-cbta的
△
h﹥
△
hv且
△
h/
△
hv的比值等于4.58,说明tma-cbta为单体蛋白质且内部含有两个以上结构域,结合图3可知,tma-cbta与纤维寡糖的复合物在变性过程中有三个热容扫描峰,说明tma-cbta折叠形成了三个结构域,在变性过程中有两个结构域具有相同或相近的溶解温度tm,在100—110℃之间,另一个结构域的溶解温度较低,在80-90℃之间。
[0134]
除此之外,量热焓与范特霍夫焓之间的关系还可以判断蛋白质热变性过程中蛋白质的转变模式。本发明使用分析软件中的非二态模式对底物结合蛋白tma-cbta及其与纤维寡糖的热变性曲线进行拟合,得到tma-cbta及其与各种纤维寡糖形成的复合物的范特霍夫焓
△
h
v
和量热焓
△
h(具体见表5)。从表5可知,底物结合蛋白tma-cbta及其与各种纤维寡糖形成的复合物的
△
h
v
﹤
△
h,所以,底物结合蛋白tma-cbta的变性过程是非二态转变模式。tma-cbta结合纤维二糖、纤维三糖、纤维四糖、纤维五糖以后,
△
h
v
和
△
h均有不同程度增大,
△
h增大幅度更为明显,
△
h
v
/
△
h比值增大,说明tma-cbta与纤维寡糖之间发生了相互作用,并且tma-cbta与纤维寡糖形成复合物以后,tma-cbta在热变性过程中介于自然状态和变性状态之间的稳定中间态协同去折叠作用加强。结合图3可知,tma-cbta与纤维寡糖形成复合物以后,去折叠的温度范围比天然状态下的tma-cbta更窄,从侧面说明了tma-cbta与纤维寡糖形成复合物以后,tma-cbta的非二态变性过程的协同去折叠作用加强。
[0135]
纤维寡糖与底物结合蛋白的相互作用是分子内部和分子间共同作用的结果,这些相互作用表现在焓变化量
△
h、熵变化量
△
s、吉布斯自由能变化量
△
g。通常地,tm值是衡量蛋白质热稳定性的一项重要指标,tm值越高,蛋白质的热稳定性越好。当纤维寡糖只结合天然状态的底物结合蛋白时,形成的复合物会更稳定,与未结合纤维寡糖的天然状态的底物结合蛋白相比,该复合物具有较高的tm值;若纤维寡糖只结合去折叠状态的底物结合蛋白时,形成的复合物稳定性降低,与未结合纤维寡糖的天然状态的底物结合蛋白相比,该复合物的tm值比未结合纤维寡糖的天然状态的底物结合蛋白的tm值要小。结合图3和表5可知,纤维二糖、纤维三糖、纤维四糖只结合天然状态的底物结合蛋白tma-cbta,所以,纤维二糖、纤维三糖、纤维四糖与tma-cbta形成的复合物稳定性提高,各自的tm值较未结合纤维寡糖
的天然状态的底物结合蛋白tma-cbta升高。
[0136]
3.2等温滴定量热法对tma-cbta与相应特异性底物的亲和能力分析
[0137]
按照步骤2方法,使用终浓度为1mmol/l的纤维寡糖溶液分别滴定底物结合蛋白tma-cbta,测定tma-cbta的结合热力学特性,结果如图4所示,图4-a是纤维二糖滴定tma-cbta,图4-b是纤维三糖滴定tma-cbta,图4-c是纤维四糖滴定tma-cbta,其中图4-d纤维五糖滴定tma-cbta。
[0138]
从图4可知,tma-cbta可与纤维二糖、纤维三糖、纤维四糖、纤维五糖结合,等温滴定量热法的结果与微量热差示扫描量热法结果一致。使用一个结合位点模式对滴定结果进行拟合分析,各结合热力学参数见表6。
[0139]
表6底物结合蛋白tma-cbta与纤维寡糖相互作用的热力学参数
[0140][0141][0142]
从表6可知,tma-cbta与纤维二糖、纤维三糖、纤维四糖、纤维五糖均有结合,对纤维五糖亲和力最强,结合常数k
b
为(2.25
±
0.88)
×
10
5
m-1
,对纤维三糖亲和力最弱,结合常数k
b
为(1.09
±
0.04)
×
10
5
m-1
。
[0143]
由表6可知,tma-cbta与各种纤维寡糖的结合熵变化量(
△
s)均小于0,说明tma-cbta和各种纤维寡糖的结合反应在本实验条件下为自发的正向不可逆过程;结合焓变化量(
△
h)均小于0,说明tma-cbta和各种纤维寡糖的结合反应在本实验条件下为放热反应;
△
s、
△
h、
△
g均小于0,由此可以判断,在本实验条件下,底物结合蛋白tma-cbta与各种纤维寡糖的结合反应是由焓驱动的正向自发进行的不可逆反应,温度t起能动作用。纤维二糖、纤维三糖、纤维四糖、纤维五糖与底物结合蛋白的转变温度依次为599.82(k)、649.02(k)、579.70(k)、1564.96(k),均比滴定温度t(323.15k)大,从表6可知,
△
h﹤0、
△
s﹤0均小于0,所以反应正向自发进行。
[0144]
从表6和表7可知,tma-cbta对纤维五糖的亲和力最强,在50℃测得其解离常数k
d
为4.81μm,纤维四糖次之,解离常数为6.25μm,然后是纤维二糖,解离常数为7.13μm,对纤维三糖的亲和力最弱,解离常数为9.18μm。各自的吉布斯自由能也都是负值,说明各个复合物也是稳定的,其复合物的稳定性由大到小依次为:g5-tma-cbta(-7.89kcal/mol)﹥g4-tma-cbta(-7.71kcal/mol)﹥g2-tma-cbta(-7.61kcal/mol)﹥g3-tma-cbta(-7.45kcal/mol),从表7可知,tma-cbta与各种纤维寡糖形成的复合物中,对纤维五糖和纤维四糖有较小的解离常数和较大的负数吉布斯自由能,而对分子量较小的纤维二糖、纤维三糖的亲和力较弱。
[0145]
3.3底物结合蛋白的热力学特征
[0146]
每次微量热差示扫描的样品,纤维寡糖浓度比底物结合蛋白浓度要高,确保天然状态的底物结合蛋白的每个结合位点都被饱和。根据以上分析可知,纤维寡糖只与天然状态的底物结合蛋白结合,因此,dsc可以分析底物结合蛋白与纤维寡糖在tm值时的结合焓
△
h
b
°
、结合熵
△
s
b
°
,结合焓可由以下等式获得:
[0147]
△
h
b
°
=
△
h
°
(tm)-[
△
h
°
m+
△
c
p
(tm
–
t
°
m)]
[0148]
△
h
°
(tm)存在纤维寡糖的情况下,实验测得的纤维寡糖与底物结合蛋白在tm值时的焓变;
△
h
°
m、
△
c
p
在不含纤维寡糖的情况下,实验测得的底物结合蛋白在t
°
m值时的焓变和热容变化量。在tm值时
△
g
b
°
=0,结合熵由吉布斯-亥姆霍兹方程导出:
[0149]
△
g
b
°
=-rt lnk
eq
=
△
h
b
°
-t
△
s
b
°
[0150]
底物结合蛋白tma-cbta与相应纤维寡糖的结合焓
△
h
b
°
、结合熵
△
s
b
°
见表7。
[0151]
随着温度的不断升高,蛋白质的天然构象逐渐发生改变,溶液中同时存在天然构象的蛋白质和去折叠的蛋白质,当温度t升高至tm时,天然蛋白质和变性蛋白质处于平衡状态,此时,体系的吉布斯自由能
△
g
°
m=0,根据吉布斯-亥姆霍兹方程有
△
h
°
m=tm
△
s
°
m,底物结合蛋白tma-cbta及其与相应纤维寡糖形成复合物后的
△
s
°
m。
[0152]
焓、熵均是温度的状态函数,因此,当温度t改变时,吉布斯自由能不可以直接积分,其它任意温度下的吉布斯自由能、焓变和熵变可根据修饰的吉布斯-亥姆霍兹方程(the modified gibbs-helmholtzeqμation)和基尔霍夫法则(kirchhoff's law)计算:
[0153][0154]
△
h
°
(t)=
△
h
°
m+
△
c
p
(t
–
tm)
[0155][0156]
每次实验可检测到的热力学参数有转变中点温度tm,在转变中点温度的焓变
△
h
°
m和热容变化量
△
c
p
。体系每升高1k时所需要吸收的热量称为该体系的热容,蛋白质发生变性以后,与天然状态的蛋白质相比,具有更高的热容c
p
。
[0157]
表7底物结合蛋白tma-cbta的dsc热力学参数
[0158][0159]
从表7可知,底物结合蛋白tma-cbta在变性过程中,热容变化量
△
c
p
均大于0,可能与蛋白质变性将其疏水侧链暴露于溶液中有关。
[0160]
3.4底物结合蛋白tma-cbta特异性底物的亲和力分析
[0161]
等温滴定量热法结果表明:来自海栖热袍菌的底物结合蛋白可以结合聚合度更高的纤维寡糖,对纤维二糖至纤维五糖均有不同程度的亲和力,该底物结合蛋白与纤维寡糖的结合参数见表8。
[0162]
tma-cbta:能够结合纤维二糖至纤维五糖,在50℃滴定时,tma-cbta对纤维五糖的亲和力最强,解离常数为4.81
±
1.87μm,对纤维三糖亲和力最弱,解离常数为9.18
±
0.35μ
m。
[0163]
表8底物结合蛋白的热力学参数
[0164][0165]
3.5结论
[0166]
(1)从海栖热袍菌thermotoga maritima中克隆到一个atp结合盒转运蛋白超家族(atp-binding cassette transporters,abc transporters)的底物结合蛋白(substrate-binding protein,sbp)基因,该底物结合蛋白均能够在e.coli中成功重组表达。dsc分析表明底物结合蛋白tma-cbta在天然状态下是以折叠成多个结构域的单体形式存在。
[0167]
(2)tma-cbta的溶解温度是106.3
±
0.06℃,且该底物结合蛋白能够结合纤维二糖、纤维三糖、纤维四糖、纤维五糖。
[0168]
(3)等温滴定量热法的结果表明tma-cbta能够结合纤维二糖、纤维三糖、纤维四糖、纤维五糖,与dsc结果相符,itc的结果显示tma-cbta与纤维五糖亲和力最强,解离常数k
d
为4.81
±
1.87μm,其次为纤维四糖和纤维二糖,解离常数k
d
分别为6.25
±
1.42μm、7.13
±
0.40μm,对纤维三糖亲和力最弱,解离常数k
d
为9.18
±
0.35μm。
[0169]
(4)虽然底物结合蛋白tma-cbta在ncbi中的注解为寡肽底物结合蛋白,但是经过我们验证,该底物结合蛋白能够结合纤维二糖至纤维五糖。
[0170]
以上所述的实施例仅是对本发明的优选方式进行描述,并非对本发明的范围进行限定,在不脱离本发明设计精神的前提下,本领域普通技术人员对本发明的技术方案做出的各种变形和改进,均应落入本发明权利要求书确定的保护范围内。
起点商标作为专业知识产权交易平台,可以帮助大家解决很多问题,如果大家想要了解更多知产交易信息请点击 【在线咨询】或添加微信 【19522093243】 与客服一对一沟通,为大家解决相关问题。
与客服一对一沟通,为大家解决相关问题。
此文章来源于网络,如有侵权,请联系删除

 热门咨询
热门咨询




tips