
 商标分类
商标分类  商标转让
商标转让 
西梅流相防腐保鲜方法与流程
 2021-01-07 10:01:40|
2021-01-07 10:01:40| 610|
610| 起点商标网
起点商标网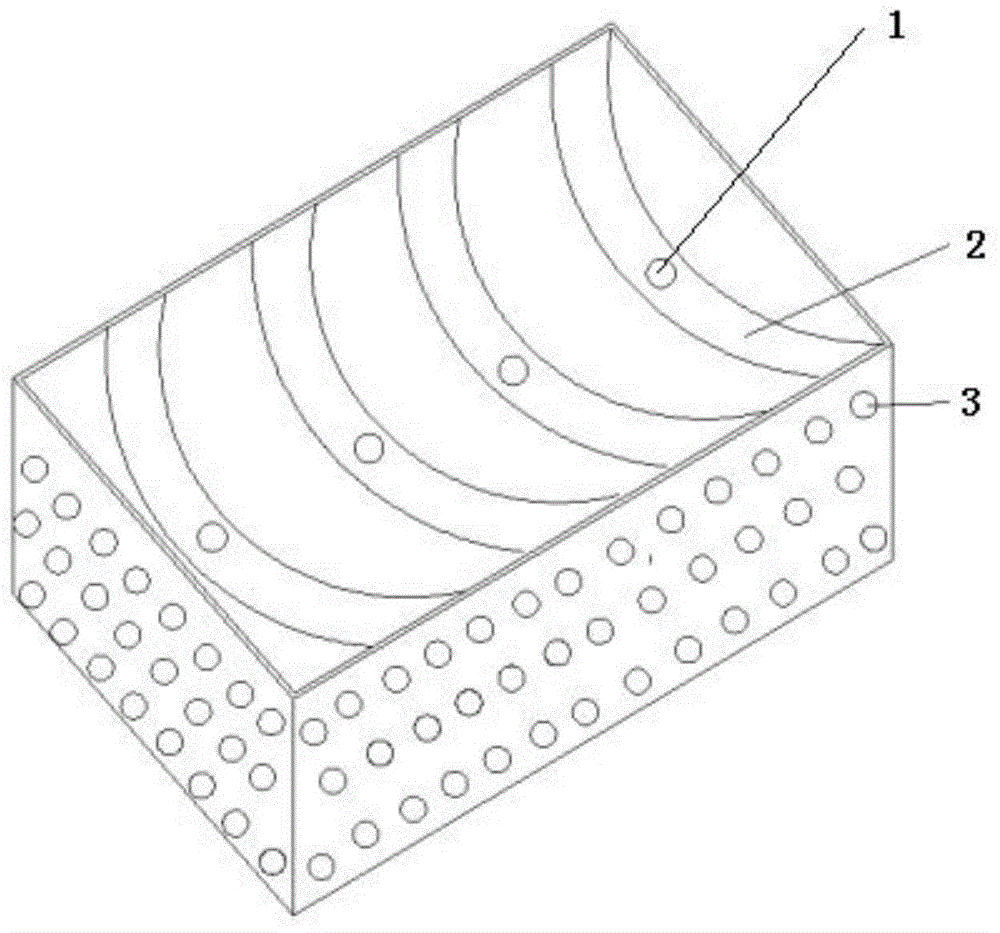
本发明属于水果保鲜领域,涉及西梅,尤其是一种西梅流相防腐保鲜方法。
背景技术:
采后西梅呼吸越变和乙烯释放引起的果实软化,极易失水等问题。采后易遭受侵染性病害,如黑根霉、青霉、曲霉、交链孢霉等。采收标准不统一,采收机械伤大,果实采收、分级、包装和搬运过程中,磕碰、刺划等机械伤在所难免,导致果实伤呼吸、伤乙烯、酶诱导等系列次级应急保护反应加剧,加速褐变、软化、失水。采后预冷效率低,效果差,静止预冷24小时降至0-2℃,失水。冷库气调库贮藏,传热传质、需要不断地“加湿、除霜”导致库温波动大、能耗高,加湿点附件湿度大,腐烂严重,加湿点远端湿度小,失水萎蔫严重,化学保鲜剂浸泡处理残留量高。
cn101595909a提供了一种红枣保鲜的新工艺。其主要技术特征是:红枣经清洗、20℃~150℃清水热烫杀菌3s~1h、风干表面水分,再经浓度为0.2ppm~15ppm臭氧二次杀菌,杀菌时间为1min~4h,然后转入无菌包装车间进行真空包装。本发明是一种在真空包装前杀菌的新工艺,杀菌工艺简单、方便操作、无残留、成本低,保鲜时间长,适用于大规模加工生产。
该专利热处理杀菌过长导致果实损伤,臭氧杀菌时间过长对果实有一定损伤。处理过程温度波动过大。
cn103947746a提供了一种葡萄保鲜或使葡萄抗灰霉病的方法,包括如下步骤:将采后的葡萄果实浸泡在肉桂酸溶液中,以实现葡萄保鲜或抗灰霉病。所述肉桂酸溶液包括肉桂酸和水,所述肉桂酸在所述肉桂酸溶液中的终浓度为1-10mm。本发明的实验证明,本发明利用肉桂酸溶液可以实现采后葡萄果实的保鲜,并达到抗灰霉病的效果。
该专利肉桂酸溶液具有一定的杀菌效果。但是浸泡式保鲜工艺复杂,风干难,导致水分残留,贮藏期易滋生霉菌。
cn104509585a公开了一种葡萄涂膜保鲜方法,解决了现有技术的葡萄保鲜方法会对人体产生危害,安全性差,贮藏后葡萄的品质较差的问题,其包括以下步骤:(1)制备改性纳米sio2;(2)制备复合涂膜保鲜剂;(3)浸剂成膜;(4)预冷保藏。
该专利采用纳米sio2具有一定的抑菌效果。但是涂膜保鲜工艺复杂,成本高;涂膜保鲜的杀菌效果难以保证。
技术实现要素:
本发明的目的在于克服现有技术的不足,提供一种西梅流相防腐保鲜方法,提高西梅贮藏期和贮藏品质。
本发明解决技术问题所采用的技术方案是:
一种西梅流相防腐保鲜方法,包括如下步骤:
(1)采前品质调控;
(2)无伤采收;
(3)简易隧道高湿差压预冷;
(4)相温库流相防腐贮藏,贮藏期间流向风内循环联合tio2光催化处理。
而且,所述的采前品质调控是在采收前15天内在叶面和果实表面喷施品质诱导剂,所述的品质诱导剂由组分a和组分b构成,a组份先喷施,隔天喷施b组份,均稀释300倍,各喷施1~3次;
a组份的制备方法为:按预定要求将l-arg0.1~0.3g/l、亚油酸10~20mmol/l、mesa3.0~8.0mg/l、cys3.0~5.0mg/l溶解在水中,在35~55℃下搅拌15~20min,之后在上述获得的溶液中加入醋酸锌0.2~0.6g/l、氯化钙1~3g/l搅拌均匀,最后在上述溶液中加入sod制剂0.5~1.0g/l定容至1l的混合溶液;
b组份的制备方法为:将br0.1~0.5mg/l、吲哚乙酸0.2~0.4mg/l、赤霉酸10~30mg/l、油菜素内酯0.1~1.0mg/l、meja3~5mg/l、过氧化氢10~20mg/l用少量乙醇溶解后再加入水中,置于35~55℃下搅拌至均匀溶解形成混合插层剂溶液,之后加入硝普钠5~10mmol/l,经30~50khz超声处理10~20min,定容至1l得到插层混合液b。
而且,所述的无伤采收是通过无伤周转箱转运,所述的无伤周转箱的收纳仓主体结构为圆弧形,且中间底部有排水孔,收纳仓内表面为五层复合的保鲜垫,包括依次设置的阻隔层、施药层、载药层、吸附层、防雾层,各层之间采用粘结的方式结合到一起。
而且,所述的阻隔层为无纺布、施药层为微囊化植物精油保鲜剂,载药层为聚丙烯膜或聚乙烯膜中的一种,吸附层由5wt%~8wt%的1-甲基环丙烯、94wt%~91wt%的高锰酸钾和1wt%的聚丙烯酸酯组成,防雾层为淋膜纸。
而且,所述的微囊化植物精油保鲜剂的壁芯比4∶1,芯材为丁香精油:肉桂精油按体积比1:1组成,壁材由50wt%β-环糊精,40wt%壳聚糖,10wt%吐温80组成。
而且,所述的周转箱的箱壁为夹层结构,在侧壁制有可开启式通风孔,夹层内根据实际需要放置蓄冷剂。
而且,所述的简易隧道高湿差压预冷是在冷库内,将西梅裸果装筐码放在托盘上,托盘放在静压箱开孔一侧,紧贴静压箱,依次对齐排放,用pvc布把货堆上面和侧面遮严,风机转动时,在静压箱内产生负压,抽吸冷库中的冷空气,冷空气从货堆尾部穿流进入静压箱,所述的静压箱顶部安装两个轴流风机,静压箱正面开孔,静压箱内安装有加湿器。
而且,预冷过程中同时使用1μl/l1-mcp处理,处理时间12h。
而且,所述的相温库流相防腐贮藏是将西梅放入相温库内,所述相温库为子母夹套结构,蒸发器安装在母库上,蒸发器吹出的冷风通过子库的铝板将冷量传递到子库内,在子库上安装内循环通风系统,采用>1.5m/s的流相风速处理。
而且,所述的流向风内循环联合tio2光催化处理,tio2的粒子比表面积为11.7m2g-1,当量粒径为93.7nm,紫外光波长为365nm,tio2包覆的表面积为592.8cm2,在风速1.5m/s条件下,贮藏期内协同使用流相tio2光催化,每7天处理1次,每次30min,tio2光催化启动时,流相风速内循环同步启动,流相风速内循环每天运行2h。
本发明的优点和积极效果是:
(1)采前品质调控,临采期分子共激发生理病理调节剂,提高果实硬度,抑止呼吸强度和乙烯催熟转红,延缓衰老酒化和失水软化,为提高果实耐藏性和货架期奠定基础。
(2)无伤采收,采用田间无伤包装周转箱,防止机械伤并且具有防腐和拮抗乙烯作用。
(3)使用简易隧道高湿差压预冷装备,快速脱除田间热,使果实采后5-6小时,果温降到5℃以下,高湿预冷协同1-mcp熏蒸处理,减少失水而同时完成防衰老处理。
(4)相温库流相防腐贮藏。首创子母相分控的传热不传质技术体系,突破了不加湿恒湿、蒸发器无霜、精准控温波动<±0.1℃、恒温恒湿气调、脉冲式雾化防腐低残留、流相防腐零残留、原位隧道差压预冷、常闭与常开联动、充分利用自然冷源、充分利用生物能、生物膜零点相变、设备零腐蚀、基于保鲜芯智能控制和互联网+物联网+远程诊断等15大新技术。
(5)本专利首次发现了果蔬贮运环境全封闭条件下空气介质内循环(流相防腐)具有显著防腐作用,当风速>1.5m/s时,黑曲霉菌、灰霉菌、青霉菌和链格孢菌等4种真菌,膜质结构胁迫伤害显著,菌落生长抑制显著,延缓孢子萌发,流相风速导致分生孢子形状变异和细胞体系异常。
(6)1.5m/s流相风速内循环联合tio2光催化处理,抑制了西梅贮藏过程的呼吸速率和乙烯产量,降低了ppo和pod酶活性,提高了保鲜效果,西梅保鲜期超过60天,腐烂率仅3.2%,对照组20.5%。
附图说明
图1为无伤周转箱结构图;
图2为复合保鲜垫的结构示意图;
图3为静压箱结构示意图;
图4为双垛堆纵向无间隙差压预冷物摆放方式效果图;
图5为6米双堆垛纵向无间隙差压预冷降温曲线图;
图6为并联相温库平面图;
图7为图6的a-a向截面剖视图;
图8为图6的b-b向截面剖视图;
图9为相温库(a,b)和冷库(c,d)在0.5m和3.5m高度处的温度场分布图;
图10为不同风速条件对病原真菌菌落直径的影响(3d);
图11为不同风速条件对病原真菌菌丝干重的影响(3d);
图12为风速对黑曲霉菌丝生长的影响(3d)(a)ck;(b)1m/s;(c)1.5m/s;(d)2.0m/s;(e)3.0m/s;
图13为tio2光催化机理及流相防腐示意图;
图14为不同处理对西梅果实贮藏呼吸速率和乙烯生成量的影响(a)呼吸速率;(b)乙烯产量;
图15为不同处理对西梅果实贮藏ppo活性、总酚类物质含量、腐烂率、mda含量的影响(a)ppo活性;(b)总酚含量;(c)mda含量;(d)腐烂率。
具体实施方式
下面通过具体实施例对本发明作进一步详述,以下实施例只是描述性的,不是限定性的,不能以此限定本发明的保护范围。
一种西梅流相防腐保鲜方法,包括如下步骤:
1、采前品质调控
(1)使用方法。在西梅采收前15天内在叶面和果实表面喷施品质诱导剂,诱导剂由品质提升剂a和品质诱导剂b,各组分协同作用,提高植物吸收效果,促进果实成熟、提高色泽、糖度、脆度等品质。a组份先喷施,隔天喷施b组份,均稀释300倍,各喷施1~3次。
(2)配方。包括组份a、b,其中a组份包括l-arg(l-精氨酸)0.1~0.3g/l、醋酸锌0.2~0.6g/l、sod制剂0.5~1.0g/l、氯化钙1~3g/l、亚油酸10~20mmol/l、mesa(水杨酸甲酯)3.0~8.0mg/l、cys(胱氨酸)3.0~5.0mg/l;b组份包括br(油菜素内酯)0.1~0.5mg/l吲哚乙酸、赤霉酸10~30mg/l、油菜素内酯0.1~1.0mg/l、过氧化氢10~20mg/l、meja(茉莉酮酸甲酯)3~5mg/l、硝普钠5~10mmol/l:
(3)制备工艺
a组份:按预定要求将l-arg(l-精氨酸)0.1~0.3g/l、亚油酸10~20mmol/l、mesa(水杨酸甲酯)3.0~8.0mg/l、cys(胱氨酸)3.0~5.0mg/l溶解在水中,在35~55℃下搅拌15~20min,之后在上述获得的溶液中加入醋酸锌0.2~0.6g/l、氯化钙1~3g/l搅拌均匀,最后在上述溶液中加入sod制剂0.5~1.0g/l、定容至1l的混合溶液;
b组份:
将br(油菜素内酯)0.1~0.5mg/l、吲哚乙酸0.2~0.4mg/l、赤霉酸10~30mg/l、油菜素内酯0.1~1.0mg/l、meja(茉莉酮酸甲酯)3~5mg/l、过氧化氢10~20mg/l用少量乙醇溶解后再加入水中,置于35~55℃下搅拌至均匀溶解形成混合插层剂溶液,之后加入硝普钠5~10mmol/l,经30~50khz超声处理10~20min,定容至1l得到插层混合液b:
2、采收
采前10-15天,严禁灌水,如遇大雨须延迟8-10天采收。长期贮藏果实,采收成熟度7-8成熟为宜,一般着色50%-60%。长期贮藏果实必须手工采摘,轻拿轻放,采后直接放入采果袋,再装入无伤周转箱里。
无伤周转箱,箱体外观为方形,如图1,内部收纳仓2主体结构为圆弧形,减少西梅在运输周转过程中的碰撞与挤压,可注塑一体成型,且中间底部有排水孔1,圆弧形表面为五层复合的保鲜垫,如图2,包括依次设置的阻隔层4、施药层5、载药层6、吸附层7、防雾层8。其中,阻隔层为无纺布。施药层为微囊化植物精油保鲜剂,壁芯比4∶1,芯材为丁香精油:肉桂精油=1:1(体积比),壁材由50wt%β-环糊精,40wt%壳聚糖,10wt%均相乳化剂(聚氧乙烯脱水山梨醇单油酸酯,简称聚山梨酯-80或者吐温80)组成。载药层为聚丙烯膜或聚乙烯膜中的一种。吸附层由5wt%~8wt%的1-甲基环丙烯、94wt%~91wt%的高锰酸钾和1wt%的聚丙烯酸酯组成,防雾层为淋膜纸。
周转箱的箱壁为夹层结构,且侧壁设计有可开启式通风孔3,夹层内可根据实际需要放置蓄冷剂(此时箱壁通风孔关闭),也可用于果蔬通风(不放置蓄冷剂,此时箱壁通风孔打开)。
3、简易隧道高湿差压预冷
简易隧道差压高湿预冷系统,包括塑料帐、静压箱、轴流式排风机、熏蒸加湿装置构成。系统主体为一静压箱9,静压箱顶部安装两个轴流风机10,静压箱正面开长方形孔11,静压箱内安装有加湿器,可定时定量控制加湿过程中的环境湿度,保证预冷过程库内湿度保持在90-95%。静压箱长、宽、高分别为3500mm、400mm、2100mm,静压箱正面中间位置开600mm×1600mm的长方形进气孔,孔底边距地面100mm,静压箱顶部装两个风量为11000m3/h的轴流风机,静压箱板缝之间做气密处理,静压箱结构图如图3。
使用方式:果蔬裸果装筐13码放在托盘14上,托盘放在静压箱开孔一侧,紧贴静压箱,依次对齐排放,用pvc布12把货堆上面和侧面遮严。风机转动时,在静压箱内产生负压,抽吸冷库中的冷空气,冷空气从货堆尾部穿流进入静压箱,如此,达到对果蔬预冷的目的。预冷过程中同时使用1μl/l1-mcp处理,拮抗乙烯,处理时间12h。果实放入筐内,单个筐的尺寸为:长×宽×高=500mm×500mm×170mm,每筐包装量约为15kg,托盘尺寸为:长×宽=1500mm×1000mm,托盘每层放6筐西梅,共放11层,如此,单个托盘货物尺寸为:长×宽×高=1500mm×1000mm×2100mm,货物重量约1000kg。差压预冷摆货方式:6米纵向无间隙摆放:双垛堆、纵向、无间隙、平行排列,共8个托盘。即长龙垛堆宽度2000mm、高度2100mm,两垛堆间距为零(靠紧),龙长度6000mm,可摆放8t货物,如图4。
在货物摆放之前,将无线测温系统温度传感器插入果心,以监测果实温度。温度探头共19个,其中一个放在冷库内,监测冷库温度,其余分别放入货堆不同部位:将货堆长度方向平均分为前(靠近静压箱)、中、后(长龙尾部);高度方向平均分为:上层、中层、下层;宽度方向分为内侧和外侧。在前中后、上中下和内外交叉的各点布置温度探头,共18个探头,监测果心温度变化。设置冷库风机停机温度-2℃,回温差1℃。以普通静止预冷为对照,将8个整托盘果实散放在普通冷库中,托盘之间间隔1米,以便于果实接触更多的冷风。从图5中可以看出,平均5-6h降至0℃。
4、相温库贮藏
相温贮藏,精准控温±0.1℃,不加湿恒湿。相温库结构主要包括母库17、相温库地面20、子库16、子库内循环通风系统19。
(1)相温库为子母夹套结构,母库库体保温材料为150mm厚高密度聚氨酯双面彩钢板(容重40kg/m3),母库气密保温,配有标准气调门,如图6。
(2)子库板使用1mm厚铝板,子库气密不保温,子母库间距500mm,顶间距1000mm,子库临门两侧各安装1个子母库通风与作业窗18,联合进行子母库大幅度热量交换,也可用作维修作业窗,流相防腐系统安装在子库上,如图7、图8。
分别测定了传统冷库和相温库的温度波动,温度波动的随机变化会导致冷冻农产品的中心温度暂时降低到阈值水平以下,超过这个阈值,冷损伤就可能发生。此外,温度波动可能加剧水分凝结,导致微生物生长和果实腐烂。分别测量了冷库和相温库中0.5和3.5m高平面的温度分布。在温度设置为0℃时,与冷库图9中(c,d)的±0.5~±1.0℃相比,相温库的温度波动很小图9中(a,b),只有±0.1~±0.2℃。冷库由制冷设备控制温度,采用直接冷却的方式,蒸发器15吹出的冷风直接与库内产品接触。而相温库的子库的冷量是通过空气流经内库的墙壁、地板和天花板周围的封闭空间或夹套进行冷却的,而不是通过房间内空气的直接循环进行冷却的。通过增加内部结构,避免了产品与蒸发器的直接接触,保持了内部房间的低温度波动。
5、贮藏期流向风内循环联合tio2光催化处理抑菌并提高贮藏品质
5.1流相风速内循环具有显著的抑菌作用
西梅贮藏过程中的主要致病菌有黑曲霉、灰霉菌、青霉菌和链格孢等。研究证明流相风速内循环具有显著的抑菌作用。当风速>1.5m/s时,四种真菌菌落生长受到抑制,流相风速处理抑制了四种病原真菌的菌丝体生长,延缓孢子萌发。但是,风速循环对菌种表现出一定的选择性,黑曲霉对风速更加敏感,风速1.5m/s流相处理表现出较好的延缓病原真菌生长的能力。风速流相内循环均抑制了黑曲霉菌落的生长,1.5m/s风速处理破坏了黑曲霉菌丝细胞膜结构,使细胞膜透性增大,胞内物质流出,可溶性蛋白含量提高,同时,风速导致黑曲霉麦角甾醇生物合成的大量减少,使黑曲霉质膜功能受损。sem结果显示,风速通过改变分子孢子的表面速度,导致作用于分子孢子上的阻力变化,导致分生孢子的弯曲,从而降低了孢子的释放通量。1.5m/s和2.0m/s风速处理导致菌丝形态不规则,菌丝细且褶皱严重,扭曲形变,部分断裂导致内容物流失。研究结果如下:
(1)不同种类病原真菌对流相风速响应分析
风速对四种主要果蔬采后病原真菌的菌落生长均具有一定的抑制作用(表1)。与对照组相比,其他风速处理均影响平板中菌落的萌发速度,随着培养时间的增加,平板中的菌落数均呈现递增状态,但当培养至第5天时,不同风速处理组的菌落数目跟对照组相比,都无显著差异(p>0.05),这表明风速处理可以延缓菌落的生长和萌发,但是不能完全抑制菌落的萌发。四种病原真菌中,孢子接种浓度和接种量相同的条件下,各个风速处理的黑曲霉菌的平均菌落数低于其他三种病原真菌(p<0.05)。另外,在培养的前4天,当风速超过1.5m/s时,病原真菌的菌落数显著低于0m/s处理组和1m/s处理组。根据平均菌落数,四种病原真菌对风速的响应依次为黑曲霉>灰霉菌>青霉菌>链格孢。
表1不同风速对病原真菌菌落数的影响
*每一行内不同字母的平均值表示处理之间的显著差异(p<0.05)
(2)流相内循环风速对病原真菌体外生长速率的影响
图10为流相循环培养第3d时,不同真菌的平均菌斑直径,从图中可以看出,随着风速的增大,菌斑直径呈减少趋势。其中,黑曲霉的菌斑直径显著低于其他3种病原真菌(p<0.05)。而链格孢霉菌、灰霉菌和青霉菌在0m/s和1m/s两个风速处理条件下,菌斑直径差异不显著(p>0.05)。链格孢和青霉菌在1.5m/s和3.0m/s两个风速处理条件下,菌斑直径差异显著(p>0.05)。从菌斑直径可以看出,流相内循环风速处理可以显著抑制四种病原真菌的体外生长。而黑曲霉菌的菌斑直径最小,说明风速流相内循环对黑曲霉的生长抑制效果最显著。为了量化风速对病原真菌生长的抑制作用,还测定了不同风速处理条件下培养3d积累的菌丝量,将各个处理组平板的菌丝收集下来并恒温烘干后称重,从图11中可以看出,当风速>1.5m/s时,黑曲霉和灰霉菌的菌丝干重为0m/s处理的1/2,结果表明,风速可以显著抑制黑曲霉和灰霉菌的生长和生物量的积累。
(3)流相内循环风速对病原真菌产孢子量和孢子萌发率的影响
分生孢子的萌发和发展是真菌生活史和循环侵染的重要的组成部分,它与真菌的扩散和传播密切相关。本实验结果显示,不同风速对果蔬病原真菌在pda上培养时的产孢子量有显著的差异(表2),且不同菌种、不同风速条件产孢子量和孢子萌发率不同。流相风速内循环降低了四种病原真菌的产孢子量(表2)。同一菌种,在四种风速处理条件下,当风速为1.5m/s时,其孢子产量和萌发率开始显著下降,以灰霉菌为例,其在时的产孢子量为23.62×107个,在0m/s时产孢子量为98.63×107个,但是1.5m/s、2.0m/s和3.0m/s三个风速处理的产孢子量差异不显著(p>0.05),这说明风速超过1.5m/s时,风速的增加对真菌的产孢子量影响不大。值得注意的是,不同种类病原真菌间对比,在接种量相同的条件下,黑曲霉具有最小的产孢子量,其在生长后第4天,1.5m/s处理组的产孢子量为8.53×107个,链格孢为10.14×107个,灰霉菌为23.62×107个,青霉菌为44.35×107个。
表2不同风速对病原真菌产孢子量的影响
接种后4d测定孢子产孢子量。每行相同字母表示的平均值差异不显著,p<0.05,方差分析采用duncan’s多量程检验。
风速处理也降低了病原真菌的孢子萌发率。与对照组相比,风速处理组均降低链格孢、黑曲霉和灰霉菌的孢子萌发率(表3)。最低的孢子萌发率是1.5m/s处理组,风速对真菌的孢子萌发率的影响取决于风速大小和真菌种类,对于青霉菌,其在0m/s和1m/s条件下,孢子萌发率差异不显著(p>0.05)。1.5m/s风速流相处理下,链格孢、黑曲霉、灰霉菌和青霉菌的孢子萌发率分别为:58.5%、30.5%、51.4%和66.9%。黑曲霉在5种不同风速处理条件下的孢子萌发率分别为:51.4%,31.4%,20.5%,22.0%和19.4%。结果表明,不同风速条件对四种病原真菌的孢子萌发率会有不同影响,这与不同的菌种有关,也与风速大小有关。不论是产孢子量和孢子萌发率,在1.5m/s流相处理下均受到明显抑制。真菌的主要生长发育阶段生命通常包括孢子萌发、菌丝扩张生殖结构的形成,在这些阶段受到各种环境因素的调节通过分析产孢子量、孢子萌发率、菌落扩展率和菌丝积累量,发现流相内循环风速处理明显降低了四种病原真菌的生长速率。风速循环对菌种表现出一定的选择性,与链格孢霉菌和青霉菌相比,黑曲霉和灰霉菌对风速更加敏感,在流相风速试验方面,1.5m/s的风速表现出较好的延缓病原真菌生长的能力。
表3不同风速对病原真菌孢子萌发率的影响
接种后1d测定孢子萌发率。每行相同字母表示的平均值差异不显著,p<0.05,方差分析采用duncan’s多量程检验。
(4)不同风速对黑曲霉菌丝和孢子形态的影响
通过sem电镜观察不同风速条件下孢子和菌丝状态,可以看出,风速处理对菌丝形态产生明显的影响(图12)。对照组菌丝饱满,表明光滑,形状规则,孢子数量多。而1.0m/s风速处理组的菌丝形态已经发生不规则的形态变化,主要表明为卷曲,干瘪,皱缩形变,这表明菌丝内容物流失。对比之下,1.5m/s和2.0m/s风速处理组的菌丝形态更加不规则,菌丝细且褶皱严重,扭曲形变,部分断裂,且孢子数目明显减少,这说明风速处理对黑曲霉的菌丝有明显的损伤作用。许多杀菌剂能够杀灭真菌的重要作用模式之一是改变病原菌的菌丝形态,导致菌丝形变和断裂,菌丝的断裂可能导致菌丝细胞表明的通透性改变,导致内含物流出。据报道,在空气流速过程中,由于空气阻力的作用,分生孢子会向空气中弯曲,从而导致分生孢子数量较低,风速使菌丝发生了摇摆和弯曲,而长时间暴露在风速中,打破了菌丝的自平衡能力,导致真菌菌丝表面弯曲甚至塌陷。
5.2利用流相tio2光催化提高西梅采后品质的影响
子库内循环通风系统,如图13,由上下通风口背向采用铝箔蛇形管或pvc管(ft)由法兰盘连结而成,上孔安装轴流风机,风量是库容积的30-40倍(7000-8000m3/h)。其中下孔为圆形密封窗,直径400mm,向子库内打开。贮藏期流相防腐。其中tio2的粒子比表面积为11.7m2g-1,当量粒径为93.7nm,紫外光波长为365nm,tio2包覆的表面积为592.8cm2。在风速1.5m/s,条件下贮藏期内协同使用流相tio2光催化,每7天处理1次,每次30min,tio2光催化启动时,流相风速内循环同步启动,另外,流相风速内循环每天运行2h。
(1)呼吸速率和乙烯产量
如图14中a所示,ck和lft治疗组在第20d达到第一个呼吸峰,其峰值分别为32.96和30mgco2kg-1h-1的,而lft+o3和lft+tio2处理组在贮藏期内只有一个呼吸风,出现在第50d,分别为24.68和26.52mgco2kg-1h-1。ck和lft处理组在第50d分别达到最大值35.25和32.25mgco2kg-1h-1。另外,lft+o3处理的西梅子在20~50d呼吸频率显著低于其他三组(p<0.05)。臭氧主要通过抑制水果细胞线粒体和正常电子传递呼吸链的氧化磷酸化来抑制呼吸lft+o3和lft+tio2处理组的呼吸轻度均低于对照组和lft组。作为典型的更年期果实,抑制或延缓呼吸峰的出现是保持西梅果实贮藏品质的关键措施。结果表明,tio2光催化反应器或臭氧联合lft使用能显著抑制西梅的呼吸。
乙烯的释放可以加速西梅子在贮藏期间的成熟和衰老。如图14中b所示,lft+o3和lft+tio2组的最大乙烯释放量分别为12.7和13.6μlkg-1h-1,在40d贮藏时显著低于ck和lft组(p<0.05)。ck组与lft+o3组果实乙烯产量差异极显著,ck组果实乙烯产量是lft+o3组果实的两倍以上。此外,我们的研究结果表明,臭氧或tio2光催化分别联合低温度波动处理对西梅贮藏过程中乙烯的降解具有良好的协同效应。在先前的一项研究中也报告了类似的结果,tio2和紫外光的存在可以从储存大气中去除乙烯气体。
(2)ppo活性和总酚含量
ppo一直被认为是导致果实采后变色的主要因素。如图15中a所示,各处理贮藏前20dppo活性无显著差异。但从40d开始,lft处理和ck处理的果实中ppo活性均迅速下降,这可能是由于果实衰老和过熟所致。贮藏20d后,对比对照组,lft+o3andlft+tio2处理组可以有效抑制西梅果实的ppo活性。此外,在60d的存储期内,lft+tio2处理比其他处理可以降低ppo活性。
在所有西梅子处理中,总酚含量在前40d逐渐增加,然后在贮藏期间下降,并且用lft+tio2处理的西梅子具有最低的总酚含量和最低的增长率(图15中b)。这可能与tio2光催化有关。作为酶促褐变的底物,总酚含量在褐变程度上表现出正相关的响应降低总酚含量和ppo活性是果实抗褐变的主要方式之一。在本实验里,lft+o3和lft+tio2处理可以有效地减少总酚的形成,抑制ppo的活性。贮藏40d后,lft+o3组果实中ppo活性和总酚含量较高。这可能与西梅果实过度暴露于臭氧中的生理损伤有关。总之,lft+tio2处理能有效降低西梅子ppo活性和相应的总酚含量。
(3)丙二醛(mda)含量
如图15中c所示,所有处理的丙二醛含量都显著增加。但lft+tio2组西梅果实mda含量明显受到抑制,贮藏结束时,其仅为ck组果实腐烂速率的60.6%。lft处理降低和延缓了mda的积累,在第60d,lft处理果实中的mda含量为2.9mmolkg-1,比对照降低约12%。在较大的温度波动下,细胞膜从凝胶相变为液晶相,这增加了半透膜损失的风险。然而,贮藏40d后,lft+o3处理组果实中mda含量较高,这可能是由于臭氧处理的果实在低温下缺乏清除自由基的能力。此外,膜完整性受损后,酚类化合物与ppo的相互作用增强,导致果实组织退化或衰老。在本研究中,观察到lft+tio2处理降低了mda含量。
(4)腐烂率
如图15中d所示,ck和lft组的果实在前20d内腐烂,并且腐烂程度明显高于lft+o3和lft+tio2组,lft+tio2和lft+o3处理30d后果实出现腐烂症状,西梅果腐烂率明显降低。在60d时,lft+tio2和lft+o3处理的衰减率分别比对照组降低65.2%和75.3%。tio2光催化反应器在特定波长的光照射下产生的超氧阴离子自由基(·o2-)和羟基自由基(·oh)具有很强的氧化分解能力,可以通过破坏细胞膜中的蛋白质来杀灭细菌。
以上所述的仅是本发明的优选实施方式,应当指出,对于本领域的普通技术人员来说,在不脱离发明构思的前提下,还可以做出若干变形和改进,这些都属于本发明的保护范围。
起点商标作为专业知识产权交易平台,可以帮助大家解决很多问题,如果大家想要了解更多知产交易信息请点击 【在线咨询】或添加微信 【19522093243】 与客服一对一沟通,为大家解决相关问题。
与客服一对一沟通,为大家解决相关问题。
此文章来源于网络,如有侵权,请联系删除

 热门咨询
热门咨询



