
 商标分类
商标分类  商标转让
商标转让 
水稻osa-miR5504基因在水稻矮化育种中的应用的制作方法
 2021-02-02 18:02:32|
2021-02-02 18:02:32| 436|
436| 起点商标网
起点商标网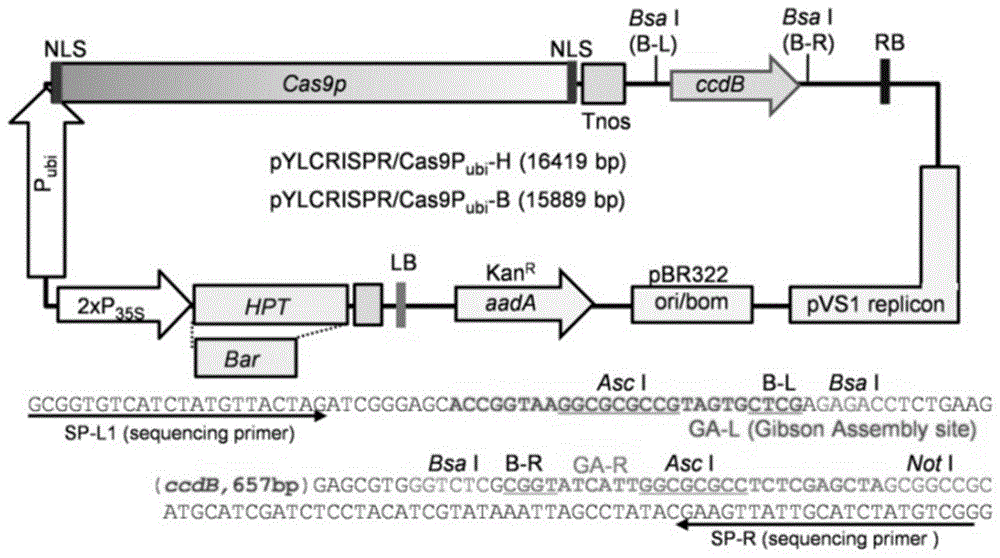
本发明属于植物基因工程技术领域,具体涉及osa-mir5504在水稻矮化育种中的应用,通过抑制osa-mir5504的表达来创制矮秆、抗倒伏水稻新品种的分子育种。
背景技术:
水稻养活了近一半的世界人口。因此,如何保障水稻的产量与品质对于解决世界粮食安全问题至关重要。在水稻生产过程中,由于台风、飓风、肥水管理等多变的人为或自然因素的影响,水稻植株往往会出现不同程度的倒伏现象,导致水稻的产量与品质明显降低,世界粮食安全也就面临着严峻的考验。
通常情况下,高杆水稻品种易于倒伏,而矮秆水稻品种的抗倒伏能力明显增强;因此,利用现代的基因工程手段培育优良的矮化水稻品种,以降低水稻品种的倒伏率、提高或保障水稻品种的产量与品质,是现代水稻分子育种关注的焦点问题之一。
mirna(microrna)是生物体内普遍存在的一类20-24nt的内源性小分子非编码rna。研究表明,mirna主要是通过控制它直接作用的靶基因表达水平来影响植物的生长发育和逆境适应。现在已知,水稻中至少存在着592个mirna前体,能产生至少713个成熟的mirna。目前,人们已经发现了一些与水稻植株高矮等性状调控相关的mirna。例如,mir159可通过影响其靶基因osgamyb和osgamybl1的表达,正向调控水稻的多种株型性状,包括茎节的长短粗细等[zhaoetal.suppressionofmicrorna159impactsmultipleagronomictraitsinrice(oryzasatival.).bmcplantbiology,2017,17:215.];microrna172可通过调控sui4/snb基因的表达来影响水稻的茎节长度等[jietal.mutationsinthemicrorna172bindingsiteofsupernumerarybract(snb)suppressinternodeelongationinrice.rice,2019,12:62.]。本专利首次公开了一种通过抑制osa-mir5504的表达以创制矮化水稻品种的方法,该方法目前尚未见报道,为利用osa-mir5504以培育抗倒伏的优良矮化水稻品种提供了一种新的分子育种途径。
技术实现要素:
本发明的目的是提供osa-mir5504在水稻矮化育种中的应用。运用了基因工程技术以抑制osa-mir5504基因的表达,为利用osa-mir5504基因以培养优良的矮秆水稻品种提供了一种新的分子育种方法。
为实现上述目的,本发明采用的技术方案是:
水稻osa-mir5504基因在水稻矮化育种中的应用,通过基因工程技术对水稻osa-mir5504基因进行敲除或抑制其表达,选育矮杆的水稻品种,所述水稻osa-mir5504基因的成熟序列如seqidno.1所示,其发夹结构序列如seqidno.2所示。
本发明优选crispr基因编辑技术进行敲除或抑制水稻osa-mir5504基因表达,包括以下步骤:
(1)osa-mir5504的crispr/cas9敲除表达载体构建:利用crispr/cas9敲除靶点引物对、水稻内源u6a启动子以及pylcrispr/cas9质粒,经过限制性内切酶酶切、连接、转化、菌落pcr检测、sgrna序列的测序分析,获得osa-mir5504的crispr/cas9敲除表达载体;
(2)osa-mir5504缺失表达矮秆水稻植株的获得:将上述构建好的osa-mir5504的crispr/cas9敲除表达载体,经农杆菌介导法,转化粳稻日本晴,获得转基因水稻植株,在随后的世代分离过程中,可采用pcr、测序分析、qrt-pcr、northern-blot杂交方法以鉴定获得无t-dna插入、无脱靶现象的osa-mir5504缺失表达转基因纯合水稻植株,并将其与野生型水稻同时种植于大田中,于抽穗晚期和成熟期进行株高和有效分蘖数的比较分析,以确定获得了osa-mir5504缺失表达的矮秆水稻植株。
与现有技术相比,本发明的有益效果是:
本发明提供了水稻osa-mir5504基因在水稻矮化育种的应用,通过抑制osa-mir5504的表达来降低水稻植株的高度,以防止水稻倒伏,达到高产或稳产的目的,为利用osa-mir5504基因培养优良的矮秆水稻品种提供了一种新的分子育种途径,其应用前景广阔。本发明可显著降低水稻的株高,同时还能显著增加水稻的单株有效分蘖数。
附图说明
图1为pylcrispr/cas9质粒结构示意图。其中,p35s为花椰菜花叶病毒的35s启动子;pubi为玉米泛素蛋白基因启动子;ccdb为ccdb致死基因,经bsai酶切后被sgrna表达盒替换,以完成敲除载体构建;tnos为终止子;rb为右边界;lb为左边界;kanr为卡那霉素抗性基因;hpt为潮霉素抗性基因;pbr322ori为pbr322的复制起始位点;cas9p为cas9蛋白基因;sp-l1和sp-r为菌落pcr引物对和测序引物。
图2为osa-mir5504的crispr/cas9敲除靶位点和脱靶位点的测序鉴定。a:osa-mir5504的茎环结构序列、成熟体序列(红色字的序列)和敲除靶点序列(黄色底的序列);b:靶点突变类型的测序分析;c:可能的脱靶靶点序列及其在基因组上的位置;d、e和f:在nip、cr1和cr2突变体基因组中os09g0307050、os11g0171500和os12g0169950这3个区段里最可能脱靶靶点序列未发生突变的测序结果。nip是野生型水稻日本晴,cr1是在osa-mir5504成熟体序列中发生了1个c碱基缺失的突变体,cr2是在osa-mir5504成熟体序列中发生了12个碱基缺失的突变体。
图3为crispr/cas9转基因纯合突变体中成熟osa-mir5504的表达检测。a:stemloopqrt-pcr鉴定结果。b:stemloop半定量rt-pcr鉴定结果。c:northern-blot杂交检测结果。m:markerdna;nip:日本晴(野生型);cr1和cr2:分别为2个mir5504的crispr/cas9敲除纯合突变体;u6为内参基因。
图4为osa-mir5504缺失表达水稻的株高和单株有效穗数分析。a:抽穗晚期osa-mir5504缺失表达突变体的株高。b:成熟期osa-mir5504的株高测量统计结果。c:成熟期osa-mir5504突变体每个单株所有有效分蘖的各茎节长度测量统计结果。nip:日本晴(野生型);cr1和cr2:分别为osa-mir5504缺失表达突变体。i、ii、iii、iv和v:分别为紧挨着穗子的第1、2、3、4、5茎节。**:p<0.01的极显著差异。
具体实施方式
以下实施例中所涉及的实验方法如无特别说明均为常规的实验方法。本发明中,引物序列合成和dna序列测序分析均由上海生物工程技术有限公司完成。
实施例1:osa-mir5504的crispr/cas9敲除载体构建及水稻遗传转化
在mirbase数据库(http://www.mirbase.org/)中获得水稻osa-mir5504的成熟序列seqidno.1及其发夹结构序列seqidno.2,进而利用在线软件crispr-p2.0(http://crispr.hzau.edu.cn/crispr2/)设计获得osa-mir5504的crispr/cas9敲除靶点引物对seqidno.3:gccgacagtgacgggaggactgca(下划线为bsai酶切位点)和seqidno.4:aaactgcagtcctcccgtcactgt(下划线为bsai酶切位点);随后,经变性退火与水稻u6启动子连接成sgrna表达盒,再经酶切、连接到pylcrispr/cas9质粒中,获得mir5504的crispr/cas9敲除载体,进而参考已有的方法[hieietal,efficienttransformationofrice(oryzasatival.)mediatedbyagrobacteriumandsequenceanalysisoftheboundariesofthet-dna.plantj,1994,6(2):271-282],利用农杆菌介导法转化粳稻日本晴,大致过程可为诱导、继代培养、预培养、共培养、抗性筛选、分化、生根培养、炼苗和移栽等,共获得33株遗传转化t0代水稻植株,利用ctab法提取其基因组dna后,进行潮霉素基因特异引物的pcr鉴定,确定获得的转基因水稻植株全为阳性,潮霉素基因扩增的特异引物对序列如下:
hpt-f:5’-ctgaactcaccgcgacgtctgtc-3'(seqidno.5);
hpt-r:5’-tagcgcgtctgctgctccataca-3’(seqidno.6);
pcr扩增条件为:94℃4min;[94℃30sec,62℃30sec,72℃50sec],32个循环;72℃7min。
实施例2:osa-mir5504的crispr/cas9敲除缺失表达突变体的分子鉴定
根据osa-mir5504的crispr/cas9敲除靶点序列信息(图2a),在其对应的水稻基因组上设计一对特异引物(seqidno.7和seqidno.8),以野生型(日本晴)和上述提取的33份水稻基因组dna为模板,扩增获得包含该敲除靶点序列的单一目的基因组dna片段,进行测序分析,确定获得发生了敲除突变的t0代转基因植株,并在其分离后代苗期进一步进行潮霉素基因阳性检测和靶位点的测序分析,获得了不含t-dna插入的osa-mir5504敲除突变体。其中有两个在成熟osa-mir5504序列中发生突变、无外源t-dna插入、无脱靶现象的纯合敲除突变体(cr1和cr2)(图2b-f)可用于后续实验,经rt-pcr、qrt-pcr和northern-blot杂交实验证实,成熟osa-mir5504在cr1和cr2突变体中均是缺失表达的(图3)。northern-blot杂交探针序列为seqidno.9(cttgcagtcctcccgtcact),其中5’端的核苷酸加dig修饰,3’端的3个核苷酸进行lna锁定。
实施例3:osa-mir5504缺失表达显著降低水稻株高和增加单株有效穗数
随后,将野生型日本晴(nip)与osa-mir5504缺失表达突变体(cr1和cr2)同时种植于大田中,于抽穗晚期发现:osa-mir5504缺失表达突变体cr1和cr2的株高明显比野生型nip的要矮(图4a)。于是,在成熟期对nip、cr1和cr2的株高和有效分蘖数进行了比较分析,发现:与nip相比,cr1和cr2的株高显著降低(图4b),cr1和cr2的株高显著降低与其紧挨着穗子的第i和第ii茎节明显变短密切相关(图4c);同时,cr1和cr2的单株有效分蘖数却显著增多(图4d)。
序列表
<110>南昌大学
<120>水稻osa-mir5504基因在水稻矮化育种中的应用
<160>9
<170>siposequencelisting1.0
<210>1
<211>21
<212>rna
<213>水稻(oryzasativa)
<400>1
agugacgggaggacugcaagg21
<210>2
<211>98
<212>rna
<213>水稻(oryzasativa)
<400>2
auugcugaauuuuguucguuggagcuuguucuguuauuuuauuugcacugccguaggaaa60
uacacagugacgggaggacugcaagguucuucagcuuu98
<210>3
<211>24
<212>dna
<213>人工序列(artificialsequence)
<400>3
gccgacagtgacgggaggactgca24
<210>4
<211>24
<212>dna
<213>人工序列(artificialsequence)
<400>4
aaactgcagtcctcccgtcactgt24
<210>5
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>5
ctgaactcaccgcgacgtctgtc23
<210>6
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>6
tagcgcgtctgctgctccataca23
<210>7
<211>28
<212>dna
<213>人工序列(artificialsequence)
<400>7
aatgctgctcaaaatgtaaaagagacca28
<210>8
<211>25
<212>dna
<213>人工序列(artificialsequence)
<400>8
gatgttttagatggggccttttggt25
<210>9
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>9
cttgcagtcctcccgtcact20
起点商标作为专业知识产权交易平台,可以帮助大家解决很多问题,如果大家想要了解更多知产交易信息请点击 【在线咨询】或添加微信 【19522093243】 与客服一对一沟通,为大家解决相关问题。
与客服一对一沟通,为大家解决相关问题。
此文章来源于网络,如有侵权,请联系删除

 热门咨询
热门咨询



