
 商标分类
商标分类  商标转让
商标转让 
用包含NC-1-Fc的蛋白寡聚体治疗血管发生、纤维化和癌症相关疾病的手段和方法与流程
 2021-02-02 17:02:16|
2021-02-02 17:02:16| 268|
268| 起点商标网
起点商标网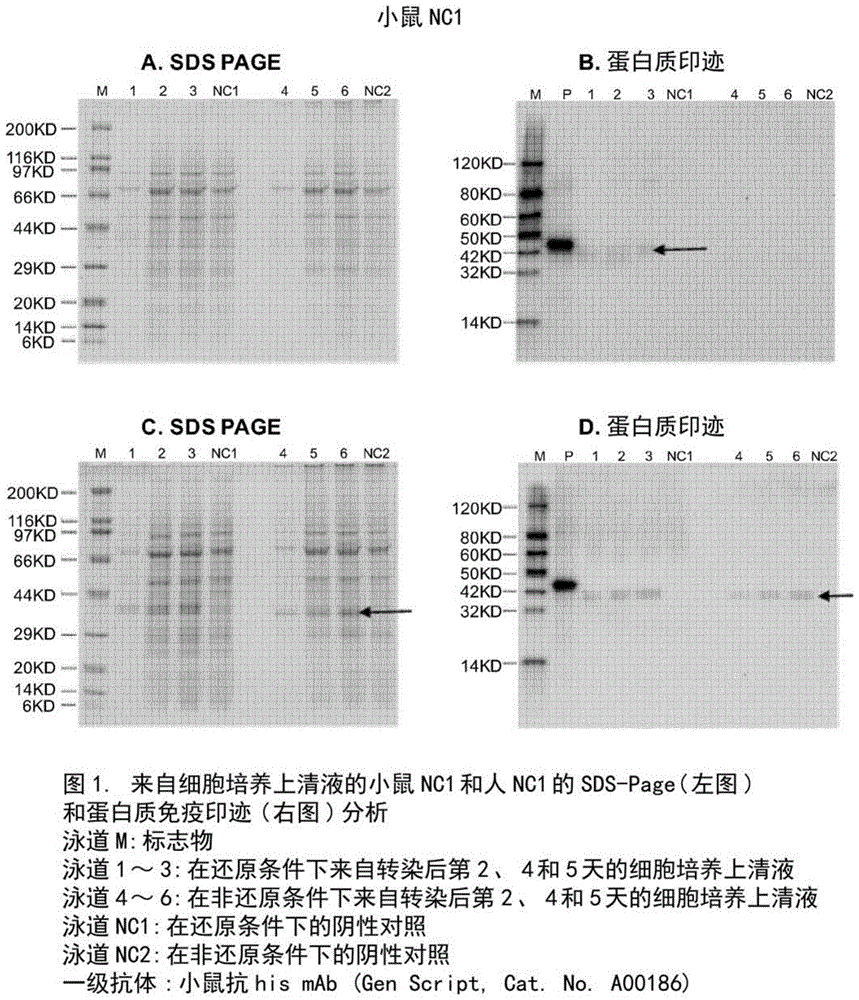
本发明涉及生产包含至少两个,且优选三个异源二聚体人nc-1-fc蛋白的蛋白寡聚体的方法,所述方法包括:a)在允许形成包含至少两个,且优选三个异源二聚体人nc-1-fc蛋白的蛋白寡聚体的条件下培养表达(i)和(ii)的宿主细胞,所述(i)为融合蛋白,其从n-至c-端包含,融合至具有“杵”突变的人igg1fc的来自胶原蛋白18的人nc-1,或融合至来自胶原蛋白18的人nc-1的具有“杵”突变的人igg1fc,所述(ii)为具有“臼”突变的人igg1fc,其中(i)所述的融合蛋白和(ii)所述的具有“臼”突变的人igg1fc以2∶1或更高的比例表达,以及b)从步骤a)的所述宿主细胞中获得包含至少两个,且优选三个异源二聚体人nc-1-fc蛋白的蛋白寡聚体。本发明进一步涉及生产包含至少两个,且优选三个单体人nc-1-fc蛋白的蛋白寡聚体的方法,所述方法包括:a)在允许形成包含至少两个,且优选三个单体人nc-1-fc蛋白的蛋白寡聚体的条件下培养表达融合蛋白的宿主细胞,所述融合蛋白从n-至c-端包含:融合至人igg1fc的来自胶原蛋白18的人nc-1,或融合至来自胶原蛋白18的人nc-1的人igg1fc,其中所述人igg1fc包含至少一个单体突变,优选地单体突变f405r,和一个或更多个半衰期延长突变,优选半衰期延长突变m252y、s254t和t256e,以及b)从步骤a)的所述宿主细胞中获得包含至少两个,且优选三个单体人nc-1-fc蛋白的蛋白寡聚体。进一步地,本发明提供蛋白寡聚体,其包含至少两个,且优选三个异源二聚体人nc-1-fc蛋白,所述蛋白寡聚体用于治疗、改善或预防与血管发生有关的疾病,其中所述异源二聚体人nc-1-fc蛋白包含(i)融合蛋白,其从n-至c-端包含:融合至具有“杵”突变的人igg1fc的来自胶原蛋白18的人nc-1,或融合至来自胶原蛋白18的人nc-1的人igg1fc,和(ii)具有“臼”突变的人igg1fc,其中人igg1fc中的所述“杵”突变和“臼”突变是如本申请所限定的。本发明还涉及单体nc-1-fc蛋白,其包含来自胶原蛋白18的人nc-1和来自人igg1的fc结构域,其中来自人igg1的fc结构域包含至少一个单体突变,优选地单体突变f405r,和一个或更多个半衰期延长突变,优选半衰期延长突变m252y、s254t和t256e。本发明还涉及包含至少两个所述单体人nc-1-fc融合蛋白的蛋白寡聚体,其用作药物,优选地用于治疗与血管发生有关的疾病、纤维化或与纤维化相关的疾病、血管内皮生长因子(vegf)相关疾病或基质金属蛋白酶(mmp)相关疾病。
由folkman实验室(oreilly等,cell88(2):277-85,1997)1997年生产的原始重组内皮抑制素是在大肠杆菌中生成的不溶性聚集体,其被皮下注射至小鼠。随后,可溶性内皮抑制素在酵母中产生。entremed单体内皮抑制素在关键的n-端zn2+结合结构域中缺乏很大一部分。此种单体内皮抑制素在一期试验中显示出中等程度的反应,且生产成本很高,以至于仅进行了少量试验(有关综述,请参见abdollahidru2015pmid:15939343)。
中国科学家通过向n-端添加his标签来开发恩度(endostar),导致更好的增溶作用,并发现了一种重折叠程序以在细菌培养物中产生内皮抑制素单体,该内皮抑制素单体在非小细胞肺癌中有效(us7078485b2)并获得了中国fda的批准。
因此,即使内皮抑制素的生产也是一个挑战。javaherian博士与lexigen(后来的merckkga)一起开发了fc-内皮抑制素,最初是为了规避两个障碍,即使用常规的成熟的抗体生产平台来提高大量内皮抑制素的表达以用于临床试验水平生产。此外,后来的研究揭示了更好的半衰期(lee等ccr14(5):1487-93,2008),其是fc-内皮抑制素针对重组内皮抑制素分子的仅30分钟至1小时的半衰期的另一个论点。
包括内皮抑制素结构域的胶原蛋白18nc-1的生产从一开始就是一个巨大的挑战,这是由于该分子在寡聚化结构域上的聚集。因此,没有关于重组nc-1功效的体内数据,并且对迄今为止被认为是内皮抑制素的前体分子仅进行了很少的研究。
javaherian博士和其他少数实验室能通过his标签或flag标签表达很少的重组nc-1,但对于动物的临床前给药而言,这还远远不够。因此,与体内研究相关的nc-1量的产生仍然构成主要挑战。
鉴于上述内容,本领域需要开发用于生产足够用于临床前和临床研究的量的nc-1的有效方法。
可以将本发明所基于的技术问题视为提供符合上述需要的手段和方法。该技术问题已通过权利要求和下文中表征的实施方案解决。
因此,本发明涉及生产包含至少两个,且优选三个异源二聚体人nc-1-fc蛋白的蛋白寡聚体的方法,所述方法包括:
a)在允许形成包含至少两个,且优选三个异源二聚体人nc-1-fc蛋白的蛋白寡聚体的条件下培养表达(i)和(ii)的宿主细胞,所述(i)为融合蛋白,其从n-至c-端包含:融合至具有“杵”突变的人igg1fc的来自胶原蛋白18的人nc-1,或融合至来自胶原蛋白18的人nc-1的具有“杵”突变的人igg1fc,所述(ii)为具有“臼”突变的人igg1fc,其中(i)所述的融合蛋白和(ii)所述的具有“臼”突变的人igg1fc(ii)以2∶1或更高的比例表达,以及
b)从步骤a)的所述宿主细胞中获得包含至少两个,且优选三个异源二聚体人nc-1-fc蛋白的蛋白寡聚体。
在本发明的该方法中,产生蛋白寡聚体。蛋白寡聚体包含至少两个异源二聚体人nc-1-fc蛋白,但也能够包含多于两个异源二聚体人nc-1-fc蛋白,例如三个、四个、五个、六个或甚至更多个异源二聚体人nc-1-fc蛋白质。优选地,蛋白寡聚体包含三个异源二聚体人nc-1-fc蛋白质。
图9说明了异源二聚体人nc-1-fc蛋白和包含三个异源二聚体人nc-1-fc蛋白的蛋白寡聚体。
异源二聚体人nc-1-fc蛋白包含(i)融合蛋白,所述融合蛋白从n-端到c端包含:融合至具有“杵”突变的人igg1fc的来自胶原蛋白18的人nc-1,或融合蛋白,所述融合蛋白从n-端到c端包含:融合至来自胶原蛋白18的人nc-1的具有“杵”突变的人igg1fc。fc“杵臼”(kih)策略是本领域众所周知的,并在本文其他地方进行了说明。如果来自胶原蛋白18的人nc-1位于融合蛋白的n端,且带有“杵”突变的人igg1fc位于融合蛋白的c端,那么所述融合蛋白(i)在本文中也称为“nc-1-fc-杵”。因此,“fc-杵-nc-1”对应一种融合蛋白,具有“杵”突变的人igg1fc位于所述融合蛋白的n端,而来自胶原蛋白18的人nc-1位于融合蛋白的c端。
异源二聚体人nc-1-fc蛋白进一步包含(ii)具有“臼”突变的来自igg1的fc结构域。所述fc结构域(ii)在本文中也称为“fc-臼”。
包含(i)nc-1-fc-杵或fc-杵-nc-1和(ii)fc-臼的异源二聚体人nc-1-fc蛋白在本文中也称为“nc-1-fc-kih”或“nc-1-fc(kih)”。
nc-1和igg1fc是人类或源自人类的。术语“源自”在本文其他地方解释。igg1的fc结构域中的“杵”和“臼”突变允许形成fc二聚体。
(i)其从n-至c-端包含:融合至具有“杵”突变的人igg1fc的来自胶原蛋白18的人nc-1,或融合至来自胶原蛋白18的人nc-1的具有“杵”突变的人igg1fc,和(ii)具有“臼”突变的人igg1fc的融合蛋白在允许形成蛋白寡聚体的条件下在宿主细胞中表达,其中所述蛋白寡聚体包含至少两个或更多个异源二聚体人nc-1-fc蛋白。为此,宿主细胞可以例如用两个不同的表达载体共转染,一个表达载体包含在合适的启动子控制下的编码融合蛋白的核酸序列,所述融合蛋白从n-至c-端包含融合至具有“杵”突变的人igg1fc的来自胶原蛋白18的人nc-1,或融合至来自胶原蛋白18的人nc-1的具有“杵”突变的人igg1fc,且另一个表达载体包含在合适的启动子的控制下具有“臼”突变的人igg1fc。但是,对于本领域技术人员而言明显的是,宿主细胞也可以用单个的表达载体转染,该载体包含编码融合蛋白(i)和(ii)的在合适的启动子的控制下的核酸序列,所述融合蛋白(i)从n-至c-端包含融合至具有“杵”突变的人igg1fc的来自胶原蛋白18的人nc-1,或融合至来自胶原蛋白18的人nc-1的具有“杵”突变的人igg1fc,所述(ii)为具有“臼”突变的人igg1fc。
本发明人发现,为了防止两个具有“臼”突变的人igg1fc结构域的二聚化(“臼-臼fc二聚体”)和实现异源二聚体人nc-1-fc蛋白的高效生产(所述异源二聚体人nc-1-fc蛋白之后形成具有所需生物学活性的蛋白寡聚体,如本文别处解释和下面实施例中所示),融合蛋白(i)和(ii)的具有“臼”突变的人igg1fc以2:1或更高的比例,诸如3:1或4:1或甚至更高比例表达是重要的,所述融合蛋白(i)从n-至c-端包含,融合至具有“杵”突变的人igg1fc的来自胶原蛋白18的人nc-1,或融合至来自胶原蛋白18的人nc-1的具有“杵”突变的人igg1fc。换句话说:从n-至c-端包含融合至具有“杵”突变的人igg1fc的来自胶原蛋白18的人nc-1,或融合至来自胶原蛋白18的人nc-1的具有“杵”突变的人igg1fc的融合蛋白(i)与具有“杵”突变的人igg1fc(ii)相比,必须在宿主细胞中过量表达。从n-至c-端包含融合至具有“杵”突变的人igg1fc的人nc-1,或融合至来自胶原蛋白18的人nc-1的具有“杵”突变的人igg1fc的融合蛋白(i)与具有“杵”突变的人igg1fc(ii)的2:1或更高的理想比例可以例如通过使用以不同启动子强度指导表达的合适的启动子来实现。
为了达到期望的比例,本发明人利用双倍量的“杵”质粒用于瞬时表达系统。他们还通过使用两个不同的病毒颗粒(每个都携带一个构建体),利用高达4:1的“杵”:“臼”的感染复数(moi)。在一种备选的方法中,还可以在双启动子系统中包括nc-1-fc-杵或fc-杵-nc-1构建体和fc-臼,用于瞬时转染和稳定的细胞生成,如上所述。然后,不是dna(瞬时)或moi(病毒)的量需要超过2:1以利于nc-1-fc-杵或fc-杵-nc-1构建体,而是这可以通过不同的启动子强度,例如cmv“杵”对ef1“臼”等而控制。
优选地,从n-至c-端包含融合至具有“杵”突变的人igg1fc的来自胶原蛋白18的人nc-1,或融合至来自胶原蛋白18的人nc-1的具有“杵”突变的人igg1fc的融合蛋白(i)和(ii)具有“杵”突变的人igg1fc以高于2:1的比例表达。因为“杵”构建体中nc-1寡聚化结构域的聚集,比例等于或高于5:1可能会适得其反。因此,优选的比例是2∶1、3∶1或4∶1。
最后,通过本领域公知的方法从宿主细胞获得包含至少两个或更多个异源二聚体人nc-1-fc蛋白的蛋白寡聚体;参见例如,sambrook等,molecularcloning:alaboratorymanual/sambrook,joseph;russell,davidw.--.3rded.--newyork:coldspringharborlaboratory,2001.ausubel等,currentprotocolsinmolecularbiology。为了从宿主细胞或上清液获得包含至少两个异源二聚体人nc-1-fc蛋白的蛋白寡聚体,本发明人进行了蛋白a纯化和尺寸排阻色谱,指示以1∶1的比例存在两个峰。第一个峰是预想的异源二聚体构建体,第二个峰由“臼-臼二聚体”组成。因此,发明人将“杵”与“臼”移动至2:1以增加nc-1-fc-杵的产生,这几乎减少了第二峰(fc-臼-二聚体)的形成。无论如何,异源二聚体nc-1-fc-kih可以被部分收集器中的第一个峰很好地分离。
在本发明的用于生产包含至少两个异源二聚体人nc-1-fc蛋白的蛋白寡聚体的方法的优选实施方案中,所述方法进一步包括步骤c),其中通过交联非变性异源二聚体人nc-1-fc蛋白来测试(优选的)三聚体的形成。本领域中充分描述了如何进行非变性蛋白质的交联实验。本发明人已经进行了交联以确保天然蛋白质结构(尤其是三级和四级)在电泳时不会丢失。因此,进行交联以鉴定该分子的体内组成,发现该分子以与自然或天然nc-1结构域相同的方式形成三聚体。所述方法优选地用于治疗物质的生产过程中,以确保在未来生产流水线的生产过程中确实形成了nc-1-三聚体。
在本发明的用于生产包含至少两个异源二聚体人nc-1-fc蛋白的蛋白寡聚体的方法的另一个优选实施方案中,人igg1fc包含“杵”突变s354c/t366w,且人igg1fc包含“臼”突变y349c/t366s/l368a/y407v。
优选地,所述具有“杵”突变s354c/t366w的人igg1fc包含seqidno:25,并且具有“臼”突变y349c/t366s/l368a/y407v的人igg1fc包含seqidno:26。
更优选地,从n-至c-端包含融合至具有“杵”突变的人igg1fc的来自胶原蛋白18的人nc-1的融合蛋白(i)包含seqidno:27,29,或31或由其组成,且(ii)的具有“臼”突变的人igg1fc包含seqidno:28,30,或32或由其组成。
特别优选的是,从n-至c-端包含融合至具有“杵”突变的人igg1fc的来自胶原蛋白18的人nc-1的融合蛋白(i)包含seqidno:31或由其组成,且(ii)的具有“臼”突变的人igg1fc包含seqidno:32或由其组成。(参见图9和表1,分子#1)。
在本发明的又一个用于产生包含至少两个异源二聚体人nc-1-fc蛋白的蛋白寡聚体的本发明方法的实施方案中,使用kih改造的人igg4fc代替kih改造的人igg1fc。igg4的fc结构域中的“杵”和“臼”突变允许形成fc二聚体。例如在us2013/0177555a1的图6中,显示了人igg4fc的相应氨基酸序列。
作为抗血管发生和抗癌药,从内皮抑制素转移到nc-1是本申请的发明人第一个主要步骤;参见wo2013/026913和wo2017/093569。
自从这一发现以来,这些发明人迄今已经花费了近十年的时间来实现用于大规模生产nc-1的系统,并且许多构建体和经典标签都失败了,如本文和以下实例所证实的。
发明人的主要目标是在免疫球蛋白iggfc缀合领域中前进,取得众多生物技术、药代动力学和生物学优势。
然而,nc-1-fc的产生不是直接了当的过程,并且对于本领域技术人员根本不是显而易见追求的事情。
其原因是存在两种寡聚力,第一种是由nc-1寡聚(或缔合)结构域诱导的三聚化和聚集力,第二种是由igg-fc诱导的二聚化力。发明人已经花费了大量的时间用所有不同的现有技术,即,nc-1与标准iggfc融合物的c-或n-端的缀合,如先前用内皮抑制素所进行的,而所有这些方法均失败了,如以下实施例中所解释的。
不得不考虑的是,在经典的fc标签方法中,将产生n-端或c-端缀合的nc-1分子,如果没有nc-1寡聚域,该分子将形成二聚体。但是,具有nc-1寡聚域,人们则收到聚集体和非常纯净的表达。
就发明人所知,没有关于成功和有效生产nc-1-fc融合蛋白和含有该融合蛋白的寡聚体的文献。
如本文所述,本发明人已经研究了规避该主要问题的不同方法,并且经过十多年的研究能够提出成功的策略。一个核心点是生成二聚fc单价nc-1或异源二聚nc-1-fc。发明人决定使用在fc中的突变,该突变使用fc“杵臼结构”(kih)策略阻止nc-1-fc与nc-1-fc的二聚化。因此,nc-1-fc杵只会与空的fc-臼二聚化。尽管禁止杵-杵二聚化,但在较低的水平上仍可能发生臼-臼二聚化。实际上,发明人发现了两个峰,一个峰是预期的nc-1-fc-kih异源二聚体,第二个峰是fc-臼-臼二聚体(参见下面的实施例)。这表明nc-1-fc-杵对于细胞来说更难以表达。发明人通过增加nc-1-fc-杵相对fc-臼(例如2:1、3:1、4:1或更高)的表达再次规避了该问题,这主要导致nc-1-fc-kih异源二聚体的一个峰,具有出色的表达功效。然而,在尺寸排阻柱(sec)中,发明人认识到在生理条件下的分子比根据nc-1-fc-kih异源二聚体所期望的分子大得多(参见以下实施例)。随后的交联实验证实,本发明人已经惊人地实现了生产三聚体分子,即nc-1-fc-kih异源二聚体的三聚体。该分子保留了fc加三聚体nc-1的所有优异特性。令人惊讶的是,这种三聚体nc-1-fc-kih分子在结合独特的寡聚nc-1结合伴侣(如纤连蛋白,vegf和mmp-2/9)方面显示出相同的功效。凭借足够数量的重组nc-1的优异的表达率和可用性,本发明人在很长一段时间后现在已经可以使用该nc-1分子在不同疾病模型中进行体内实验。除肺癌(llc)模型和肺纤维化模型中的数据外,正在进行进一步的实验,这将极大地提高发明人对该分子的理解。
值得注意的是,nc-1-fc-杵和nc-1-fc-臼的分别表达以及随后通过氧化还原系统的二聚化均未能产生有效的nc-1-fc构建体。因此,异源二聚体fc的空间位阻对于聚集的成功和缺乏至关重要。接下来,发明人旨在评价向“空”fc-臼中添加部分是否会影响该分子的表达。为此,他们将抗体的经典fab片段缀合到fc-臼,并与nc-1-fc-杵共表达。有趣的是,与nc-1-fc-kih异源二聚体相比,通过fab部分nc-1-fc-kih-fab添加空间位阻降低表达效率,如实施例和表6所示。
同时,发明人报道的三聚体nc-1-fc-kih异源二聚体是第一个基于nc-1的fc构建体,其可以以足够的量表达用于临床前和临床研究,并且鉴于nc-1和fc两者的3d复杂性,改造这样的分子过去不是,而且现在也不是一个可以从当前文献中推论出来的微不足道的过程。
本发明进一步涉及生产包含至少两个,且优选三个单体人nc-1-fc蛋白的蛋白寡聚体的方法,所述方法包括:
a)在允许形成包含至少两个,且优选三个单体人nc-1-fc蛋白的蛋白寡聚体的条件下培养表达融合蛋白的宿主细胞,所述融合蛋白从n-至c-端包含,融合至人igg1fc的来自胶原蛋白18的人nc-1,或融合至来自胶原蛋白18的人nc-1的人igg1fc,其中所述人igg1fc包含至少一个单体突变,优选地单体突变f405r,和一个或更多个半衰期延长突变,优选半衰期延长突变m252y、s254t和t256e,以及
b)从步骤a)的所述宿主细胞中获得包含至少两个,且优选三个单体人nc-1-fc蛋白的蛋白寡聚体。
同样在本发明的该方法中,产生蛋白寡聚体。蛋白寡聚体包含至少两个单体人nc-1-fc蛋白,但也能够包含多于两个单体人nc-1-fc蛋白,例如三个、四个、五个、六个或甚至更多个单体人nc-1-fc蛋白质。优选地,所述蛋白寡聚体包含三个单体人nc-1-fc蛋白质。
图10显示了一个单体人nc-1-fc蛋白和一个包含三个单体人nc-1-fc蛋白的蛋白寡聚体。
单体人nc-1-fc蛋白包含融合蛋白,所述融合蛋白从n-端到c-端包含,融合至人igg1fc的来自胶原蛋白18的人nc-1,或融合至来自胶原蛋白18的人nc-1的人igg1fc。单体人nc-1-fc蛋白在本文中也称为“nc-1-fc(单体)”。
人igg1fc包含一个或更多个单体突变,这意味着人igg1fc结构域被改造成单体fc,在其中位于人igg1fc二聚体界面上的位置的一个、两个、三个、四个、五个或甚至更多的关键氨基酸残基被突变了。带有一个或更多个这样的单体突变的人igg1fc不再能够与另一个人igg1fc结构域二聚化,而仅形成单体。这种突变的一个例子是单体突变f405r。
显然,融合蛋白(其从n-端到c-端包含融合至人igg1fc的来自胶原蛋白18的人nc-1,或融合至来自胶原蛋白18的人nc-1的人igg1fc,其中所述人igg1fc包含一个或更多个单体突变)不再能够与另一个人igg1fc二聚化。但是,这种融合蛋白能够与其他融合蛋白寡聚化,所述其他的融合蛋白从n-端到c-端包含融合至人igg1fc的来自胶原蛋白18的人nc-1,或融合至来自胶原蛋白18的人nc-1的人igg1fc,其中所述人igg1fc包含经由nc-1的寡聚结构域包含一个或更多个单体突变,如以下实施例、图10和表4所示(分子#4,seqidno:37)。
人igg1fc进一步包含一个、两个、三个、四个、五个或更多个半衰期延长突变,诸如突变m252y、s254t和/或t256e(yte)。据报道,与野生型igg1相比,半衰期延长突变可增强与新生儿fc受体(fcrn)的结合并增加血清半衰期(dall’acqua等人,jbiolchem.2006年8月18日;281(33):23514-24.epub2006年6月21日)。
如本领域技术人员所理解的,igg1并不负责生成单体fc,并且igg4也可以在这里使用。例如,medimmune提供了一种方法,能够用在nc-1的背景下。igg4自然分裂成半抗体,然后迅速重组,该过程称为fab臂交换,并最早由vanderneutkolfschoten等,science.2007sep14;317(5844):1554-7;317(5844):1554-7,然后由labrijnetal.,natbiotechnol.2009aug;27(8):767-71.doi:10.1038/nbt.1553.epub2009jul20报道。由于这个原因,medimmune选择igg4作为产生单体fc的起点,因为与igg1相比,更容易防止这种二聚化。然而,事实证明,发现的某些突变对人igg4和igg1均起作用(甚至小鼠igg1也以这种形式起作用)。可以选择人igg4作为封闭抗体(即没有adcc或cdc)和igg1以激活免疫应答。在nc-1-fc的情况下,本发明人还没有证据表明nc-1多少的寡聚化能保留该化合物的亲合力并保持该化合物的adcc潜力。如果adcc不起作用,那么也可能会采用基于igg4的策略。
因此,在本发明的所述方法的另一个实施方案中,能够使用包含至少一个单体突变和一个或更多个半衰期延长突变的人igg4fc代替人igg1fc;参见,例如,us2013/0177555。
除了包含单体突变f405r和半衰期延长突变m252y,s254t和t256e(shanl,colazetm,rosenthalkl,yux-q,beejs,fergusona,etal.(2016)generationandcharacterizationofanigg4monomericfcplatform.plosone11(8):e0160345.https://doi.org/10.1371/journal.pone.0160345)的人igg1fc,到目前为止,本发明人已经测试了wilkinson等(mabs.2013may1;5(3):406–417;us20130177555)报道的方法,作为备选。
ying等人呈现的研究提供了其他可能的替代方法(thejournalofbiologicalchemistry287,19399-19408;andtianleiying,yangfeng,yanpingwang,weizaochen&dimiters.dimitrov(2014)。展示独特的fc受体相互作用的单体igg1fc分子可用于治疗炎症介导的疾病mabs,6:5,1201-1210),us2013/0177555和us9,200,060。
优选地,人igg1fc包含单体突变f405r和半衰期延长突变m252y、s254t和t256e(yte)。
前述融合蛋白在允许形成蛋白寡聚体的条件下在宿主细胞中表达,其中所述蛋白寡聚体包含至少两个或更多个单体人nc-1-fc蛋白。
优选地,单体人nc-1-fc蛋白包含seqidno:37或由其组成。
最后,通过本领域公知的方法从宿主细胞获得包含至少两个或更多个单体的人nc-1-fc蛋白的蛋白寡聚体;参见,例如,sambrooketal.,molecularcloning:alaboratorymanual/sambrook,joseph;russell,davidw.--.3rded.--newyork:coldspringharborlaboratory,2001;ausubeletal.,currentprotocolsinmolecularbiology。如以下实施例所示,本发明人已经接受了由三个nc1-fc(单体)组成的三聚体分子。
在一个优选的实施方案中,本发明的方法是体外方法。
本发明人可以使用携带半衰期延长突变m252y,s254t和t256e的单体突变f405r和yte突变的igg1fc结构域成功地制备单体nc-1-fc融合蛋白(参见,例如,wo2013/096291a2)(参见图10和表4,分子#4,seqidno:37)。经由nc-1低聚结构域的低聚化导致三聚体nc-1的形成,每个nc-1具有一个单体fc结构域。通过瞬时表达(30mg/l)可以高效生产这种融合蛋白。在标准条件下,单体fc无法引发抗体依赖性细胞介导的细胞毒性(adcc)。目前正在研究如果经由nc-1的fc结构域的三聚化能够增加亲合力,可能导致抗体依赖性细胞介导的细胞毒性(adcc)。
综上所述,迄今为止近十年,本发明人在许多构建体和经典标签后都失败后,终于设法实现了用于nc-1的大规模生产的系统。另外,发明人意外地发现,与同源二聚体fc-内皮抑制素(fc-es,us8703908b2,us20130165634a1)相比,本文给出的具有异源二聚体nc-1-fc或单体nc-1-fc的蛋白寡聚体在初始临床前肿瘤模型和纤维化研究中显示出更高的功效,如以下实施例以及图7和8所证实的。
如本文所用,术语“蛋白质”或“多肽”或“(多)肽”或“肽”(如果没有另外说明,所有术语可互换使用)包括基本上不含其他宿主细胞多肽的分离的和/或纯化的(多)肽。本文提及的术语“肽”包含至少2,3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,45,50,60,70,80,90,100,150,200,250,300个或甚至更多个氨基酸残基,其中一个的alpha羧基基团与另一个的alpha氨基基团结合。本文使用和设想的蛋白质或肽的翻译后修饰是新形成的蛋白质或肽的修饰,并且可以涉及氨基酸的缺失、置换和添加,某些氨基酸的化学修饰,例如对某些氨基酸的酰胺化,乙酰化,磷酸化,糖基化,焦谷氨酸形成,甲硫氨酸上磺基的氧化/还原,或类似的小分子的添加。
本文所用的术语“蛋白质”或“肽”涵盖模拟肽(peptidomimetics)。如本领域已知,模拟肽是这样的化合物,其基本元素(药效团)在3d空间中模仿天然肽或蛋白质,并保留与生物靶标(如纤连蛋白,vegf,mmp-2和/或mmp-9))相互作用的能力并产生相同的生物学效应(例如,抗纤维化活性,抗血管发生活性,抗侵入/抗转移活性,降低血管通透性活性,抗炎和抗致瘤活性,抗体依赖性细胞介导的细胞毒性(adcc)的诱导);参见例如vagneretal.2008,currentopinioninchemicalbiology12,pages292–296的综述。设计肽模拟物以避免与天然肽相关的一些问题,例如对蛋白水解的稳定性(生物活性持续时间)和差的生物利用度。某些其他特性,例如对上述生物靶标的选择性或生物活性的效力,如上述生物活性,通常可以实质上地改善。
如本文所用的蛋白质或肽修饰包括本文所述的(多)肽的合成实施方案。此外,类似物(非肽有机分子),衍生物(用公开的(多)肽序列开始获得的化学官能化的(多)肽分子)和这些蛋白质的变体(同源物)可用于本文所述的手段和方法和医学与诊断用途。本公开的每个(多)肽由氨基酸的序列组成,其可以是天然存在的或其他的l-和/或d-氨基酸。(多)肽可以通过多种化学技术进行修饰,以产生具有与未修饰的(多)肽基本相同生物学活性(例如,抗纤维化活性,抗血管发生活性,抗侵袭/抗转移活性,降低血管通透性活性,抗炎和抗致瘤活性,和/或诱导抗体依赖性细胞介导的细胞毒性(adcc))并任选地具有其他所需的特性的衍生物。例如,蛋白质的羧酸基团,无论是羧基端的还是侧链的,都可以以药学上可接受的阳离子的盐的形式提供或被酯化以形成c1-c16酯,或转化为下式nr1r2的酰胺,其中r1和r2各自独立地为h或c1-c16烷基,或结合形成杂环,诸如5元或6元环。无论氨基端还是侧链,多肽的氨基可以是药学上可接受的酸加成盐的形式,诸如hcl,hbr,乙酸,苯甲酸,甲苯磺酸,马来酸,酒石酸和其他有机盐,或者可以被修饰为c1-c16烷基或二烷基氨基或进一步转化为酰胺。可以使用公认的技术将多肽侧链的羟基基团转化为c1-c16烷氧基或c1-c16酯。多肽侧链的苯基和酚环可以被一个或多个卤素原子(例如氟,氯,溴或碘)取代,或者被c1-c16烷基,c1-c16烷氧基,羧酸及其酯,或这种羧酸的酰胺取代。多肽侧链的亚甲基基团可以延伸至同源的c2-c4亚烷基。硫醇可以用许多公认的保护基团中的任何一种(例如乙酰胺基团)保护。本领域技术人员还将认识到将环状结构引入本发明的(多)肽中以选择和提供对结构的构象限制(这导致增强的稳定性)的方法。
本文所提及的蛋白质或(多)肽也可以是融合蛋白。如本文所用的术语“融合蛋白”表示嵌合蛋白(字面上,由来自不同来源的部分组成),其通过连接最初编码分开的蛋白质的两个或更多个基因而产生。该融合基因的翻译产生具有源自每种原始蛋白质的功能特性的单个或多个多肽。例如,本发明的用于产生包含至少两种异源二聚体人nc-1-fc蛋白的蛋白寡聚体的方法使用融合蛋白,所述融合蛋白从n-至c-端包含,融合至具有“杵”突变的人igg1fc的来自胶原蛋白18的人nc-1。然后,此融合蛋白与包含适当“臼”突变的igg1fc结构域异源二聚化,如图9所示。为了提供融合蛋白的另一个实例,本发明的用于产生包含至少两个单体人nc-1-fc蛋白的蛋白寡聚体的方法使用融合蛋白,所述融合蛋白从n-至c-端包含,融合至人igg1fc的来自胶原蛋白18的人nc-1,其中所述人igg1fc包含单体突变f405r和半衰期延长突变m252y、s254t和t256e。
如本文定义的融合蛋白可以通过本领域技术人员众所周知的化学合成或重组分子生物学技术来制备。这经必要的变更适用于从宿主细胞或上清液中分离融合蛋白。参见,例如,sambrooketal.,molecularcloning:alaboratorymanual/sambrook,joseph;russell,davidw.--.3rded.--newyork:coldspringharborlaboratory,2001;ausubel,currentprotocolsinmolecularbiology,greenpublishingassociatesandwileyinterscience,n.y.(1994)。
本文定义的融合蛋白还能够包含纯化标签、标记或另一种治疗剂,诸如抗纤维化剂,抗血管发生剂和/或抗致瘤剂等。本文所提及的“标记”是可检测的化合物或组合物,其直接或间接缀合至另一分子,例如本文所定义的融合蛋白或蛋白寡聚体,以促进该分子的检测。标记的具体非限制性实例包括本领域已知的荧光标记、酶促连接和放射性同位素。在一个实施方案中,在nc-1和fc结构域之间可以存在蛋白酶切割位点和/或接头(即蛋白酶切割位点;或接头;或蛋白酶切割位点和接头两者;或包含蛋白酶切割位点的接头)。如果需要,蛋白酶切割位点可以用来通过用诸如肠激酶或凝血酶的蛋白酶处理来切掉fc结构域。例如,fc结构域可以用作表达和纯化的标签,而nc-1可以在用蛋白酶切割后分离。如本领域技术人员所公知的,除了在将功能性结构域连接在一起(如在柔性和刚性接头中)或在体内释放游离的功能性结构域(如在体内可裂解的接头中)的基本作用外,接头还可为该融合蛋白的生产提供许多其他优点,例如提高生物活性、提高表达产量并实现所需的药代动力学特征。该接头可以是,例如蛋白质/肽接头,诸如多聚甘氨酸接头或本领域已知的其他接头(参见,例如,chenetal.,advdrugdelivrev.2013;65(10):1357–1369)。显然,该接头可以以包含蛋白酶切割位点的方式来设计。如本领域技术人员所理解的,本文定义的融合蛋白不包含用于临床使用的蛋白酶切割位点和/或接头,或者在临床使用前和用于临床使用从融合蛋白中去除的蛋白酶切割位点和/或接头。
胶原18由一个中心的、间断的三螺旋结构域,在n末端(nc-11结构域)和c末端(nc-1结构域)侧翼的较大的非三螺旋状球状结构组成(ohetal.,pnas1994,91,4229;ohetal.,genomics1994,19,494;abeetal.1993,biochem.biophys.res.commun.196,576)。xviii型胶原属于胶原超家族的独特且新颖的亚类,已针对其提出名称“multiplexin家族”。
oh等人已经描述了小鼠和人胶原18蛋白的克隆(同上)。小鼠胶原蛋白18的核苷酸和氨基酸序列在登录号nm_001109991.1中示出,而相应的人序列在nm_030582.3中示出。此外,小鼠和人胶原蛋白18的氨基酸序列分别示于seqidno:1和2。
本文所用的“nc-1结构域”(或简称为nc-1或nc1)源自或来自胶原蛋白18的c-端,并包括(i)n-端缔合区(约50个氨基酸残基)(ii)中央蛋白酶敏感的铰链区(约70个氨基酸残基)和/或(iii)c-端稳定的内皮抑制素结构域(约180个氨基酸残基)(sasakietal.,1998,emboj.17,4249;wo2013/026913和wo2017/093569)。如本文所用,nc-1结构域(或简称为nc-1或nc1)优选包含(i)n-端缔合区,(ii)中央蛋白酶敏感铰链区和(iii)c-端稳定的内皮抑制素结构域。在其他优选的实施方案中,本文所用的nc-1结构域包含(i)n-端缔合区和(iii)c-端稳定的内皮抑制素结构域,其中缺少中央蛋白酶敏感铰链区(ii)。由于缺少对蛋白酶敏感的该区域,因此不具有(ii)中央蛋白酶敏感铰链区的此类nc-1结构域对于临床和治疗目的特别有用。
小鼠胶原18的nc-1结构域的氨基酸序列描述于seqidno:3,而人胶原18序列的nc-1结构域的相应序列显示于seqidno:4。优选胶原蛋白18的nc-1结构域是人的或源自人的。
如本文所用,“源自”nc-1结构域的(多)肽意指这样的(多)肽与nc-1结构域中天然(多)肽的相应氨基酸序列相同或相差1,2,3,4,5,6,7,8,9,10,15,20,25,30,35,40,50或甚至更多个氨基酸残基,同时至少保持(甚至超过)相应nc-1结构域的生物学活性(如本文其他地方所述),如寡聚化特性,抗纤维化活性,抗血管发生活性,抗侵袭/抗转移活性,降低血管通透性活性,抗炎和/或抗致瘤活性。例如,与天然人nc-1结构域相比,源自人胶原蛋白18的nc-1结构域能够携带1、2、3、4、5或甚至更多个突变。这样的突变是本领域众所周知的,并且包括例如天然(或野生型)人nc-1的核酸或氨基酸序列中的取代、添加和/或缺失。用于确定此类生物活性的测试描述于例如wo2013/026913和wo2017/093569。提及的术语“源自”人nc-1结构域的(多)肽包含人nc-1结构域的变体。优选地,所述变体序列与seqidno:4所示的人nc-1结构域的特定氨基酸序列在整个长度上具有至少50%,至少60%,至少70%,至少75%,至少80%,至少85%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%或至少99%同一性。进行序列比对和确定序列同一性的手段和方法是本领域已知的,并在本文其他地方进行了描述。
包含seqidno:4中所示氨基酸序列的约10至约60个氨基酸残基的人nc-1结构域的“缔合结构域”或“缔合区”或“寡聚结构域”(全部术语可互换)负责nc-1单体的非共价三聚化以形成球状三聚体。因此,该缔合结构域起三聚化结构域的作用。对蛋白水解切割敏感的“铰链区”包含seqidno:4中所示氨基酸序列的约61至约129个氨基酸残基。紧密的“内皮抑制素结构域”包含seqidno:4所示氨基酸序列的约130至约308个氨基酸残基;参见例如sasaki,loc.cit.;kuo2001,jcb152,1233;tjinetal.2005,cancerres65,3656。内皮抑制素结构域包含介导与锌的结合的并位于内皮抑制素的n末端的锌结合位点(dingetal.,1998,pnas95,10443;us7,524,811)。有趣的是,这种锌结合位点已显示是内皮抑制素的抗肿瘤/抗血管发生活性的原因(boehmetal.,1998,biochem.biophys.res.commun.252,190)。nc-1结构域中的缔合区和内皮抑制素结构域通过铰链区连接(参见sasaki等,同上)。已发现铰链区被例如基质金属蛋白酶(mmp)切割,例如mmp-3,-7,-9,-13和-20(heliasvaaraetal.,expcellres2005,307,192)。
本文所用的术语“fc结构域”或“fc区”是指片段可结晶区域,所述片段可结晶区域是抗体或免疫球蛋白的尾区,所述尾区与细胞表面受体即fc受体和补体系统的一些蛋白质相互作用。该特性允许抗体激活免疫系统。在igg、iga和igd抗体同种型中,fc结构域由源自抗体的两条重链的第二和第三恒定结构域的两个相同的蛋白质片段构成;在每条多肽链中igm和igefc结构域含有三个重链恒定结构域(ch结构域2-4)。igg的fc结构域携带高度保守的n-糖基化位点。fc片段的糖基化对于fc受体介导的活性是必需的。与该位点附接的n-聚糖主要是复杂类型的核心-岩藻糖基化双触角(diantennary)结构。此外,少量这些n-聚糖还携带二分glcnac和α-2,6连接的唾液酸残基。已经发现免疫球蛋白的fc结构域与蛋白质的融合增强哺乳动物细胞中融合蛋白的产生和分泌(见,例如,loetal.,1998,proteineng.11,495,caponetal.,1989,nature337,525)。此外,血管发生抑制剂与免疫球蛋白fc结构域的连接已显示出增加所述抑制剂的半衰期(参见,例如,caponetal.1989,nature337,525;gordonetal.,2001,j.clin.oncol.19,843;holashetal.,2002,proc.natl.acad.sci.usa99,11393)。但是,fc结构域不仅可以用于纯化、增溶和/或检测目的,而且可以有利地改变融合蛋白或蛋白寡聚体的生物学和药代动力学特性,如本文和以下实施例所述。在一个实施方案中,如果需要,可以通过用诸如肠激酶或凝血酶的蛋白酶处理来切掉fc结构域。对于本领域技术人员显而易见的是,原则上,任何igg同种型都可用于生成本文限定的蛋白寡聚体或融合蛋白。甚至可以使用igg的fc结构域的亚片段或单链以延长本文所述的蛋白寡聚体或融合蛋白的半衰期或寡聚化。
优选地,本文所提及的fc结构域来自人igg或源自人igg,诸如人igg1或igg4,更优选地来自人igg1或源自人igg1(bergersandjavaherianscience1999;leeetalclincancres2008)。来自igg1的人fc结构域包含seqidno:6或seqidno:24中所示的氨基酸序列或由其组成。
本文所用的“源自”人igg1或人igg4的fc结构域意指这样的fc结构域与人igg1fc结构域或人igg4fc结构域中的天然(多)肽的相应氨基酸序列相同或可以在1、2、3、4、5、6、7、8、9、10、15、20、25、30、35、40、50个甚至更多的氨基酸残基中不同,同时改变、维持甚至超过相应的天然人igg1fc结构域或人igg4fc结构域的生物学活性,诸如二聚化特性、fc受体结合(例如,与fcrn结合)并修饰构建体的药代动力学以及抗体依赖性细胞介导的细胞毒性(adcc)的诱导。例如,与天然(或野生型)人igg1fc结构域相比,源自人igg1fc结构域的fc结构域可携带1、2、3、4、5个或甚至更多个突变。这样的突变是本领域众所周知的,并且包括例如天然(或野生型)人iggfc结构域的核酸或氨基酸序列中的取代、添加和/或缺失。所述术语“源自”人igg1fc结构域的(多)肽包含人igg1结构域的变体。优选地,所述变体序列与seqidno:6或24所示的人igg1结构域的特定氨基酸序列在整个长度上有至少50%,至少60%,至少70%,至少75%,至少80%,至少85%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%或至少99%同一性。所述术语“源自”人igg4fc结构域的(多)肽包含人igg1结构域的变体。优选地,所述变体序列与例如us20130177555a1的图6中所示的人igg4结构域的特定氨基酸序列在整个长度上有至少50%,至少60%,至少70%,至少75%,至少80%,至少85%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%或至少99%同一性。
一方面,fc结构域是“杵臼结构”(kih)改造的fc结构域。如以下实施例所证实的,本发明人已经使用“杵臼结构”(kih)改造的fc结构域来生产包含异源二聚体nc-1-fc的蛋白寡聚体。杵臼结构是针对抗体的第三恒定结构域的经过充分验证的异源二聚化技术。基本上,该概念依赖于在大多数相互作用发生的两个ch3结构域之间的界面的修饰。将庞大的残基引入一个抗体重链的ch3结构域,其作用与钥匙类似。在另一条重链中,形成“臼”,其能够容纳这种大残基,模仿锁。得到的异源二聚体fc部分可以通过人工二硫键进一步稳定。在优化异源二聚化界面的过程中,使用噬菌体展示文库评价并最终优化各种合理设计(包括空间互补性、kih、二硫键和在ch3结构域任一侧与相反电荷的残基并置的盐桥)。通过引入以下六个突变可以实现产生高于97%的异源二聚化的正确重链缔合:“杵”重链中的s354c、t366w和“臼”重链中的y349c、t366s、l368a、y407v(kleinetal.,mabs.2012nov1;4(6):653–663;ridgwayet.,proteineng.1996jul;9(7):617-21)。此外,具有kih突变的抗体的特性如(热)稳定性,fcγr结合和效应子功能(例如,抗体依赖性细胞介导的细胞毒性(adcc)、fcrn结合)和药代动力学(pk)行为不受影响。非共价相互作用以及铰链区中的二硫键驱动组装形成异二聚体并使组合异质性最小化。合适的kih改造的fc结构域描述于例如seqidno.25,26,28和30。例如,seqidno:25显示具有“杵”突变s354c/t366w的人igg1fc的氨基酸序列,并且seqidno:26描述具有“臼”突变y349c/t366s/l368a/y407v的人igg1fc的氨基酸序列。
seqidno:27显示包含经由肠激酶切割位点和接头(从n-至c-末端)融合至具有“杵”突变(s354c/t366w)的人igg1fc的人nc-1的融合蛋白质的氨基酸序列。该融合蛋白能够与具有“臼”突变y349c/t366s/l368a/y407v(seqidno:28)的人igg1fc异源二聚化。这样的异源二聚体如图9所示。
seqidno:29显示包含经由接头和肠激酶位点融合至人nc-1(从n-至c-末端)的具有“杵”突变(s354c/t366w)的人igg1fc的融合蛋白的氨基酸序列。该融合蛋白能够与具有“臼”突变y349c/t366s/l368a/y407v的人igg1fc(seqidno:30)异源二聚化。
本发明人已经发现,“杵臼结构”(kih)改造的fc结构域对于生产包含至少两种或更多个异源二聚体人nc-1-fc蛋白的蛋白寡聚体特别有用,如在以下实施例中证实的。因此,优选人免疫球蛋白的kih改造的fc结构域,诸如人igg1或人igg4,更优选人igg1的kih改造的fc结构域用于产生所述蛋白质寡聚体。
在另一方面,fc结构域是单体的,即通过突变位于igg1或igg4fc二聚化界面上的关键残基而改造的。如何产生单体igg1或单体igg4是本领域已知的;参见,例如,wangetal.,front.immunol.,13november2017,https://doi.org/10.3389/fimmu.2017.01545。本发明人已经发现,单体fc结构域在制备包含两个或更多个单体nc-1-fc融合蛋白的蛋白寡聚体中特别有用,如以下实施例所证实的。优选地,fc结构域来自igg1,并且在单体nc-1-fc融合蛋白中包含对应于单体突变的突变f405r以及对应于半衰期延长突变的突变m252y、s254t和t256e。进一步的单体nc-1-fc融合蛋白将使用selexis的rcb技术进行鉴定。
术语“寡聚体”通常是指在生物化学中通过一些大分子如蛋白质或核酸的非共价键合形成的大分子复合物。根据定义,二聚体是由两个通常非共价结合的分子如蛋白质或肽形成的大分子复合物。这种复合物也可以由蛋白质结构域形成,所述蛋白质结构域是蛋白质序列的一部分并且可以独立于蛋白质链的其余部分进化,发挥功能和存在的结构。同源二聚体由两个相同的分子形成。潜在的过程称为同源二聚化。异源二聚体由两个不同的分子构建,这两个分子通过异二聚化形成。如本领域已知的,除二硫键外,生物化学中的大多数二聚体或三聚体不通过共价键连接。一些蛋白质含有专门的结构域以确保二聚化、三聚化或寡聚化,这些专门的结构域是所谓的二聚化、三聚化或寡聚化结构域,如本文定义,并且是本领域熟知的。举一个例子,二聚化可以通过免疫球蛋白的fc结构域或通过二硫键或两者或本领域已知的其他方式介导。例如,图9中显示了异源二聚体人nc-1-fc蛋白。在此,nc-1-fc杵与fc臼二聚化。
因此,三聚体是由三个通常非共价结合的肽,蛋白质或蛋白质结构域形成的大分子复合物。同源三聚体由三个相同的分子形成,而异源三聚体由三个不同的分子构成。例如,胶原蛋白18是同源三聚体蛋白。四聚体由四个分子组成,五聚体由五个分子组成,依此类推。在这些情况下,如上所述,复合物的形成通常由低聚结构域介导。例如,对于三聚化,可以使用nc-1结构域内的天然缔合区(或低聚结构域)来介导胶原18的nc-1的三聚化,因为胶原18的nc-1结构域内的天然缔合区作为三聚化结构域起作用。
在本发明的上下文中,“寡聚体”应理解为“蛋白寡聚体”,其包含一些单体单元,例如,两个,三个,四个,五个或甚至更多个单体单元。因此,寡聚体可以是例如二聚体,三聚体,四聚体,五聚体等。优选地,寡聚体是同源二聚体,同源三聚体等。单体单元(或简称单体)可以是例如异源二聚体人nc-1-fc蛋白或能够通过nc-1的低聚结构域形成三聚体的单体nc-1-fc。然后,单体单元“异源二聚体人nc-1-fc蛋白”经由nc-1的低聚(或缔合)结构域形成三聚体;参见图9。同样,单体单元“单体nc-1-fc”经由nc-1的低聚结构域形成三聚体,如图10所示。
本发明的蛋白寡聚体具有以下由寡聚nc-1介导的生物活性中的至少一种,优选至少两种,更优选至少三种,四种,五种,特别优选地所有:抗纤维化活性,抗血管发生活性,抗侵袭/抗转移活性,降低血管通透性活性,抗炎和/或抗致瘤活性。上述生物学活性和特性的测试是本领域已知的;参见例如wo2013/026913和wo2017/093569。此外,本发明的蛋白寡聚体优选具有经由二聚化的或寡聚化的fc结构域诱导抗体依赖性细胞介导的细胞毒性(adcc)的能力。用于测试adcc的手段和方法在文献中有很好的描述(参见,例如,yamashitaetal.,scientificreportsvolume6,articlenumber:19772(2016);doi:10.1038/srep19772;parekhetal.,mabs.2012may1;4(3):310–318.doi:10.4161/mabs.19873;chungetal.,jimmunolmethods.2014may;407:63-75.doi:10.1016/j.jim.2014.03.021.epub2014apr3;tadaetal.(2014)plosone9(4):e95787.https://doi.org/10.1371/journal.pone.0095787;
https://www.essenbioscience.com/media/uploads/files/8000-0580-a00-immune_cell_killing--tumor_spheroids_assay_protocol.pdf)。
如本文所定义的蛋白寡聚体可以通过化学合成或重组分子生物学技术来制造,并且可以通过本领域公知的方法从宿主细胞或细胞培养上清液中分离;参见,例如,sambrooketal.,molecularcloning:alaboratorymanual/sambrook,joseph;russell,davidw.--.3rded.--newyork:coldspringharborlaboratory,2001。
如本文所用的术语“蛋白寡聚体”还包括蛋白质制剂,其包含蛋白寡聚体和额外地其他蛋白质、试剂或化合物。例如,如本文所定义的所述蛋白寡聚体可以使用一种或更多种其他抗纤维化、抗血管发生和/或致瘤蛋白质、化合物或试剂以组合方案施用给有需要的受试者,优选人受试者。药物组合通常比单独使用一种药物更有效地抵抗本文所定义的疾病。举例来说,如本文所定义的蛋白质寡聚体可以与制管张素或制管张素融合蛋白(如与免疫球蛋白的fc结构域连接的制管张素)组合使用,或与跟纤维化过程相关的其他途径的抑制剂(包括例如tgf-β,pdgf,vegf,mtor,ctgf,整联蛋白,基质金属蛋白酶,抗炎剂如环加氧酶,ikk/nfkb,jak/stat和/或pi3k信号转导的类固醇抑制剂)一起使用。
本发明进一步涉及用于治疗,改善或预防与血管发生有关的疾病的包含至少两个,优选三个异源二聚体人nc-1-fc蛋白的蛋白寡聚体,其中所述异源二聚体人nc-1-fc包括:(i)一种融合蛋白,其从n-至c-端包含,融合至包含“杵”突变s354c/t366w的人igg1fc的来自胶原18的人nc-1,或融合至来自胶原蛋白18的人nc-1的包含“杵”突变s354c/t366w的人igg1fc,和(ii)具有“臼”突变y349c/t366s/l368a/y407v的人igg1fc。
优选地,所述具有“杵”突变s354c/t366w的人igg1fc包含seqidno:25,和具有“臼”突变y349c/t366s/l368a/y407v的人igg1fc包含seqidno:26。
更优选地,融合蛋白(i),其从n-至c-端包含,融合至具有“杵”突变的人igg1fc的来自胶原蛋白18的人nc-1包含seqidno:27,29,或31或由其组成,且(ii)的具有“臼”突变的人igg1fc包含seqidno:28,30,或32或由其组成。
特别优选的是,融合蛋白(i)包含seqidno:31或由其组成,所述融合蛋白从n-至c-端包含融合至具有“杵”突变的人igg1fc的来自胶原蛋白18的人nc-1,且(ii)的具有“臼”突变的人igg1fc包含seqidno:32或由其组成(参见图9和表1,分子#1)。
如以下实施例以及图7和图8所证实的,已经发现这种蛋白寡聚体作为用于治疗、改善或预防与血管发生有关的疾病的药物组合物是特别有用的。
如本文所用的“药物组合物”能够以治疗有效剂量用于本文提及的疾病的非人或优选人的治疗。本文所指的“受试者”优选是患有本文所提及的疾病的人。
本文定义的包含至少两个异源二聚体人nc-1-fc蛋白的蛋白寡聚体,本发明的单体人nc-1-fc蛋白或本发明的包含至少两个单体人nc-1-fc蛋白的蛋白寡聚体是药物组合物或药物的活性成分(两个术语可互换使用),并且在一个方面,以通过根据常规程序将药物与标准药物载体组合而制备的常规剂型施用。这些程序可以涉及混合、制粒和压缩或溶解适合所需制剂的成分。应该理解的是,药学上可接受的载体或稀释剂的形式和特征取决于与其组合的活性成分的量、给药途径和其他众所周知的变量。
载体在与制剂的其他成分相容并且对其接受者无害的意义上必须是可接受的。所用的药物载体可以包括固体、凝胶或液体。固体载体的实例是乳糖、白土、蔗糖、滑石、明胶、琼脂、果胶、阿拉伯胶、硬脂酸镁、硬脂酸等。液体载体的实例是磷酸盐缓冲盐溶液、糖浆、油、水、乳剂、各种类型的润湿剂等。类似地,载体或稀释剂可以包括本领域公知的延时材料,如单独的单硬脂酸甘油酯或二硬脂酸甘油酯或与蜡一起。所述合适的载体包括上面提到的那些和本领域公知的其他载体,参见例如remington′spharmaceuticalsciences,mackpublishingcompany,easton,pennsylvania。
选择稀释剂以不影响药物组合物的生物活性。所述生物活性已经在本文其他地方定义。这些稀释剂的实例是蒸馏水、生理盐水、林格氏溶液(ringer'ssolution)、葡萄糖溶液和汉克氏溶液(hank'ssolution)。此外,药物组合物或制剂还可包括其他载体,佐剂或无毒、非治疗性、非免疫原性稳定剂等。
优选地,将本文定义的包含至少两个异源二聚体人nc-1-fc蛋白的蛋白寡聚体,本发明的单体人nc-1-fc蛋白或本发明的包含至少两个单体人nc-1-fc蛋白的蛋白寡聚体配制成药物组合物,其可以通过标准途径施用。通常,药物组合物可以通过局部,透皮,腹膜内,颅内/鞘内,玻璃体内,脑室内,脑内,阴道内,子宫内,口服,直肠或肠胃外(例如静脉内,脊柱内,皮下或肌肉内)途径施用。
优选地,本文定义的包含至少两个异源二聚体人nc-1-fc蛋白的蛋白寡聚体,本发明的单体人nc-1-fc蛋白或本发明的包含至少两个单体人nc-1-fc蛋白的蛋白寡聚体通过静脉内,皮下,颅内/鞘内,玻璃体内或腹膜内施用。
治疗有效剂量是指用于药物组合物中的本文定义的包含至少两个异源二聚体人nc-1-fc蛋白的蛋白寡聚体,本发明的单体人nc-1-fc蛋白或本发明的包含至少两个单体人nc-1-fc蛋白的蛋白寡聚体的量,所述量预防,改善或治疗伴随本说明书中提及的疾病的症状。所述化合物的治疗功效和毒性可以通过细胞培养或实验动物中的标准药物程序,例如ed50(50%群体中治疗有效的剂量)和ld50(50%群体的致死剂量)来确定。治疗和毒性作用之间的剂量比是治疗指数,它可以表示为比率ld50/ed50。
剂量方案将由主治医师和其他临床因素确定。如在医学领域中众所周知的,任何一个患者的剂量取决于许多因素,包括患者体型、体表面积、年龄、待给药的具体化合物、性别、给药时间和途径、一般健康状况和同时施用的其他药物。可以通过定期评估来监测进展。
优选地,本文定义的包含至少两个异源二聚体人nc-1-fc蛋白的蛋白寡聚体,本发明的单体人nc-1-fc蛋白或本发明的包含至少两个单体人nc-1-fc蛋白的蛋白寡聚体以约1至100mg/kg体重的浓度施用。更优选地,所述浓度为约5至75mg/kg或约10至50mg/kg,最优选约15mg/kg体重。
本文提及的药物或药物组合物至少施用一次,以治疗或改善或预防本说明书中所述的疾病。然而,所述药物可以施用一次以上,例如,两次,三次,四次,五次,六次或甚至更频繁地施用。例如,本文所指的药物或药物组合物可以每天一次或每第二、第三、第四、第五或第六天或每周一次施用。也可以每第二周、每三周或每四周施用一次。
优选的靶标血浆浓度为约5至100、10至50或15微克/ml血液。
具体的药物组合物以药学领域熟知的方式制备,并包含与药学上可接受的载体或稀释剂混合的或以其他方式关联的至少一种上文提及的活性化合物。为了制备这些具体的药物组合物,通常将活性化合物与载体或稀释剂混合。所得制剂应适合于施用方式。剂量建议应在开处方者或使用者说明书中说明,以便根据所考虑的接受者预测剂量调整。
在本发明的另一方面,药物组合物可包含除本文定义的包含至少两个异源二聚体人nc-1-fc蛋白的蛋白寡聚体外,还包含本发明的单体人nc-1-fc蛋白或本发明的包含至少两个单体人nc-1-fc蛋白的蛋白寡聚体外的药物,其在药物配制过程中加入药物中。例如,它可以与组合方案中的制管张素一起使用。此外,另一方面设想了与最近批准的纤维化调节剂如vegf/pdfgrtki(例如nindetanib),tgf-beta信号传导的特异性和非特异性抑制剂(perfinidone)和整联蛋白信号传导的调节剂(西仑吉肽,或抗alphav阿比珠单抗)或炎症的调节剂(白细胞浸润,细胞因子抑制剂,抗亚群的抗体)的组合。因此,在本文定义的包含至少两个异源二聚体人nc-1-fc蛋白的蛋白寡聚体,本发明的单体人nc-1-fc蛋白或本发明的包含至少两个单体人nc-1-fc蛋白的蛋白寡聚体的优选的实施方案中,所述蛋白寡聚体或单体人nc-1-fc蛋白进一步包含制管张素(us8,206,718)。在特定的实施方案中,制管张素是fc-制管张素或制管张素-fc融合蛋白,优选人融合蛋白。
应理解,药物组合物的制剂在gmp标准化条件等下进行,以确保药剂的质量,药物安全性和有效性。
本文所用的术语“治疗”表示通过向需要的受试者施用本文定义的蛋白寡聚体或融合蛋白,改善或甚至消除与本文提及的疾病相关的一种或更多种症状。
本文提及的术语“减轻”是指通过施用本文所述的蛋白寡聚体或融合蛋白,改进或改善在本文提及的疾病的行为。改善也可以被视为疾病进展的减缓或停止。
本文所用的术语“预防”是指通过施用本文定义的蛋白寡聚体或融合蛋白来避免本文提及的疾病的发生或再发生。
血管发生相关疾病优选地选自由以下各项组成的组:血管发生依赖性癌症,包括实体瘤,黑素瘤,肿瘤转移瘤,血源性肿瘤(如白血病),良性肿瘤(如血管瘤),听神经瘤,神经纤维瘤,沙眼,化脓性肉芽肿;类风湿性关节炎;银屑病;眼部血管发生疾病如糖尿病性视网膜病变,早产儿视网膜病变,黄斑变性,角膜移植片排斥,新生血管性青光眼,晶状体后纤维组织形成,发红(rubeosis);osler-webber综合征;心肌血管发生;斑块新血管形成;毛细管扩张;血友病关节;血管纤维瘤;伤口肉芽;过度或异常刺激内皮细胞的疾病如肠粘连,动脉粥样硬化,硬皮病,肥厚性疤痕(疤痕疙瘩);具有血管发生作为病理结果的疾病,例如猫抓病(rocheleminaliaquintosa)和溃疡(幽门螺杆菌)。
与血管发生有关的疾病在wo2013/026913中进行了描述和表征。wo2013/026913的定义和实施方式经必要的变更适用于本申请。在wo2017/093569中描述的是-或可被认为是-血管发生相关疾病的疾病被明确排除和放弃在本发明的医学用途范围之外,其中本文限定的包含至少两个异源二聚体人nc-1-fc蛋白寡聚体用于治疗、改善或预防本文定义的与血管发生有关的疾病。
本发明还涉及单体nc-1-fc蛋白,其包含来自胶原蛋白18的人nc-1和来自人igg1的fc结构域,其中来自人igg1的fc结构域包含至少一个单体突变,和一个或更多个半衰期延长突变。例如,单体突变可以是f405r,而半衰期延长突变可以是m252y,s254t和/或t256e。
优选地,本发明涉及单体nc-1-fc蛋白,从n-至c-端包含来自胶原蛋白18的人nc-1和来自人igg1的fc结构域,或来自人igg1的fc结构域和来自胶原蛋白18的人nc-1,其中来自人igg1的fc结构域包含单体突变f405r和半衰期延长突变m252y,s254t和t256e。更优选地,单体nc-1-fc蛋白包含seqidno:37或由其组成(参见图10,表4,分子#4)。
在另一个实施方案中,单体nc-1-fc蛋白包含来自胶原蛋白18的人nc-1和来自人igg4的fc结构域,其中来自人igg4的fc结构域包含至少一个单体突变和一个或更多个半衰期延长突变。
本发明进一步提供蛋白寡聚体,所述蛋白寡聚体包含至少两个包含来自胶原蛋白18的人nc-1和来自人igg1或人igg4的fc结构域的单体人nc-1-fc融合蛋白,其中来自人igg1或igg4的fc结构域包含至少一个单体突变和一个或更多个半衰期延长突变。
优选地,人igg1fc中的单体突变是f405r,半衰期延长突变是m252y,s254t和t256e。
另外,本发明提供了蛋白寡聚体,所述蛋白寡聚体通过产生包含至少两个单体人nc-1-fc蛋白质的蛋白寡聚体的方法来制备,其中所述方法包括:
a)在允许形成包含至少两个,且优选三个单体人nc-1-fc蛋白的蛋白寡聚体的条件下培养表达融合蛋白的宿主细胞,所述融合蛋白从n-至c-端包含,融合至人igg1fc的来自胶原蛋白18的人nc-1,或来自人igg1的fc结构域和来自胶原蛋白18的人nc-1,其中所述人igg1fc包含至少一个单体突变,优选地单体突变f405r,和一个或更多个半衰期延长突变,优选半衰期延长突变m252y、s254t和t256e,以及
b)从步骤a)的所述宿主细胞中获得包含至少两个,且优选三个单体人nc-1-fc蛋白的蛋白寡聚体。
优选地,包含至少两个单体人nc-1-fc融合蛋白的蛋白寡聚体结合纤连蛋白,vegf,mmp-2和/或mmp-9,更优选结合这些蛋白中的每一个。
优选地,包含在包含至少两个单体人nc-1-fc融合蛋白的蛋白寡聚体中的fc结构域能够诱导抗体依赖性细胞介导的细胞毒性(adcc)。
本发明还涉及包含至少两个单体nc-1-fc蛋白的蛋白寡聚体,其中所述单体nc-1-fc蛋白从n-至c-端包含来自胶原蛋白18的人nc-1和来自人igg1的fc结构域,或来自人igg1的fc结构域和来自胶原蛋白18的人nc-1,其中来自人igg1的fc结构域包含至少一个单体突变和一个或更多个半衰期延长突变,其用作药物。
已发现本发明的包含三个单体nc-1-fc蛋白的这种蛋白寡聚体作为用于治疗、改善或预防肺纤维化模型中纤维化的药物组合物是赫然有用的,如以下实施例和图16(参见nc-1-fc(单体))所证实的。
本发明还涉及包含至少两种单体nc-1-fc蛋白的蛋白寡聚体,其中所述单体nc-1-fc蛋白在n-至c-端包含来自胶原蛋白18的人nc-1和来自于人igg4的fc结构域,或来自人igg4的fc结构域和来自胶原蛋白18的人nc-1,其中来自人igg4的fc结构域包含至少一种单体突变和一个或更多个半衰期延长突变,用作药物。
优选地,蛋白寡聚体包含至少两个单体人nc-1-fc融合蛋白,所述融合蛋白从n-至c-端包含来自胶原蛋白18的人nc-1和来自人igg1的fc结构域,或来自人igg1的fc结构域和来自胶原蛋白18的人nc-1,其中来自人igg1的fc结构域包含单体突变f405r和半衰期延长突变m252y,s254t和t256e。
另外,本发明涉及通过本发明的用于生产包含至少两个单体人nc-1-fc蛋白质的蛋白寡聚体的方法生产的蛋白寡聚体,所述蛋白寡聚体用作药物。
优选地,该药物用于治疗、改善或预防选自以下的疾病:
(i)包含以下的血管发生相关疾病:血管发生依赖性癌症,包括实体瘤,黑素瘤,肿瘤转移瘤,血源性肿瘤(如白血病),良性肿瘤(如血管瘤),听神经瘤,神经纤维瘤,沙眼,化脓性肉芽肿;类风湿性关节炎;银屑病;眼部血管发生疾病如糖尿病性视网膜病变,早产儿视网膜病变,黄斑变性,角膜移植片排斥,新生血管性青光眼,晶状体后纤维组织形成,发红(rubeosis);osler-webber综合征;心肌血管发生;斑块新血管形成;毛细管扩张;血友病关节;血管纤维瘤;伤口肉芽;过度或异常刺激内皮细胞的疾病如肠粘连,动脉粥样硬化,硬皮病,肥大性疤痕(疤痕疙瘩);具有血管发生作为病理结果的疾病,例如猫抓病(rocheleminaliaquintosa)和溃疡(幽门螺杆菌);
(ii)纤维化或与纤维化相关疾病,包括皮肤纤维化,优选硬皮病;疤痕疙瘩或瘢痕疙瘩;肥大性疤痕;硬斑病;移植物抗宿主病导致的纤维化;上皮下纤维化;心内膜心肌纤维化;子宫纤维化;骨髓纤维化;腹膜后纤维化;肾源性系统性纤维化;手术后疤痕;哮喘;肝硬化/肝纤维化;伤口愈合异常导致的纤维化;肾小球肾炎;多灶性纤维硬化;辐射引起的纤维化,优选辐射引起的肺炎或辐射引起的肺纤维化;化疗引起的或药物引起的纤维化,例如由于mtor或egfr激酶抑制;普通或特发性肺纤维化;自身免疫性疾病引起的纤维化,例如狼疮,与肿瘤内和癌症相关的纤维化/纤维发生,器官纤维化伴随的慢性炎症,例如通过病毒刺激或移植;作为慢性肾脏疾病、长期透析或糖尿病的终末期的器官纤维化;
(iii)血管内皮生长因子(vegf)相关疾病,包括取决于vegf水平失调的良性病理生理学病况,例如湿性黄斑变性,子宫内膜异位症,支气管哮喘和糖尿病,增强的vegf诱导的血管通透性(例如脑组织照射后增强的通透性,“放射性坏死”),血管紧张的改变(例如高血压),类风湿性关节炎,以及恶性vegf依赖性疾病,例如肾细胞癌和其他vegf成瘾性肿瘤,vegf依赖性腹水发展,vegf依赖性免疫系统抑制,例如源自骨髓的细胞(bmdc)、源自骨髓的抑制细胞(mdsc)、未成熟树突状细胞等的募集和微环境训练;和
(iv)基质金属蛋白酶(mmp)相关疾病,包括良性和恶性疾病,其中mmp激活促成病理生理学,例如,在局部治疗失败率高的肿瘤(例如胶质母细胞瘤,胰腺癌,肺癌)所固有的局部肿瘤侵袭和癌症转移过程中mmp的激活,以及由治疗诱导选择压力(例如肿瘤缺氧和放疗后纤维化),自身免疫疾病和慢性炎症疾病的明显免疫反应导致的获得性增强的mmp激活。
关于包含至少两种异源二聚体nc-1-fc融合蛋白的蛋白寡聚体的本发明的治疗用途的定义、实施方案和解释,经必要的变更适用于本发明的包含至少两个单体nc-1-fc融合蛋白的蛋白寡聚体。
在一个优选的实施方案中,包含至少两个异源二聚体人nc-1-fc蛋白的蛋白寡聚体,或包含至少两个单体人nc-1-fc融合蛋白的蛋白寡聚体通过静脉内、颅内/鞘内、玻璃体内、皮下或腹膜内,优选以0.1-1mg/kg/天的浓度施用。
在另一个优选的实施方案中,包含至少两个异源二聚体人nc-1-fc蛋白的蛋白寡聚体,或包含至少两个单体人nc-1-fc融合蛋白的蛋白寡聚体,具有一种或更多种选自由以下各项组成的组的生物学活性:抗纤维化活性,抗血管发生活性,抗侵入/抗转移活性,降低血管通透性活性,抗炎和抗致瘤活性,以及诱导抗体依赖性细胞介导的细胞毒性(adcc)的能力。
在又一个实施方案中,包含至少两个异源二聚体人nc-1-fc蛋白的蛋白寡聚体,或包含至少两个单体人nc-1-fc融合蛋白的蛋白寡聚体,进一步包含制管张素,血小板反应蛋白,抗pd-1/pd-l1抗体或用于治疗、改善或预防如本文所限定的与血管发生相关的疾病、纤维化或与纤维化相关的疾病、vegf相关疾病或mmp相关疾病的另一种疗法。
除了包含至少两个单体人nc-1-fc蛋白质的蛋白质寡聚体之外,还可以使用的治疗血管内皮生长因子(vegf)相关疾病的药物,例如,包括血管通透性(例如脑组织照射后增强的通透性,“放射性坏死”)和血管紧张(如内皮素拮抗剂马西替坦(macitentan),at1/ace抑制剂)的其他调节剂,用于哮喘中的β2-拟交感神经药和皮质激素,慢性炎症/自身免疫疾病中的免疫抑制剂,针对不同vegf依赖性肿瘤和腹水的化疗和放疗,在例如肾细胞癌中使用的激酶抑制剂(mtori,例如rad001,多激酶抑制剂帕唑帕尼/西尼替尼/阿西替尼,免疫调节剂,例如检查点抑制剂抗pd-1/pd-l1)。除了包含至少两个单体人nc-1-fc蛋白质的蛋白寡聚体之外,还可以使用的治疗基质金属蛋白酶相关疾病的药物包括例如具有高局部区域治疗失败率的经放(化)疗的局部侵入性肿瘤,如胶质母细胞瘤,胰腺癌,抗炎和免疫抑制疗法(抗tnfα抗体/英夫利昔单抗,霉酚酸,环磷酰胺等),癌症治疗后肿瘤侵入或假性进展,例如复发性胶质瘤中的抗血管发生疗法,具有高mmp-2/mmp-9活性的转移性疾病(例如乳腺癌)的治疗(即激素疗法他莫昔芬,her2+疾病中的曲妥珠单抗,化疗)。
本发明进一步描述包含编码异源二聚体人nc-1-fc蛋白或单体人nc-1-fc蛋白的多核苷酸。
如本文所用的术语“多核苷酸”或“核酸”是指单链或双链dna分子以及rna分子。所述术语包括基因组dna,cdna,hnrna,mrna以及这些分子种类的所有天然存在的或人工修饰的衍生物。多核苷酸在一个方面可以是线性或环状分子。此外,除了编码异源二聚体人nc-1-fc蛋白或单体人nc-1-fc蛋白的核酸序列外,多核苷酸还可以额外包含正确转录和/或翻译所需的序列,例如5‘或3‘-utr序列。鉴于遗传密码的简并性,优化的密码子可用于编码异源二聚体人nc-1-fc蛋白或单体人nc-1-fc蛋白的核酸序列。由此,可以实现例如宿主细胞中的最佳表达。
应当理解,本发明还涵盖异源二聚体人nc-1-fc蛋白或单体人nc-1-fc蛋白的特定氨基酸序列的变体或编码它们的核酸序列,只要这些变体序列还允许形成蛋白寡聚体。通过所述变体形成的蛋白寡聚体优选具有以下生物活性中的至少一种,优选至少两种,更优选至少三种,特别优选地所有:抗纤维化活性,抗血管发生活性,抗侵袭/抗转移活性,降低血管通透性活性,抗炎和/或抗致瘤活性。此外,由所述变体形成的蛋白寡聚体优选具有诱导抗体依赖性细胞介导的细胞毒性(adcc)的能力。
在一个方面,本文使用的序列变体与前面指定的特定氨基酸序列或特定核酸序列的不同之处在于一个、两个、三个、四个、五个或更多个氨基酸或核苷酸置换,添加和/或缺失。在另一方面,所述变体序列与异源二聚体人nc-1-fc蛋白或单体人nc-1-fc蛋白的各自的两条链的特定核酸序列或氨基酸序列在整个长度上或在特定序列长度的至少一半长度上有至少50%,至少60%,至少70%,至少80%,至少85%,至少90%,至少91%,至少92%,与至少93%,至少94%,至少95%,至少96%,至少97%,至少98%或至少99%同一性。优选地所述变体序列与seqidno:27,28,29,30,31,32或37在整个长度上有至少50%,至少60%,至少70%,至少80%,至少85%,至少90%,至少91%,至少92%,至少95%,至少96%,至少97%,至少98%或至少99%同一性。本文所用的术语“同一的”是指序列同一性,其特征在于确定序列之间相同氨基酸的数目,其中序列进行比对以获得最高级匹配。它可以使用在计算机程序中编码的公开技术或方法例如blastp或fasta(altschul1990,jmolbiol215,403)来计算。在一个方面,同一性百分比值是在整个氨基酸序列上或在查询序列的至少50%的序列段上计算的。基于各种算法的一系列程序可供技术人员用于比较不同的序列。在这种情况下,needleman和wunsch或smith和waterman的算法给出了特别可靠的结果。为了进行序列比对,可以使用程序pileup(higgins1989,cabios5,151)或程序gap和bestfit(needleman1970,jmolbiol48;443;smith1981,advapplmath2,482),它们是gcg软件包(geneticscomputergroup1991,575sciencedrive,madison,wisconsin,usa53711)的部分。在本发明的另一个方面,在整个序列区域上使用具有以下设置的程序gap确定以百分比(%)表述的序列同一性值:缺口权重:50,长度权重:3,平均匹配:10.000和平均不匹配:0.000,除非另有说明,否则应始终用作序列比对的标准设置。
本发明进一步描述包含编码异源二聚体人nc-1-fc蛋白或单体人nc-1-fc蛋白的多核苷酸的载体。
优选地,载体是表达载体。
术语“载体”优选包括噬菌体,质粒,病毒或逆转录病毒载体以及人工染色体,例如细菌或酵母人工染色体。此外,该术语还涉及靶向构建体,其允许靶向构建体随机或定点整合到基因组dna中。在一个方面,此类靶标构建体包含用于如下文详细描述的同源或异源重组的足够长度的dna。在一个方面,包含所提及的多核苷酸的载体还包含用于在宿主细胞中繁殖和/或选择的选择标记。载体可以通过本领域熟知的各种技术结合到宿主细胞中。例如,质粒载体可以在沉淀物(例如磷酸钙沉淀物或氯化铷沉淀物)中引入,或者与带电荷的脂质形成复合物或者在基于碳的簇中(例如富勒烯)中引入。或者,可以通过热休克或电穿孔技术引入质粒载体。如果载体是病毒,可以在应用于宿主细胞之前使用合适的包装细胞系在体外包装。逆转录病毒载体可以是复制型或复制缺陷型。在后一种情况下,病毒繁殖通常仅在互补宿主/细胞中发生。
此外,在一个方面,上述多核苷酸与表达控制序列有效地连接,从而允许在所述载体中的原核或真核宿主细胞或其分离的级分中表达。因此,在一个方面,载体是表达载体。多核苷酸的表达包括将多核苷酸转录成可翻译的mrna。确保在宿主细胞中表达的调节元件是本领域熟知的。在一个方面,它们包含确保转录起始的调节序列和/或确保转录终止和转录物稳定化的poly-a信号。其他调节元件可包括转录增强子和翻译增强子。允许在原核宿主细胞中表达的可能的调节元件包括例如大肠杆菌中的lac-,trp-或tac-启动子,并且允许在真核宿主细胞中表达的调节元件的实例是酵母中的aox1-或gal1-启动子,或哺乳动物和其他动物细胞中的cmv-,sv40-,rsv-启动子(劳氏肉瘤病毒),cmv-增强子,sv40-增强子或珠蛋白内含子。此外,诱导型表达控制序列可用于表达载体。此类诱导型载体可包含tet或lac操纵基因序列或可通过热休克或其他环境因素诱导的序列。合适的表达控制序列是本领域熟知的。除了负责转录起始的元件之外,这些调节元件还可以包含转录终止信号,例如多核苷酸下游的sv40-poly-a位点或tk-poly-a位点。在本上下文中,合适的表达载体是本领域已知的,例如okayama-bergcdna表达载体pcdv1(pharmacia),pbluescript(stratagene),pcdm8,prc/cmv,pcdna1,pcdna3(invitrogen)或psport1(invitrogen)。优选地,所述载体是表达载体和基因转移或靶向载体。来自病毒如逆转录病毒,痘苗病毒,腺伴随病毒,疱疹病毒或牛乳头瘤病毒的表达载体可用于将多核苷酸或载体递送到靶细胞群中。可以使用本领域技术人员熟知的方法构建重组病毒载体;参见,例如,sambrook,molecularcloningalaboratorymanual,coldspringharborlaboratory(2001)n.y.andausubel,currentprotocolsinmolecularbiology,greenpublishingassociatesandwileyinterscience,n.y.(1994)中描述的技术。
本发明进一步描述了包含编码异源二聚体人nc-1-fc蛋白或单体人nc-1-fc蛋白的多核苷酸的宿主细胞。
如本文所用的术语“宿主细胞”包括原核和真核宿主细胞。在一个方面,宿主细胞是细菌细胞。在一个方面,所述细菌宿主细胞是大肠杆菌宿主细胞。这种细菌宿主细胞可用于例如复制所提及的多核苷酸或载体。
在一个方面,真核宿主细胞是这样的细胞,其包含编码异源二聚体人nc-1-fc蛋白或单体人nc-1-fc蛋白的多核苷酸或其中所述多核苷酸或载体在所述宿主细胞中表达以生成包含异源二聚体人nc-1-fc蛋白或单体人nc-1-fc蛋白的蛋白寡聚体的载体。可以将多核苷酸瞬时或稳定地引入宿主细胞中。在一个方面,真核宿主细胞可以是稳定表达多核苷酸的真核宿主细胞系的细胞。另一方面,宿主细胞是真核宿主细胞,其已被多核苷酸或载体瞬时转染并表达多核苷酸。在另一个方面,所述细胞是经遗传改造以产生蛋白质的细胞。如何通过分子生物学技术对这些细胞进行遗传改造是本领域技术人员公知的。
序列
序列显示:
seqidno:1:鼠胶原蛋白18
seqidno:2:人胶原蛋白18
seqidno:3:鼠胶原蛋白18的nc-1结构域
seqidno:4:人胶原18的nc-1结构域
eqidno:5:鼠fc结构域
seqidno:6:人fc结构域
seqidno:7:鼠superstatin
seqidno:8:鼠纤连蛋白基序
eqidno:9:鼠n-端锌结合结构域内皮抑制素
seqidno:10:人n-端锌结合结构域内皮抑制素
seqidno:11:鼠rgd基序
seqidno:12:人rgd基序
seqidno:13:人superstatin
seqidno:14:人superstatin,其在位置1和3的his被ala替代
seqidno:15:人superstatin,其在第7位的gln被cys替代
seqidno:16:人superstatin,其“rgd”基序被“rad”基序替代
eqidno:17:纤连蛋白的鼠整联蛋白结合基序
seqidno:18:鼠内皮抑制素(鼠)
seqidno:19:人内皮抑制素(人类)
seqidno:20:mp1肽(鼠;n-端)
seqidno:21:内皮抑制素的e4肽(人;c-端)
seqidno:22:内皮抑制素的hp1肽(人;n-端)
seqidno:23:肠激酶切割位点
seqidno:24:野生型人igg1的fc序列
seqidno:25:fc序列“杵”人igg1fc:s354c/t366w
seqidno:26:fc序列“臼”人igg1fcy349c/t366s/l368a/y407v
seqidno:27:具有“杵”突变(s354c/t366w)的nc-1-肠激酶位点接头人igg1fc
seqidno:28:具有“臼”突变y349c/t366s/l368a/y407v的人igg1fc
seqidno:29:具有“杵”突变(s354c/t366w)-接头-肠激酶位点-nc-1的人igg1fc
seqidno:30:具有“臼”突变y349c/t366s/l368a/y407v的人igg1fc
seqidno:31:具有“杵”突变s354c/t366w的nc-1-fc融合蛋白;异源二聚体nc-1-fc(分子#1)的链1
seqidno:32:具有“臼”突变y349c/t366s/l368a/y407v的fc;异源二聚nc-1-fc(分子#1)的链2
seqidno:33:具有“杵”突变s354c/t366w的nc-1-fc融合蛋白;同源二聚体nc-1-fc(分子#2)的链1
seqidno:34:具有“臼”突变y349c/t366s/l368a/y407v的fc;同源二聚nc-1-fc(分子#2)的链2
seqidno:35:具有“杵”突变s354c/t366w的fc-内皮抑制素融合蛋白;异源二聚体fc-内皮抑制素(分子#3)的链1
seqidno:36:具有“臼”突变y349c/t366s/l368a/y407v的fc-内皮抑制素融合蛋白;异源二聚体fc-内皮抑制素(分子#3)的链2
seqidno:37:单体nc-1-fc,其中fc包括单体突变f405r和yte半衰期延长突变m252y/s254t/t256e(分子#4)
seqidno:38:单体fc-内皮抑制素,其中fc包括单体突变f405r和yte半衰期延长突变m252y/s254t/t256e(分子#5)
附图说明
这些图显示:
图1:来自细胞培养上清液的小鼠nc-1和人nc-1的sds-page(左)和蛋白质印迹(右)分析。
m泳道:标志物
第1-3泳道:在还原条件下,转染后第2、4和5天的细胞培养上清液
第4-6泳道:在非还原条件下,转染后第2、4和5天的细胞培养上清液
nc1泳道:还原条件下的阴性对照
nc2泳道:非还原条件下的阴性对照
一级抗体:小鼠抗hismab(genscript)
图2:小鼠nc-1和人nc-1的细胞裂解物的sds-page(左)和蛋白质印迹(右)分析。
第1-3泳道:在还原条件下,转染后第2、4和5天的细胞裂解液
第4泳道:在还原条件下,转染后第4天的细胞碎片
第5-7泳道:在非还原条件下,转染后第2、4和5天的细胞裂解液
第8泳道:在非还原条件下,转染后第4天的细胞碎片
nc1泳道:还原条件下的阴性对照
nc2泳道:非还原条件下的阴性对照
一级抗体:小鼠抗hismab(genscript)
图3:使用两个编码nc-1-fc-杵和fc-臼的质粒的异源二聚nc-1-fc的瞬时哺乳动物(expi)表达导致蛋白a纯化后在sec中形成两个峰。每个峰的组分在还原和非还原条件下通过sds-page显示。峰1由还原条件下的两个带组成,分别包含在62kda的nc-1-fc-杵和在30kda的fc-臼。相比之下,峰2仅由在还原条件下单个fc臼带组成,表明生成了fc-臼二聚体。因此,峰1对应于非还原条件下的88kda异源二聚体nc-1-fc(kih)蛋白,而fc-臼二聚体峰2的大小对应于非还原条件下的约50kda蛋白。
图4:蛋白上清液的蛋白a纯化后,进行sec-hplc以检测天然条件下表达的蛋白的大小。为了更好地估计表达的蛋白质(两个峰)的大小,使用了由igg1/2b,白蛋白,人fc和igg1-fab组成的对照中众所周知的波峰(spike)。如所预期的,由fc-臼二聚体组成的峰2与fc结构域同时运行。但是,峰1似乎比标准igg(150kda)更大。这是出乎意料的,因为图3中还原的凝胶清楚地表明峰1同时包含nc-1-fc-杵和fc-臼部分,从而产生了88kda的异二聚nc-1-fc蛋白。
图5:以比例为2:1的nc-1-fc-杵vs.fc-臼质粒转导哺乳动物细胞增强了异源二聚体nc-1-fc-kih的形成,并几乎减少了fc-臼二聚体的形成。
图6:交联研究最终证实了sec的观察结果,即异源二聚体nc-1-fc必须比非还原的凝胶(带2)中发现的88kda单体分子大得多。事实上,异源二聚体nc-1-fc(kih)在生理条件下形成三聚体(带3)。交联程序的详细信息可见图17的描述。
图7:nc-1-fc-kih在lewis肺癌模型中的优异抗癌作用。同基因lewis肺癌(llc)模型在c57bl6小鼠中的肿瘤生长动力学。与所有重组单体内皮抑制素分子以及在fc部分上形成二聚体的fc-内皮抑制素(fc-es)相比,三聚体nc-1-fc(kih)展示更高的活性。使用了三个单体内皮抑制素化合物,包括异源二聚体fc(kih)内皮抑制素(fc-(kih)-es),其与二聚体fc形成单体内皮抑制素,即异源二聚体nc-1-fc(kih)的参考骨架构建体,恩度(在中国获得批准的单体重组大肠杆菌his标签内皮抑制素),和entemed内皮抑制素(es-entremed)(在美国/欧洲进行i/ii期试验的单体重组巴斯德毕赤酵母内皮抑制素)。所有药物均经皮下注射内皮抑制素等同剂量为20μg/小鼠/天。条形表示n:7+/-sem的平均值。
图8:肿瘤生长抑制。在治疗开始后第9天,三聚体nc-1-fc(kih)表现最佳,对llc肿瘤的生长具有51%的抑制作用(b)。与所有单体内皮抑制素分子相比,这种作用显著地更为明显(p<0.001)。有趣的是,nc-1-fc(kih)比二聚体fc-es更有效(a,p<0.01)。因此,除了寡聚化之外,整个nc-1序列vs.仅n-端内皮抑制素结构域的其他生物学特性可能与该分子的抗癌活性有关。
图9:异源二聚体nc-1-fc(分子#1)
图10:单体nc-1-fc(分子#4)
图11:同源二聚体nc-1-fc(分子#2)
图12:异源二聚体fc-内皮抑制素(分子#3)
图13:单体fc-内皮抑制素(分子#5)
图14:类似于fc-内皮抑制素(分子#6),使用igg1野生型骨架的n和c-端nc-1-fc融合蛋白
图15:(a)异源二聚体蛋白,其由融合至不同部分(包括iggfab片段)的nc-1-fc(杵)和fc(臼)组成(分子#7)。(b)
图16:在肺纤维化模型中用包含单体nc-1-fc(nc-1-fc(单体))(seqidno:37)的蛋白寡聚体的体内治疗(参见实施例4)。
纤维化数据每组n:10
ir剂量:15gy
单体:图案结构
*p<0.05,**p<0.01
ir(红色)vs.未辐照(黑色)线后16周的虚线。
图17:nc-1-fc-单体形成三聚体
即使在非还原条件下,通过凝胶电泳,蛋白质缔合和3d特性也会部分丢失。因此,需要天然蛋白质的交联来破译nc-1-fc-mono的组成。为此,利用按照(kuo,javaherianetal.jcb2001,pmid:11257123)描述的交联方案,加一些改变。简而言之,在50μl含有50μm氯化锌的pbs中使用50μg蛋白质。向其中加入以10mm浓度溶解于dmso中的5μl交联剂egs(乙二醇-双(琥珀酸n-羟基琥珀酰亚胺酯)。将样品在37℃下温育1/2小时后,用4μl的1mtris,ph8终止反应。最后,将样品在非还原条件下进行sds-page。发明人的数据清楚地表明,如在泳道3中交联后发现的,nc-1-fc(单体)在nc-1寡聚结构域上形成了三聚体。这些数据与在sec中天然条件下对nc-1-fc(单体)大小的观察相符。
表格
表1:异源二聚体nc-1-fc(分子#1)
表2:同源nc-1-fc(分子#2)
表3:异源二聚体fc-内皮抑制素(分子#3)
表4:单体nc-1-fc(分子#4)
表5:单体fc-内皮抑制素(分子#5)
表6:设计但失败了的其他构建体
现在将通过实施例说明本发明,但是,这些实施例不应解释为限制本发明的范围。
实施例
实施例1:nc-1的重组表达在临床前和临床研究中不可行
胶原蛋白18nc-1的生产从一开始就是一个巨大的挑战,这是由于该分子在寡聚化结构域上的聚集。因此,没有关于nc-1功效的体内数据,并且对迄今为止被认为是内皮抑制素的前体分子仅进行了很少的研究。
javaherian博士和其他少数实验室能分别通过his标签和flag标签表达很少的重组nc-1,但对于动物的临床前给药而言,这还远远不够。因此,与临床前和临床研究相关的nc-1量的产生构成了挑战。在证实该假设后,本发明人以下提供关于人和小鼠his标签nc-1表达的超低产量的数据。
设计并合成了编码小鼠nc-1和人nc-1的dna序列。然后将完整序列,即ecori-kozak序列-前导序列-his6标签-小鼠nc-1-终止密码子-hindiii(seqidno.40),以及ecori-kozak序列-前导序列-his6标签-人nc-1-终止密码子-hindiii(seqidno.42)亚克隆到pcdna3.4载体中用于expi293f细胞表达。expi293f细胞在无血清expi293ftm表达培养基(thermofisherscientific)中生长。将细胞在定轨摇床(vwrscientific,chester,pa)上在37℃,8%co2下保持在锥形瓶(corninginc.,acton,ma)中。转染前一天,将细胞以适当的密度接种在康宁锥形瓶中。转染当天,将dna和转染试剂以最佳比例混合,然后将其与准备用于转染的细胞一起加入锥形瓶中。将编码小鼠nc-1和人nc-1靶蛋白的重组质粒分别瞬时转染到40ml悬浮expi293f细胞培养物中。在第2、4和5天收集的细胞培养上清液用于蛋白质表达评价。转染后第2天、第4天和第5天收集约1ml细胞培养上清液和细胞沉淀样品,并通过sds-page和蛋白印迹分析法进行分析,以评价小鼠nc-1和人nc-1的表达水平,如图1和2所示。用于蛋白质印迹的一级抗体是小鼠-抗-hismab(genscript,cat.no.a00186)。
本发明人已经试图在悬浮的expi293f细胞培养物中表达和检测小鼠nc-1和人nc-1。
对于小鼠nc-1,通过蛋白质印迹分析从细胞培养上清液中阳性地检测到弱的目的条带,如图1b所示。通过还原条件sds-page和蛋白质印迹分析,从细胞碎片中观察到具有约为40kda(cal.mw.38.17)的估计的分子量的目的信号,如图2e和f所示(黑色箭头)。这表明靶蛋白的分泌不足。靶蛋白的估计表达水平为~0.1mg/l。这表明靶蛋白的分泌不足。
对于人类nc-1,在还原条件下,通过sds-page和蛋白质印迹分析,从细胞培养上清液和细胞碎片中检测到明显的估计分子量约为40kda(ccal.mw.37.47)的目的条带,如图1c和d和图2g和h所示。靶蛋白的估计表达水平低<3mg/l。与小鼠nc-1相比,人nc-1的表达更好,但在大多数下游评价(例如体内研究)中仍处于较低水平。
该数据表明nc-1的重组表达具有挑战性并且对于临床前和临床研究是不可行的。
实施例2:改造胶原蛋白18nc-1-fc构建体
本发明的主要目的是在免疫球蛋白iggfc缀合领域中前进,取得众多生物技术、药代动力学和生物学优势。
然而,从以下事实可以明显看出,fc-nc-1的产生不是直接的过程,发明人迄今已经花费了近十年的时间来实现用于大规模生产nc-1的系统,并且许多构建体和经典标签都失败了。
其原因是存在两种寡聚化力,第一种是由nc-1寡聚化结构域诱导的三聚化力,第二种是由igg-fc诱导的二聚化力。发明人已经花费了大量的时间用所有不同的现有技术,即,nc-1的c-或n-端与fc的缀合,如先前用内皮抑制素所进行的,而所有这些方法均失败了,如例如表2和6。
不得不考虑的是,在经典的fc标记方法中,将产生n-端或c-端缀合的fc-nc-1分子,如果没有nc-1寡聚结构域,该分子将形成二聚体。但是,具有nc-1寡聚结构域,人们则收到聚集体和非常纯净的表达。
就本发明人所知,没有关于成功的nc-1-fc缀合的文献。
如下所述,本发明人已经研究了规避该主要问题的不同方法,并且经过十多年的研究发现了成功的策略。一个核心点是生成单价nc-1-二聚体fc或异二聚nc-1-fc。发明人决定使用在fc中的突变,该突变使用fc“杵臼结构”(kih)策略阻止nc-1-fc与nc-1-fc的二聚化。因此,nc-1-fc杵只会与空的fc-臼二聚化。尽管禁止杵-杵二聚化,但仍可能发生臼-臼二聚化。实际上,本发明人发现了两个峰,一个峰是预期的nc-1-fc-kih异源二聚体,第二个峰是fc-臼-fc-臼二聚体。这表明nc-1-fc-杵对于细胞来说更难以表达。发明人通过增加nc-1-fc-杵对fc-臼(以2:1的起始比例,然后是4:1)的表达再次规避了该问题,这主要导致异源二聚的nc-1-fc-kih的一个峰具有出色的表达功效。然而,在尺寸排阻柱中,本发明人认识到在生理条件下的分子比根据nc-1-fc-kih异源二聚体所期望的分子大得多。随后的交联实验证实,他们已经惊人地实现了生产三聚体分子,即nc-1-fc-kih异源二聚体的三聚体。该分子保留了fc加三聚体nc-1的所有优异特性。令人惊讶的是,这种三聚体nc-1-fc-kih分子在结合独特的寡聚nc-1结合伴侣(如纤连蛋白,vegf和mmp-2/-9)方面展示出相同的功效。凭借优异的表达率,本发明人在很长一段时间后现在已经可以使用该nc-1分子在不同模型中进行体内实验。除肺癌(llc)模型和肺纤维化模型中的数据外,正在进行进一步的实验,这将极大地提高了他们对该分子的理解。
值得注意的是,nc-1-fc-杵和nc-1-fc-臼的分别表达以及随后通过氧化还原系统的二聚化均未能产生有效的nc-1-fc构建体。因此,异源二聚体fc的空间位阻对于聚集的成功和缺乏至关重要。接下来,本发明人旨在评价向“空”fc-臼中添加部分是否会影响该分子。为此,他们将抗体的经典fab片段缀合到fc-臼,并与nc-1-fc-杵共表达。有趣的是,与nc-1-fc-kih异源二聚体相比,通过fab部分nc-1-fc-kih-fab添加空间位阻降低了表达效率。
总之,三聚体nc-1-fc-kih异源二聚体(seqidno:31/32;表1;图9;分子#1)似乎是本发明人报道的第一个基于nc-1的fc构建体,并且考虑到nc-1和fc的3d复杂性,改造这种分子过去不是,而且现在也不是一个可以从当前文献中推论出来的不重要的过程。
实施例2a:异源二聚体nc-1-fc(seqidno:31/32)
使用两个编码nc-1-fc-杵和fc-臼的质粒的异源二聚nc-1-fc-kih的瞬时哺乳动物(expi)表达导致蛋白a纯化后在sec中形成两个峰。每个峰的组分通过在还原和非还原条件下的sds-page显示在图3中。峰1由还原条件下的两个带组成,分别包含在62kda的nc-1-fc-杵和在30kda的fc-臼。相比之下,峰2在还原条件下仅由单个fc臼带组成,表明生成了fc臼二聚体。因此,峰1对应于非还原条件下的88kda异源二聚体nc-1-fc-kih蛋白,而fc-臼二聚体峰2的大小对应于非还原条件下的约50kda蛋白,见图3。
本发明人针对这两个分离的峰运行了sec-hplc,并将它们与具有已知大小的其他蛋白质进行了比较(参见图4至6)。如所预期的,峰2运行的大小与fc结构域相同。峰1似乎比标准igg更大。这是出乎意料的,因为图3中还原凝胶清楚地表明峰1既包含杵部分又包含臼部分(即62kda处的nc-1-fc-杵和30kda处的fc-臼)。
交联研究最终证实,本发明人首次能够在fc骨架的顶部生成nc-1三聚体,即nc-1-fc-kih(或nc-1-fc(kih))在生理条件下的三聚体。
通过增加nc-1-fc-杵vs.fc-臼的量(以2:1的起始比例,然后是4:1),他们能够通过几乎消除臼-臼形成来进一步优化此nc-1-fc-kih三聚体构建体的生产(峰2);参见图5。
迄今为止的结合研究证实了,nc-1-fc-kih三聚体保留了nc-1的中心特性,像与纤连蛋白以及vegf和mmp-2/-9的结合。
交联实验证实了在生理条件下三聚体nc-1-fc-kih异源二聚体的形成,参见图6。
实施例2b:单体nc-1-fc(seqidno:37)
本发明人还生成了单体nc-1-fc(seqidno:37;图10;分子#4)。igg1fc包括单体突变f405r和yte半衰期延长突变m252y/s254t/t256e。表达时可获得高产量的单体nc-1-fc(在本文中也称为nc-1-fc(单体))。如在图17的泳道3中交联后发现的,nc-1-fc(单体)在nc-1寡聚结构域上形成三聚体;另请参见实施例5。如实施例4所证实的,包含单体nc-1-fc(nc-1-fc(单体))(seqidno:37)的蛋白寡聚体在肺纤维化模型中显示出优异的抗纤维化活性。
实施例3:胶原蛋白18nc-1-fc构建体的抗癌作用
作为肿瘤模型,已经使用了在c57bl6小鼠背景下的lewis肺癌(llc)皮下同基因模型。小鼠已用以下的等同剂量治疗:
-在nc-1寡聚结构域(nc-1-fc(kih))上形成三聚体的异源二聚体nc-1-fc(kih)(seqidno:31/32),
-异源二聚体fc(kih)内皮抑制素(fc-(kih)-es)(seqidno:35/36),其与二聚体fc形成单体内皮抑制素,即异源二聚体nc-1-fc(kih)的参考构建体,
-在fc部分上形成二聚体的fc-内皮抑制素(fc-es),
-恩度,其是在中国获批的单体重组大肠杆菌his标签内皮抑制素,
-entemed内皮抑制素(es-entremed),一种在美国/欧洲进行i/ii期试验的单体重组巴斯德毕赤酵母内皮抑制素。
肿瘤生长延迟数据清楚地表明,基于nc-1的药物设计,三聚体nc-1-fc(kih)在原型llc模型中在直接背对背比较中在肿瘤生长抑制方面优于所有其他构建体;参见图7。
值得注意的是,相比二聚体fc-es的抑制作用为30%(p<0.01),三聚体nc-1-fc(kih)在治疗开始后第9天对肿瘤生长的抑制作用为51%的优越的活性清楚地表明,整个nc-1序列vs.仅n-端内皮抑制素结构域的不仅寡聚化(三聚体vs.二聚体)而且其他生物学特性都有助于三聚体nc-1-fc(kih)的治疗效果;参见图8。
实施例4:胶原蛋白18nc-1-fc构建体的抗纤维化作用
在体内治疗中,已经在肺纤维化模型中生成了包含单体nc-1-fc(nc-1-fc(单体))(seqidno:37)的蛋白寡聚体,其结果显示在图16中。
本发明人先前已经表明,通过nc-1模拟(寡聚fc-内皮抑制素,fces)证实的内皮抑制素的n-端和寡聚化对于防止由电离辐射诱导的肺纤维化的发展是至关重要的(ir;wo2017/093569)。在与fce和三种单体内皮抑制素构建体(即entremed内皮抑制素、恩度和fc(单体)-es)进行背靠背比较中,本发明人旨在发现包含重组单体nc-1-fc(nc-1-fc(单体))(seqidno:37)的新型蛋白寡聚体以抑制并最终逆转纤维化的潜力。使用了c57b16辐射诱导的肺纤维化(rilf)模型。在进行15gy整个胸腔照射后16周开始治疗,其中平均肺密度从对照小鼠的中-380hounsfield单位(hu,黑色虚线)增加到辐照肺部的-320hu(红色虚线)(图16a)。所有化合物均经皮下每天以等摩尔剂量持续施用8周。寡聚构建体fces和nc-1-fc(单体)均通过降低肺密度(图16b),增加肺体积(图16c)和改善基于ct的定量肺纤维化指数(图16d)显示抗纤维化作用)(fi,zhouetal.2017,16;https://www.ncbi.nlm.nih.gov/pubmed/29116014)。
有趣的是,在该干预试验中,包含重组单体nc-1-fc(nc-1-fc(单体))(seqidno:37)的新型蛋白寡聚体是唯一能够逆转纤维化的化合物。与fces相比,本发明的所述蛋白寡聚体的抗纤维化作用明显更强(p<0.01)。所有三个单体构建体均显示为图案条形。条形表示n:10+/-sem的平均值。
实施例5:nc-1-fc(单体)(seqidno.37)形成三聚体
即使在非还原条件下,通过凝胶电泳,蛋白质缔合和3d特性也会部分丢失。因此,需要天然蛋白质的交联来破译nc-1-fc-单体的组成。为此,利用按照(kuo,javaherianetal.jcb2001,pmid:11257123)描述的交联方案,加一些改变。简而言之,在50μl含有50μm氯化锌的pbs中使用50μg蛋白质。向其中加入以10mm浓度溶解于dmso中的5μl交联剂egs(乙二醇-双(琥珀酸n-羟基琥珀酰亚胺酯)。将样品在37℃下温育1/2小时后,用4μl的1mtris,ph8终止反应。最后,将样品在非还原条件下进行sds-page。发明人的数据清楚地表明,如在图17的泳道3中交联后发现的,nc-1-fc(单体)在nc-1寡聚结构域上形成了三聚体。这些数据与在sec中天然条件下对nc-1-fc(单体)大小的观察相符。
表1:异源二聚体nc-1-fc
表2:同源二聚体nc-1-fc
表3:异源二聚体fc-内皮抑制素
表4:单体nc-1-fc
表5:单体fc-内皮抑素
图6:失败的额外构建体
起点商标作为专业知识产权交易平台,可以帮助大家解决很多问题,如果大家想要了解更多知产交易信息请点击 【在线咨询】或添加微信 【19522093243】 与客服一对一沟通,为大家解决相关问题。
与客服一对一沟通,为大家解决相关问题。
此文章来源于网络,如有侵权,请联系删除

 热门咨询
热门咨询



