
 商标分类
商标分类  商标转让
商标转让 
油酸衍生物的选择性水合的制作方法
 2021-02-02 17:02:56|
2021-02-02 17:02:56| 474|
474| 起点商标网
起点商标网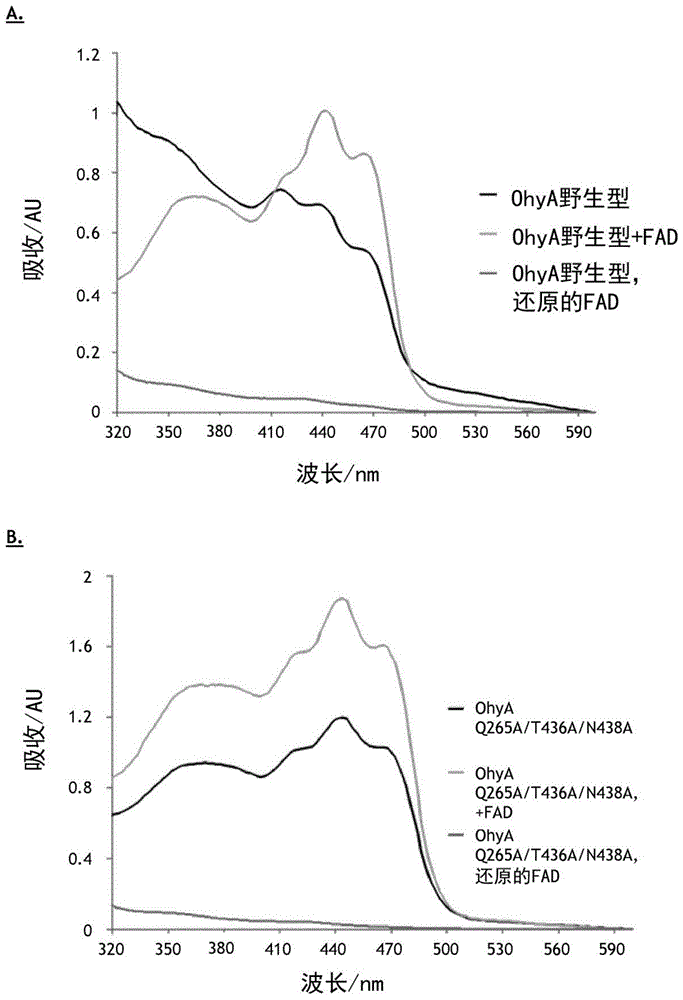
本发明涉及通过经修饰的油酸水合酶的作用对包括油酸酯的油酸衍生物进行区域选择性和立体选择性水合。脂肪酸水合酶(fahy;ec4.2.1.-)通过将水加成到游离的单不饱和或多不饱和脂肪酸的分离的碳-碳双键来催化中链和长链羟基脂肪酸的形成。因此,它们提供了仲醇和叔醇的取得方法,这使得它们成为了用于生产各种化学品(包括风味添加剂、化妆品、表面活性剂、润滑剂和聚合物化学品中的前体)的有价值工具。fahy在合成中的使用有望带来与它们的出色的区域选择性和立体选择性有关的明显优势,所述区域选择性和立体选择性允许使用非选择性的酸催化化学水合不可能发生的反应。多数已知的fahy在对不饱和脂肪酸的顺式-9或顺式-12双键进行水合中为高度区域选择性的。但尚没有关于通过任何已知的fahy进行脂肪酸酯的水合的任何已知信息。迄今为止,最充分表征的fahy是来自脑膜炎败血伊丽莎白菌(elizabethkingiameningoseptica)的油酸水合酶(ohya,ec4.2.1.53)。它催化油酸(oa)的区域选择性和立体选择性水合,从而在不需要辅因子再循环的情况下以至少98%的卓越对映体过量(enantiomericexcess,ee)产生(r)-10-羟基硬脂酸。到目前为止,fahy在工业环境中的适用性仍然受到限制,这可主要归因于其狭窄的底物范围:所有工作表明,羧基、顺式构型中的双键、羧酸根与顺式双键之间7个碳的最小距离和11个碳的最小脂肪酸链长度对于转换而言是必不可少的。迄今为止尚未有人描述在末端碳处的水合,因为在伯碳处形成部分正电荷将与所提出的反应机理相冲突。因此,需要拓宽作为迄今为止所表征的最佳酶的fahy,具体是是ohya的底物谱,以便将它用作工业过程中的工具。然而,卓越的区域选择性和立体选择性不应受到影响。令人惊讶地,我们现在发现ohya对油酸衍生物的底物耐受性是由对位于活性侧空腔(底物结合区)中的某些氨基酸,即高度保守的氨基酸残基的修饰触发的(根据水合酶工程改造数据库hyed中的序列比对;参见表1)。具体地,本发明涉及具有fahy(ec4.2.1.-)的活性,例如ohya(ec4.2.1.53)的活性的经修饰的酶,所述酶以至少98%的ee催化水合反应,其中所述经修饰的酶在与根据seqidno:1的多肽中的选自由265位、436位、438位、442位以及它们的组合组成的组的残基对应的位置处包含一个或多个氨基酸取代。显示出ohya活性的根据seqidno:1的多肽,包括由根据seqidno:2的多核苷酸编码的多肽,已从脑膜炎败血伊丽莎白菌中分离出来(序列来源于genbank登录号act54545.1)。已从不同来源,包括哺乳动物、酵母或植物、或细菌分离出了fahy。如本文所用,与未修饰的酶相比,“经修饰的”酶,即经修饰的fahy,特别是经修饰的ohya,具有朝向油酸衍生物的区域选择性和立体选择性水合的优选活性和/或特异性。如本文所用,“未修饰的”fahy,特别是未修饰的ohya是指不携带如本文所定义的一个或多个氨基酸取代的相应酶,在本文中也称为野生型酶。如本文所用,携带如本文所定义的经修饰的fahy活性,特别是包含如本文所定义的一个或多个氨基酸取代的ohya的宿主细胞被称为“经修饰的”宿主细胞。携带未修饰的酶活性,即编码野生型ohya基因的相应宿主细胞被称为“未修饰的”宿主细胞。在一个实施方式中,如本文所定义的经修饰的酶,特别是经修饰的ohya活性,包含在与根据seqidno:1的多肽中的残基265对应的位置处的氨基酸取代,优选地由丙氨酸取代谷氨酰胺(q265a)。优选地,具有经修饰的ohya活性的酶源自脑膜炎败血伊丽莎白菌。所述突变可以与如本文所定义的1个、2个、3个或更多个突变组合。在一个实施方式中,如本文所定义的经修饰的酶,特别是经修饰的ohya活性,包含在与根据seqidno:1的多肽中的残基436对应的位置处的氨基酸取代,优选地由丙氨酸取代天冬酰胺(t436a)。优选地,具有经修饰的ohya活性的酶源自脑膜炎败血伊丽莎白菌。所述突变可以与如本文所定义的1个、2个、3个或更多个突变组合。在另一个实施方式中,如本文所定义的经修饰的酶,特别是经修饰的ohya活性,包含在与根据seqidno:1的多肽中的残基438对应的位置处的氨基酸取代,优选地由丙氨酸取代天冬酰胺(n438a)。优选地,具有经修饰的ohya活性的酶源自脑膜炎败血伊丽莎白菌。所述突变可以与如本文所定义的1个、2个、3个或更多个突变组合。在另一个实施方式中,如本文所定义的经修饰的酶,特别是经修饰的ohya活性,包含在与根据seqidno:1的多肽中的残基442对应的位置处的氨基酸取代,优选地由丙氨酸取代组氨酸(h442a)。优选地,具有经修饰的ohya活性的酶源自脑膜炎败血伊丽莎白菌。所述突变可以与如本文所定义的1个、2个、3个或更多个突变组合。优选地,可以将与seqidno:1中的残基q265a对应的位置处的氨基酸取代与其他取代(例如在与seqidno:1中的t436a和/或seqidno:1中的n438a和/或seqidno:1中的h442a对应的一个或多个位置处的氨基酸取代)进行组合。优选的经修饰的酶为这样的酶,所述酶具有ohya活性并且至少包含在与seqidno:1中的q265a、t436a、n438a对应的位置处的氨基酸取代,显示出相应油酸衍生物转换的至少2倍增加(参见图3和图4)。当使用异羟肟酸(hydroxamicacid)作为底物时,通过在生物转换中应用经修饰的ohya变体q265a/n438a和ohyaq265a/t436a/n438a,可以实现至少5倍增强的相对水合活性。在使用油醇,即底物(5)时,使用ohyaq265a/t436a/n438a的水合为与使用野生型ohya相比的至少2倍高。当使用三重突变时,对底物(6)油酸甲酯和底物(7)油酸乙酯的相对活性为6倍高,并且对底物(9)油酸正丙酯的相对活性为甚至20倍高。如本文所用,ohya的活性被修饰。这可以通过以下方式来实现:例如将一个或多个突变引入编码ohya的基因中,即,如本文所述的一个或多个位置上的氨基酸取代。技术人员知道如何遗传操纵细胞,从而导致对ohya活性的修饰。这些遗传操纵包括但不限于例如使用质粒、病毒或其他载体进行基因置换、基因扩增、基因破坏、转染、转化。可以以不同的方式来执行核酸或氨基酸中的突变产生,即诱变,诸如通过随机诱变或定点诱变,由诸如辐射等试剂引起物理损伤、化学处理,或插入遗传元件。技术人员知道如何引入突变。本发明特别地涉及如本文所定义的此类经修饰的ohya酶在用于转换非天然油酸衍生物的方法中的用途。优选地,将本发明的经修饰的酶引入和/或在合适的宿主细胞(例如大肠杆菌(e.coli))中表达,即表达为重组或异源酶。转换反应可与来自含有如本文所定义的重组表达的经修饰的ohya的大肠杆菌的无细胞提取物(cell-freeextract,cfe)或大肠杆菌全细胞一起在生物转化反应中使用。优选地,如本文所定义的底物转换是在生物转化反应中,例如使用带有重组ohya的大肠杆菌细胞在96小时生物转化后执行的。来自生物转化的产物可以纯化并通过nmr分析验证。利用如本文所述的经修饰的酶,各种非天然油酸衍生物可被水合,从而导致至少98%,例如99%或甚至100%的ee。在一个方面中,本发明涉及一种用于使用如本文所定义的经修饰的酶来转换非天然脂肪酸衍生物的方法,即缺乏游离羧酸根头基(headgroup)的脂肪酸的水合反应,其中实现了至少98%的ee。优选地,该方法独立于任何已知的共底物(co-substrate)或辅因子,例如氧化或还原的fad或二硫苏糖醇(dtt)。优选地,该方法在全细胞生物转化中进行至少22小时。如本文所用,术语“油酸衍生物”是指用作本文所述的ohya酶的底物的缺乏游离羧酸根头基的非天然脂肪酸衍生物。它包括但不限于短链油酸酯,例如油酸甲酯、油酸乙酯、油酸异丙酯、油酸正丙酯、油酸正丁酯或酰胺(例如油酰胺或n-oh油酰胺)、异羟肟酸或醇(例如油醇)。优选地,它包括如本文所定义的底物(1)至(10),例如油酸(1)、油胺(2)、油酰胺(3)、n-oh油酰胺(4)、油醇(5)、oa甲酯(6)、oa乙酯(7)、oa异丙酯(8)、oa正丙酯(9)、或oa正丁酯。如本文所用,关于酶的术语“比活性”或“活性”是指其催化活性,即其催化从给定底物形成产物的能力。比活性定义了在给定时间段内和在限定温度下每限定量的蛋白质消耗的底物和/或产生的产物的量。通常,比活性表示为每mg蛋白质每分钟消耗的底物或形成的产物的μmol数。通常,μmol/min缩写为u(=单位)。因此,在本文件全文中可互换地使用μmol/min/(mg蛋白质)或u/(mg蛋白质)的比活性单位定义。如果酶在体内(即在如本文所限定的宿主细胞内)或在合适的底物存在下的合适的(无细胞)系统内执行其催化活性,则所述酶是有活性的。技术人员知道如何测量酶活性,例如通过hplc。关于本发明,应当理解的是,生物(诸如微生物、真菌、藻类或植物)也包括具有相同生理性质的此类物种的同义词或基名(basonyms),如由国际原核生物命名法(internationalcodeofnomenclatureofprokaryotes)或关于藻类、真菌和植物的国际命名法(internationalcodeofnomenclature)(melbourne法)所限定的。附图说明图1.纯化的ohya在黄素蛋白重构之前(黑色曲线)和重构之后(橙色曲线)以及在用dtt还原fad辅因子之后(蓝色曲线)的uv-vis(紫外-可见光)吸收光谱。野生型ohya的uv-vis吸收光谱(图1a)。突变型ohyaq265a/t436a/n438a的uv-vis吸收光谱(图1b)。有关更多说明,请参见文本。图2.由作为全细胞大肠杆菌生物催化剂的ohya野生型(wt)和氨基酸交换变体在酶过表达后对油酰胺(图2a)、n-羟基油酰胺(图2b)、油醇(图2c)、oa甲酯(图2d)、oa乙酯(图2e)、oa异丙酯(图2f)、oa正丙酯(图2g)、oa正丁酯(图2h)的转换。对照反应含有添加到不含细胞的反应缓冲液中的底物或添加到大肠杆菌空载体对照(emptyvectorcontrol,evc)中的底物。图3.由脑膜炎败血伊丽莎白菌突变油酸水合酶(ohya)对油酸(oa)(即底物(1))和oa衍生物(即底物(2)至(10))的区域选择性和立体选择性水合。在生物转化测定中使用具有过度表达的水合酶的全细胞大肠杆菌生物催化剂。有关更多说明,请参见文本。图4.ohya野生型(wt)和ohyaq265a/t436a/n438a对底物(1)油酸和非天然油酸衍生的底物(2)至(10)的转换。在酶表达时,使用全细胞大肠杆菌生物催化剂进行反应22小时。以下实施例仅是说明性的,并不意图以任何方式限制本发明的范围。实施例实施例1:通用方法、菌株和质粒除非另有说明,否则标准实验室试剂以可得的最高纯度获自(steinheim,germany)或carlrothgmbh&co.kg(karlsruhe,germany)。油酸(oa)及其酯(甲酯、乙酯、异丙酯)、油醇和油胺购自(steinheim,germany)。在使用前将oa甲酯和oa乙酯以及油醇蒸馏至根据gc-fid分析纯度>90%。根据标准工序(ausubel等人,currentprotocolsinmolecularbiology,2003)来进行表达载体的分子克隆,并且通过测序(lgcgenomics,berlin,germany)确认插入物的正确整合。使用高保真dna聚合酶(thermofisherscientificinc.,st.leon-rot,germany)根据推荐的pcr方案进行基因扩增。ohya的密码子优化基因变体(脑膜炎败血伊丽莎白菌xp_001209325油酸水合酶)购自dna2.0(menlopark,ca)。为了表达重组ohya,构建经修饰的pms470表达载体pms470-histev-ohya。对于所有克隆步骤和质粒复制,使用来自lifetechnologies(vienna,austria)的大肠杆菌top10f'(f'[laciqtn10(tetr)]mcraδ(mrr-hsdrms-mcrbc)δlacx74deornupgreca1arad139δ(ara-leu)7697galugalkrpsl(strr)enda1λ-)。重组ohya在大肠杆菌bl21startm(de3)(f-ompthsdsb(rb-mb-)galdcmrne131(de3))(lifetechnologies,vienna,austria)中表达。将ohya的蛋白质序列与水合酶工程改造数据库(hyed)中的氨基酸序列进行了比较,在所述hyed中收集了总共2046个序列。由于ohya被归类于hyed的同源家族11(hfam11)中,因此选择来自hfam11的所有氨基酸序列进行多重序列比对。如sievers等人(mol.syst.biol.7,539,2011)中所述以clustalomega序列比对工具使用默认设置从数据库中提取序列进行多重序列比对,并使用ugene软件进行可视化。表1中列出了用于构建ohya突变体的高度保守残基的位置。表1:在水合酶工程改造数据库(hyed)中收集的来自hfam11的氨基酸序列的多重序列比对,突出显示了参与羧酸根结合的保守残基。应注意,h442在除一个成员外的所有成员中均为保守的,在所述一个成员中h442被用q取代。有关更多说明,请参见文本。为了构建ohya突变体,根据制造商的手册(stratagenequikchangetm定点诱变方案)使用以下引物。分别制备25μl的两个单独的含有正向引物和反向引物的pcr反应(表2)。在5个循环步骤后,合并pcr反应,并根据手册继续将pcr进行20个附加循环。通过对构建体的编码区进行dna测序来验证突变的质粒。表2:用于构建ohya突变体的引物。粗体字母表示突变的密码子。为了纯化重组的ohya,将细胞沉淀重悬于含有10mm咪唑的50mmhepes,ph7.4中。使用250(branson,danbury,ct)超声处理细胞4分钟,将占空比设置为80%并且将输出控件设置为8级来裂解细胞。通过在48300xg和4℃下离心35分钟将无细胞提取物(cfe)与总细胞裂解物(tcl)分离,并且滤过0.22μm过滤器(millipore,bedford,ma),然后将其上样到预平衡的自填充ni-nta亲和色谱柱(gehealthcare,unitedkingdom)上。在进行任何进一步分析之前,将纯化的ohya与10倍摩尔过量的fad在4℃下孵育过夜。根据推荐的方案,通过经由pd-10脱盐柱(gehealthcare,unitedkingdom)进行缓冲液交换来从蛋白质等分试样中去除未结合的fad。野生型ohya和突变型ohyaq265a/t436a/n438a的紫外-可见光(uv-vis)吸收光谱是在specord205双光束分光光度计(analytikjenaag,germany)上使用光程长度为1cm的石英比色皿在250nm至1000nm的光谱范围内记录的。在含有50mmnacl的50mmhepes,ph7.4中进行光谱测量。通过以下方式确定fad与纯化的ohya的结合:基于负载fad的酶的为111115m-1cm-1的和与ohya结构非共价连接的fad的先前测量为8074m-1cm-1的来计算蛋白质浓度。通过气相色谱-质谱(gc-ms)鉴定和分析游离脂肪酸,并将所得的质谱裂解图谱与真实标准品进行比较。使用在配备有质量选择性检测器的hewlett-packard6890系列iigc上的hp-5柱(经交联的5%ph-me硅氧烷;长30m,直径0.25mm并且膜厚0.25μm)。在恒定流量模式下用n2作为载体以设置为36cms-1的流率将1μl的样品等分试样在240℃进样器温度和290℃检测器温度下以分流模式(分流比30:1)进样。温度程序如下:100℃持续1分钟,以15℃min-1升温至300℃,保持5分钟。总运行时间为19.33分钟。质量选择性检测器在1765v的电子倍增器电压下,在50-400amu的质量范围内操作。使用gc-ms数据分析软件(agilenttechnologies,austria)评估结果。实施例2:在大肠杆菌中表达的ohya突变体的全细胞生物转化使ohya在大肠杆菌中重组表达。首先,用带有pms470-histev-ohya野生型酶或其变体的大肠杆菌bl21star(de3)细胞接种预培养物,并使其在28℃和130rpm下在补充有100μgml-1氨苄青霉素的lb中生长过夜。将主要培养物在自诱导培养基(autoinductionmedium,aim)中接种至od600为0.1,所述aim是含有100μgml-1氨苄青霉素的包含微量元素的terrific肉汤基础培养基(terrificbrothbase,来自于formedium,uk)。使重组蛋白在28℃和130rpm下表达22小时。通过在4400xg和22℃下离心10分钟收获细胞,并立即用于全细胞生物转化或在-20℃下冷冻直至进行蛋白质纯化。为了进行油酸(oa)和oa衍生物(即底物(1)至(10))的生物转换测定,将50od600单位的由此制备的细胞(对应于50mg细胞干重)在玻璃培养管(corning,ny)中重悬于补充有100mm葡萄糖和0.2mmfad的50mmhepes,ph6.0中。通过从乙醇储备溶液(100mm)中添加底物至终浓度为2mm,开始进行1ml规模的生物转化。将正十五烷酸(1mm)用作内标。在限定的管角度(55°)下,在30℃和150rpm振荡下在作为共溶剂的2%(v/v)的乙醇的存在下进行反应。生物转化进行22小时或96小时。通过用0.12mhcl酸化至ph2.0来猝灭测定体系,并且在vibraxvxr基本型摇床(ika,germany)上搅动30分钟的同时将脂肪酸衍生物用2ml乙酸乙酯提取两次。将悬浮液在2900xg和22℃下离心5分钟以改善相分离。将合并的有机相在n2流下浓缩。将脂肪酸衍生物用10μl吡啶和50μln,o-双(三甲基甲硅烷基)三氟乙酰胺(bstfa)甲硅烷基化。在500rpm下孵育30分钟后,将提取物用200μl乙酸乙酯稀释,并通过gc-ms分析。实施例4:fad辅因子的还原和厌氧体外转换为了还原与酶结合的黄素辅因子,将野生型ohya和突变型q265a/t436a/n438a在含有50mmnacl、5mmedta、1μm5-脱氮核黄素和4μm甲基紫精的50mmhepes,ph7.4中稀释,以得到在440nm下约0.2au的吸收。将溶液转移到比色皿中,并通过在手套箱中孵育至少2小时去除氧气。之后,通过厌氧光还原或用二硫苏糖醇(dtt)进行化学还原来还原该辅因子。对于厌氧光还原,通过用常规led(lumineanc-691;pearlgmbh,buggingen,germany;10w,5400k)照射,将fad辅因子还原150分钟。在光还原期间,将比色皿冷却至10℃以补偿光源产生的热量。对于化学还原,将100倍摩尔过量的dtt添加到样品中。在厌氧条件下用还原的ohya和氧化的ohya进行活性测定。测定反应含有在2%(v/v)乙醇存在下,0.15mgml-1还原的ohya或氧化的ohya和以1ml规模在50mmhepes,ph6.0中的2mm底物(1)、(6)和(7)。在手套箱中使用正十五烷酸作为内标,将oa(即底物(1))的转换反应在手动振荡下在21℃孵育10分钟。与oa酯底物(6)和(7)的反应在玻璃培养管中以55°角在25℃和150rpm下孵育过夜。然后停止反应以执行如本文所述进行的gc-ms分析。实施例4:用大肠杆菌全细胞以半制备规模进行oa衍生物的水合以半制备规模进行oa衍生物,即底物(3)至(10)的水合。将20-150mg的非生理底物以1ml规模的全细胞生物转换进行转换。每个反应在过表达ohyaq265a/t436a/n438a后在玻璃培养管中含有200mg大肠杆菌细胞,然后重悬于含有100mm葡萄糖和0.2mmfad的50mmhepes,ph6.0中。生物转化反应在限定的管角度(55°)下在30℃和150rpm下孵育96小时。在通过用0.12mhcl酸化至ph2.0而猝灭后,将悬浮液用乙酸乙酯提取(3×2ml提取30分钟),并在2900xg和22℃下间歇离心5分钟以改善相分离。定量收集有机相并在n2流下浓缩。结果显示在表3中。表3:从大肠杆菌细胞提取物分离的oa衍生物和水合产物的收率。条目起始材料/mg回收的起始材料/mg分离的水合产物/mg3204605207.86.368062.45.478060.910.488059.11.696010150底物(1)至(10)的生物转换是用全大肠杆菌细胞进行的——大肠杆菌空载体对照(evc)和在ohya过表达之后的细胞进行的生物转化——其中底物(1)为油酸,底物(3)为油酰胺,底物(4)为要转换成n,10-二羟基硬脂酸酰胺的n-羟基油酰胺,底物(5)为要转换成1,10-十八烷二醇的油醇,底物(6)为要转换成10-羟基十八烷酸甲酯的oa甲酯(油酸甲酯),底物(7)为要转换成10-羟基十八烷酸乙酯的oa乙酯(油酸乙酯),底物(8)为要转换成10-羟基十八烷酸异丙酯的oa异丙酯(油酸异丙酯),底物(9)为要转换成10-羟基十八烷酸正丙酯的oa正丙酯(油酸正丙酯),并且底物(10)为要转换成10-羟基十八烷酸正丁酯(未显示)的oa正丁酯(油酸正丁酯)。oa衍生的异羟肟酸(即底物(4))和水合反应产物在gc-ms分析条件下发生洛森重排(lossenrearrangement)后均被检测到为相应的异氰酸酯。此外,用大肠杆菌evc和表达ohya的菌株转换底物(4)导致了油酰胺(即底物(3))的意外形成,所述油酰胺随后仅在ohya生物转化中水合为10-羟基硬脂酸酰胺。由于底物(4)最初不含油酰胺,因此必须假定油酰胺是通过底物(4)在大肠杆菌中的降解形成的。使用底物(3)至(10)的转换率如图2所示。表4中显示了针对全细胞大肠杆菌生物催化剂的生物质归一化的数值。表4:针对带有用于进行油酸及其衍生物的区域选择性和立体选择性水合的ohya野生型(wt)和突变型ohyaq265a/t436a/n438a酶的全细胞大肠杆菌生物催化剂的生物质归一化的表观水合活性。实施例5:用无细胞提取物(cfe)以半制备规模进行oa衍生物的水合在玻璃培养管中用2mg的重组蛋白质表达之后的大肠杆菌cfe进行体外活性测定。将cfe与2mm底物(1)至(10)一起在1ml的50mmhepes,ph6.0和2%(v/v)乙醇中孵育。在作为内标的1mm正十五烷酸存在下,将测定反应在25℃和150rpm下振荡过夜。如实施例2中所述,将转换反应猝灭,并且提取脂肪酸并衍生化。在还原fad辅因子后,用纯化的野生型ohya和突变型ohyaq265a/t436a/n438a进行底物(1)和油酸酯底物(6)和(7)的厌氧体外水合反应。在10分钟后将含有底物(1)的反应物猝灭,并且在过夜孵育后将含有底物(6)或(7)的反应物猝灭(未显示)。在底物(6)的转换中,在与真实oa标准品、野生型ohya和突变型ohyaq265a/t436a/n438a孵育后,如通过gc-ms分析证实的(未显示),我们仅在所述变体的情况下观察到了水合产物的预期保留时间的峰。序列表<110>帝斯曼知识产权资产管理有限公司<120>油酸衍生物的选择性水合<130>case32973-wo-pct<160>12<170>patentinversion3.5<210>1<211>646<212>prt<213>脑膜炎败血伊丽莎白菌(elizabethkingiameningoseptica)<400>1metasnproilethrserlyspheasplysvalleuasnalaserser151015glutyrglyhisvalasnhisgluproaspserserlysgluglngln202530argasnthrproglnlyssermetpropheseraspglnileglyasn354045tyrglnargasnlysglyileprovalglnsertyraspasnserlys505560iletyrileileglyserglyilealaglymetseralaalatyrtyr65707580pheileargaspglyhisvalproalalysasnilethrpheleuglu859095glnleuhisileaspglyglyserleuaspglyalaglyasnprothr100105110aspglytyrileileargglyglyargglumetaspmetthrtyrglu115120125asnleutrpaspmetpheglnaspileproalaleuglumetproala130135140protyrservalleuaspglutyrargleuileasnaspasnaspser145150155160asntyrserlysalaargleuileasnasnlysglygluilelysasp165170175pheserlyspheglyleuasnlysmetaspglnleualaileilearg180185190leuleuleulysasnlysglugluleuaspaspleuthrilegluasp195200205tyrphesergluserpheleulysserasnphetrpthrphetrparg210215220thrmetphealaphegluasntrphisserleuleugluleulysleu225230235240tyrmethisargpheleuhisalaileaspglyleuasnaspleuser245250255serleuvalpheprolystyrasnglntyraspthrphevalthrpro260265270leuarglyspheleuglnglulysglyvalasnilehisleuasnthr275280285leuvallysaspleuaspilehisileasnthrgluglylysvalval290295300gluglyileilethrgluglnaspglylysgluvallysileproval305310315320glylysasnasptyrvalilevalthrthrglysermetthrgluasp325330335thrphetyrglyasnasnlysthralaproileileglyileaspasn340345350serthrserglyglnseralaglytrplysleutrplysasnleuala355360365alalyssergluilepheglylysproglulysphecysserasnile370375380glulysseralatrpgluseralathrleuthrcyslysproserala385390395400leuileasplysleulysglutyrservalasnaspprotyrsergly405410415lysthrvalthrglyglyileilethrilethraspserasntrpleu420425430metserphethrcysasnargglnprohispheprogluglnproasp435440445aspvalleuvalleutrpvaltyralaleuphemetasplysglugly450455460asntyrilelyslysthrmetleuglucysthrglyaspgluileleu465470475480alagluleucystyrhisleuglyilegluaspglnleugluasnval485490495glnlysasnthrilevalargthralaphemetprotyrilethrser500505510metphemetproargalalysglyaspargproargvalvalproglu515520525glycyslysasnleuglyleuvalglyglnphevalgluthrasnasn530535540aspvalvalphethrmetgluserservalargthralaargileala545550555560valtyrlysleuleuasnleuasnlysglnvalproaspileasnpro565570575leuglntyraspilearghisleuleulysalaalalysthrleuasn580585590aspasplysprophevalglygluglyleuleuarglysvalleulys595600605glythrtyrphegluhisvalleuproalaglyalaalaglugluglu610615620gluhisgluserpheilealagluhisvalasnlyspheargglutrp625630635640vallysglyilearggly645<210>2<211>1974<212>dna<213>脑膜炎败血伊丽莎白菌(elizabethkingiameningoseptica)<400>2atgcatcaccatcaccatcaccatcaccatcacaacccaatcaccagcaaattcgacaaa60gtcctgaacgcatccagcgagtacggccacgttaatcacgaaccggatagcagcaaagag120cagcaacgcaacaccccgcagaagtccatgccatttagcgatcaaatcggcaactatcaa180cgtaacaaaggtattccggttcagagctatgataattcgaagatttacatcattggttct240ggtattgcgggtatgtcggctgcgtactacttcatccgtgacggtcacgttccggcgaag300aacatcacgttcctggagcaactgcacattgatggcggctctctggatggtgctggcaac360ccgaccgacggctatatcatccgtggtggtcgtgaaatggatatgacctacgagaacctg420tgggatatgttccaggatattccggcgctggagatgccggcaccgtatagcgttctggat480gaatatcgtctgattaatgacaacgatagcaattacagcaaagcacgtctgatcaacaat540aagggcgaaatcaaggacttcagcaagtttggtctgaataagatggaccagctggccatc600atccgtctgttgctgaaaaacaaagaagagctggacgacttgaccattgaggactacttc660tctgagagctttctgaaaagcaatttctggacgttttggcgcacgatgttcgcgtttgag720aactggcatagcctgttggaactgaagctgtacatgcaccgcttcctgcacgccattgac780ggtctgaacgacctgagcagcctggtgtttccgaagtacaatcaatatgacacgtttgtc840acgccgctgcgtaaattcctgcaagaaaagggtgttaacatccacttgaataccttggtc900aaggatctggatattcacatcaataccgagggtaaagtcgtcgagggcatcattaccgag960caagacggtaaagaggtaaagattccggtgggtaagaatgactatgttatcgtgacgacc1020ggttccatgaccgaggacacgttttacggtaacaacaaaaccgcaccgatcattggtatc1080gacaatagcactagcggtcagagcgctggctggaaactgtggaagaacctggctgccaag1140agcgaaatcttcggcaagccggagaaattctgtagcaatattgagaaatccgcgtgggaa1200agcgcgaccctgacgtgtaaaccttccgcgttgatcgacaaactgaaagaatattcggtc1260aacgacccgtacagcggtaagaccgtgaccggcggtatcattactatcaccgatagcaac1320tggttgatgagctttacctgcaatcgccagccgcatttcccggagcagccggatgacgtc1380ctggtgctgtgggtgtatgcgctgtttatggataaagaaggtaactacattaagaaaacc1440atgctggagtgcaccggtgatgagattttggcggagctgtgttaccatctgggcattgaa1500gatcagctggaaaatgtgcagaagaatacgattgttcgcaccgcattcatgccgtatatc1560acgagcatgtttatgccacgtgccaaaggtgaccgccctcgtgtggtcccggaaggttgc1620aaaaacctgggcctggttggtcaatttgtggaaacgaacaatgacgtcgtgtttacgatg1680gaatctagcgttcgcacggcccgtattgcggtgtacaagttgctgaatctgaacaagcag1740gtgccggacattaatccgctgcaatacgatattcgccatctgctgaaagcggcaaagacc1800ctgaatgatgacaaaccgttcgtgggcgaaggcttgctgcgtaaggttctgaaaggcacc1860tattttgagcacgttctgcctgcgggtgcagcggaagaagaagagcatgagagcttcatt1920gcggaacatgttaacaagttccgtgagtgggtcaagggtatccgtggctaataa1974<210>3<211>34<212>dna<213>人工序列<220><223>引物<400>3gtttccgaagtacaatgcatatgacacgtttgtc34<210>4<211>34<212>dna<213>人工序列<220><223>引物<400>4gacaaacgtgtcatatgcattgtacttcggaaac34<210>5<211>33<212>dna<213>人工序列<220><223>引物<400>5tggttgatgagctttgcgtgcaatcgccagccg33<210>6<211>33<212>dna<213>人工序列<220><223>引物<400>6cggctggcgattgcacgcaaagctcatcaacca33<210>7<211>35<212>dna<213>人工序列<220><223>引物<400>7gatgagctttacctgcgcacgccagccgcatttcc35<210>8<211>35<212>dna<213>人工序列<220><223>引物<400>8ggaaatgcggctggcgtgcgcaggtaaagctcatc35<210>9<211>35<212>dna<213>人工序列<220><223>引物<400>9ctgcaatcgccagccggccttcccggagcagccgg35<210>10<211>35<212>dna<213>人工序列<220><223>引物<400>10ccggctgctccgggaaggccggctggcgattgcag35<210>11<211>35<212>dna<213>人工序列<220><223>引物<400>11gatgagctttgcgtgcgcacgccagccgcatttcc35<210>12<211>35<212>dna<213>人工序列<220><223>引物<400>12ggaaatgcggctggcgtgcgcacgcaaagctcatc35当前第1页1 2 3
起点商标作为专业知识产权交易平台,可以帮助大家解决很多问题,如果大家想要了解更多知产交易信息请点击 【在线咨询】或添加微信 【19522093243】 与客服一对一沟通,为大家解决相关问题。
与客服一对一沟通,为大家解决相关问题。
此文章来源于网络,如有侵权,请联系删除

 热门咨询
热门咨询




tips