
 商标分类
商标分类  商标转让
商标转让 
一个水稻粒长基因的基因工程应用的制作方法
 2021-02-02 16:02:57|
2021-02-02 16:02:57| 439|
439| 起点商标网
起点商标网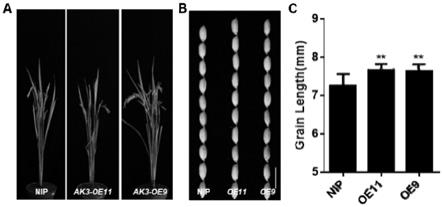
[0001]
本发明属于植物基因工程技术领域,涉及一种控制水稻粒长基因osak3的克隆与应用。
背景技术:
[0002]
水稻是世界上最重要的农作物之一。提高水稻产量是全球人口快速增长的迫切需求。粒型是决定水稻产量的主要因素之一,因此粒型一直是水稻育种的重要目标。水稻粒型性状主要包括粒长、粒宽、粒厚、长宽比,属于数量性状,受胚、胚乳及母体植株等不同遗传体系的控制。水稻粒型还影响到稻米的外观品质和商品价值。在中国南方、美国和大多数亚洲国家,人们更习惯长或细长型稻米,而韩国、日本、中国北方则习惯短圆型稻米(harberd n p,2015)。此外,粒型也是进化中一个易于被选择的表型,是研究水稻进化的一个重要性状(meyer r s,2013)。因此,克隆粒型相关的基因并深入研究和阐明粒型形成的基因调控网络,可为水稻高产、优质分子育种提供重要的理论基础和基因资源。
[0003]
目前的研究表明,水稻粒型受多个遗传体系所控制,已知的有g蛋白信号通路、泛素—蛋白酶体通路、mapk信号通路、表观修饰及植物激素信号通路等多个水稻粒型调控通路。gs3是第一个被鉴定出来的水稻粒型qtl。利用明恢63(大粒)和川7(小粒)杂交获得的近等基因系,通过图位克隆的方法定位到了gs3。该基因编码一个g蛋白γ亚基,gs3功能的缺失促进细胞伸长,进而产生长粒(fan et al,2006)。而水稻g蛋白α亚基d1功能缺失突变体d-1表现矮秆小粒表型,对br的敏感性降低,表明d1介导的异源三聚体g蛋白与br信号转导途径共同调控水稻粒型(miura k et al,2009)。g蛋白β亚基(rgb1)表达量的降低也会导致水稻出现籽粒变小的表型(mao h et al,2010)。gw2是控制粒宽的一个主效qtl,该基因编码一个含有ring环的e3泛素连接酶,功能缺失突变体小穗外壳细胞增殖增强,加速了籽粒灌浆,从而提高了粒宽、粒重和产量(song et al,2007)。脱泛素酶osotub1/wtg1的功能缺失会使粒宽增加,籽粒变厚,每穗粒数也增多(huang et al,2017)。osotub1主要通过影响细胞扩增来调控粒型。osotub1和理想株型的重要调控因子osspl14/ipa1互作,这种互作限制了osspl14的k63连接泛素化,从而促进了k48连接泛素化依赖性蛋白酶体降解(wang et al,2017)。mapk级联是真核生物中进化保守的信号模块,在植物发育和防御信号转导中起着关键作用(meng 2013;zhang 2015)。最近的研究发现,osmkk4功能的缺失导致小粒(duan et al,2014)。osmapk6也在粒型的调控中起到正调控的作用(liu et al,2015)。osmkk4可以与osmapk6互作并且磷酸化osmapk6(liu et al,2015),并且这两个蛋白可以影响油菜素内酯(br)的响应,可以调控br相关基因的表达。这说明不同的粒型调控通路之间是有关联的。
[0004]
在植物激素调控水稻粒型的网络中,发挥主要作用的激素有油菜素内酯、生长素、细胞分裂素等。其中油菜素内酯是一类多羟基甾体植物激素,对植物生长发育过程中多种生理过程的合理调控起到至关重要的作用。br缺乏突变体dwarf1和dwarf2会产生小而短的籽粒,说明br可以促进籽粒的生长(hong et al,2005;fang et al,2016)。br被定位在膜上
面的受体复合物bri1-bak1感知,进一步使bin2失活,激活两个转录因子bzr1和bzr2对下游基因的转录调控(clouse 2011)。在水稻中,osbri1和osbak1的功能缺失导致br钝感的表型并且籽粒变小(morinaka et al,2006)。而过表达osbzr1会增长水稻粒长、粒宽和粒重(zhu et al,2015)。bin2的同源基因osgsk2负调控水稻粒型(tong et al,2012)。在水稻中,osgsk2可以磷酸化正调控因子osdlt(tong et al,2012)。qgl3是控制粒长的一个主效qtl,编码一个丝氨酸/苏氨酸磷酸酶(osppkl1),含有两个kelch结构域。水稻中还有osppkl1的两个同系物osppkl2和osppkl3。osppkl1和osppkl3会抑制籽粒长度,而osppkl2作为拟南芥atbsu1和atbsl1的同系物起到促进籽粒伸长的作用。研究osppkls是否通过br信号途径调控水稻粒长具有重要意义。我们利用qgl3/osppkl1作为诱饵蛋白筛选水稻幼穗cdna表达文库,得到了一个编码腺苷酸激酶的基因osak3。
技术实现要素:
[0005]
本发明的目的在于公开一个控制水稻籽粒粒长的基因osak3的克隆与水稻粒长性状的基因工程应用。过量表达水稻基因osak3可以增加水稻粒长,利于水稻粒型的遗传改良。osak3功能缺失突变体籽粒粒长变短,株高变矮,分蘖数减少,可用于水稻产量性状的遗传改良。
[0006]
本发明的第一个目的是提供水稻基因osak3,所述osak3基因的cdna序列如seq id no.1所示,所述水稻基因osak3的编码区位于seq id no.1所示cdna序列的154bp~879bp,全长726bp。
[0007]
所述水稻基因osak3编码的氨基酸序列如seq id no.2所示。
[0008]
本发明的第二个目的是提供含有前述水稻基因osak3的编码区的过量表达载体。
[0009]
进一步的,所述的过量表达载体是将前述水稻osak3基因的编码区插入植物双元表达载体pcambia1300s的kpn
ꢀⅰ
和sal
ꢀⅰ
酶切位点间所得。
[0010]
本发明的第三个目的是提供前述水稻基因osak3或前述水稻基因osak3的过量表达载体在基因工程中的应用。
[0011]
本发明的第四个目的是提供前述水稻基因osak3或前述水稻基因osak3的过量表达载体在通过基因工程手段改良水稻粒长中的应用。
[0012]
进一步的,过量表达所述水稻基因osak3,能够增加水稻的粒长。
[0013]
进一步的,将水稻基因osak3的过量表达载体导入水稻,提高水稻基因osak3的表达,能够增加水稻的粒长。
[0014]
进一步的,前述应用具体包括以下步骤:
[0015]
(1)提取水稻品种“dj”的总rna,反转录合成cdna第一链;
[0016]
(2)水稻基因osak3的克隆:
[0017]
以步骤(1)反转录合成的cdna第一链为模板,采用引物对:
[0018]
osak3f:5
’-
ttgtcaccgtctcaccag-3
’
(seq id no.3),
[0019]
osak3r:5
’-
cagaaggaataccctgtcaagatt-3
’
(seq id no.4)进行pcr扩增,将克隆片段连入peasy blunt simple载体,转入大肠杆菌菌株dh5α中,测序筛选得到含有seq id no.1所示的水稻osak3基因cdna序列的重组质粒t-osak3;
[0020]
(3)过量表达载体的构建:根据水稻osak3基因的cdna序列,设计扩增出完整编码
阅读框的引物对:
[0021]
osak3oef:5
’-
agctttcgcgagctcggtaccatggcggcgaacctgg-3
’
(seq id no.5),包含入kpn
ꢀⅰ
限制性内切酶位点,
[0022]
osak3oer:5
’-
cttgcatgcctgcaggtcgacttatgatagggctttctggacctc-3
’
(seq id no.6),包含sal
ꢀⅰ
限制性内切酶位点。
[0023]
以步骤(2)获得的测序正确的重组质粒t-osak3为模板,进行pcr扩增,得到水稻osak3基因的编码区,将扩增产物插入植物双元表达载体pcambia1300s的kpn
ꢀⅰ
和sal
ꢀⅰ
酶切位点间,获得过量表达载体pcambia1300s-osak3;
[0024]
(4)转基因植株的获得:将获得的过量表达载体pcambia1300s-osak3通过热激法转入农杆菌菌株eha105,利用农杆菌介导的水稻遗传转化将过量表达载体的t-dna区整合到水稻粳稻品种日本晴的基因组中,从而获得osak3过量表达的转基因水稻。
[0025]
有益效果
[0026]
1、本发明公开了一种过量表达水稻osak3基因的粒型基因工程应用。该基因来自水稻(oryza sativa l.),过量表达该基因可以增加水稻粒长,利于水稻粒型的遗传改良。
[0027]
2、本发明中克隆出来的osak3基因为水稻的高产优质育种提供了新资源。
附图说明
[0028]
图1.重组载体pcambia1300s-osak3的构建
[0029]
a.过量表达载体pcambia1300s酶切位点示意图;b.osak3过量表达转基因株系qrt-pcr鉴定。
[0030]
图2.osak3过量表达转基因株系的表型
[0031]
a.osak3过量表达转基因株系的株型与野生型的对比图;b.osak3过量表达转基因株系的粒型与野生型的对比图;c.osak3过量表达转基因株系的粒长统计图。
[0032]
图3.osak3t-dna插入突变体osak3的表型
[0033]
a-d.分别为osak3的粒长、株高、剑叶长度与宽度、节间数与野生型的对比图;
[0034]
e.osak3的粒长统计图;f.osak3的株高统计图;g.osak3的分蘖数统计图。
具体实施方式
[0035]
实施例1
[0036]
选用水稻品种“dj”,于人工气候培养箱(16h光照/8h黑暗,白天30℃夜晚26℃)中水培生长,营养液采用国际水稻所常规营养液配方。待幼苗生长至3-4叶期时,取幼苗于液氮中速冻并于-80℃冰箱中保存备用。取保存的水稻幼苗样品,参照invitrogen公司的trizol法提取总rna的提取。总rna的质量以及浓度通过1%琼脂糖凝胶电泳分析,获得符合质量的总rna进一步用于合成cdna第一链。cdna第一链的合成参照vazyme公司反转录系统的操作手册进行。设计引物,上游osak3f:5
’-
ttgtcaccgtctcaccag-3
’
(seq id no.3);下游osak3r:5
’-
cagaaggaataccctgtcaagatt-3
’
(seq id no.4),以反转录合成的cdna第一链为模板,采用pcr方法进行cdna克隆。pcr扩增使用primestar gxl dna聚合酶(takara,dalian,china)。pcr程序如下:98℃预变性5min,98℃变性15s,56℃复性15s,72℃延伸45s,28个循环后,72℃10min,将pcr产物进行琼脂糖凝胶电泳,使用vazyme公司的dna凝胶回收
试剂盒回收目的片段。将克隆片段连入peasy blunt simple载体,利用热激转化法将其转入大肠杆菌菌株dh5α中,获得质粒t-osak3,测序后获得具有完整编码区的osak3 cdna序列seq id no.1。osak3 orf全长726bp,bioxm软件(版本2.6)分析表明osak3共编码241个氨基酸seq id no.2,估算其等电点pi=8.47,分子量mw=26.41kda。
[0037]
实施例2
[0038]
根据水稻osak3基因的cdna序列,设计扩增出完整编码阅读框的引物,上游引物osak3oef:5
’-
agctttcgcgagctcggtaccatggcggcgaacctgg-3
’
(seq id no.5)包含入kpn
ꢀⅰ
限制性内切酶位点,下游引物osak3oer:5
’-
cttgcatgcctgcaggtcgacttatgatagggctttctggacctc-3
’
(seq id no.6)包含sal
ꢀⅰ
限制性内切酶位点。以实施例1中获得的测序正确的重组载体t-osak3为模板,进行pcr扩增,所得产物使用vazyme公司的cloneexpress试剂盒,进一步克隆到植物双元表达载体pcambia1300s(图1a),获得过量表达载体pcambia1300s-osak3。
[0039]
将获得的重组表达载体pcambia1300s-osak3通过热激法转入农杆菌菌株eha105,利用农杆菌介导的水稻遗传转化将重组载体的t-dna区整合到水稻粳稻品种日本晴的基因组中,从而获得osak3过量表达的转基因水稻oe9、oe11。
[0040]“nip”野生型以及osak3过量表达的转基因水稻成苗后,对各株系进行qrt-pcr鉴定。取少量转基因水稻叶片,参照invitrogen公司的trizol法提取总rna的提取,反转成cdna。设计osak3基因qrt-pcr特异引物qosak3f:5
’-
tccacgtacaaactaagcctgtg-3
’
(seq id no.7)、qosak3r:5
’-
ggctttctggacctcaacagtc-3
’
(seq id no.8),以水稻actin rrna的表达为内参,按照roche公司fast start universal sybr green master(rox)操作说明对转基因株系oe9、oe11进行qrt-pcr验证。验证结果表明两个osak3过量表达株系oe9、oe11中osak3表达量相对于“nip”野生型均有提高(图1b)。
[0041]
实施例3
[0042]
本实验分为两组:第一组为将从韩国突变体库购买的dj背景osak3 t-dna插入突变体osak3,及野生型(dj);第二组为实施例2得到的过表达转基因株系(oe9、oe11)及野生型(nip)。种子分别浸种露白后,播种于牌楼,用于成株期农艺性状考察。每个株系3行,行距20cm,每行种植10株,每株间距15cm,常规土肥管理。灌浆结束后,量取转基因和野生型水稻每行中间8株的株高、分蘖数,进行统计分析。
[0043]
突变体osak3株系,osak3过量表达株系(oe9、oe11)、野生型株系(dj、nip)种子成熟后,分别将各组水稻的穗茎节剪下,种子42℃烘干5d后进行粒长考种分析。
[0044]
统计结果显示,实施例2得到的osak3过量表达株系(oe9、oe11)的水稻籽粒粒长显著长于野生型nip,株高无显著变化(图2)。说明osak3过量表达可以增加水稻粒长,有利于水稻粒型的遗传改良。
[0045]
而缺失表达突变体osak3的粒长、株高、分蘖数均显著低于野生型dj,突变体osak3的剑叶长度变短、宽度变窄,节间长度变短,节间数增加(图3),说明osak3参与水稻粒长发育的正调控。
起点商标作为专业知识产权交易平台,可以帮助大家解决很多问题,如果大家想要了解更多知产交易信息请点击 【在线咨询】或添加微信 【19522093243】 与客服一对一沟通,为大家解决相关问题。
与客服一对一沟通,为大家解决相关问题。
此文章来源于网络,如有侵权,请联系删除

 热门咨询
热门咨询




tips