
 商标分类
商标分类  商标转让
商标转让 
植物花粉特异表达启动子POsPTD1及应用的制作方法
 2021-02-02 14:02:46|
2021-02-02 14:02:46| 378|
378| 起点商标网
起点商标网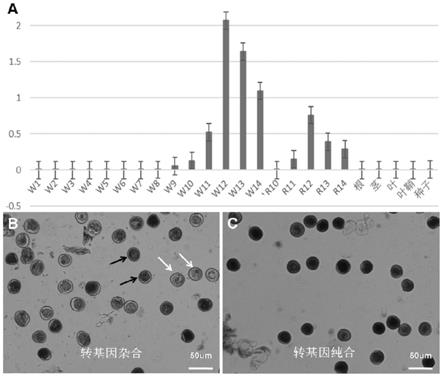
植物花粉特异表达启动子posptd1及应用
技术领域
[0001]
本发明属于基因工程领域,涉及一种植物花粉特异性启动子posptd1及应用,该启动子能驱动目的基因或dna片段在花粉中特异表达,从而改变花粉育性,控制转基因漂移、制备雄配子体不育和繁殖孢子体核雄性不育等。
背景技术:
[0002]
启动子一般位于基因编码区的上游,启动子中含有顺式作用元件,是一些特殊的dna序列,决定特殊转录因子的结合,从而实现基因的转录调控。启动子还提供rna聚合酶定位信息,rna聚合酶识别并结合启动子相关区域,以启动rna的转录。因此,启动子是基因转录所必须,特殊的启动子决定与之相连的基因的特殊表达模式。筛选和分离一些具有组织或器官高表达水平的启动子以及分析有关的重要特异性调控元件功能,在基因工程研究中有着广泛的用途。
[0003]
花粉是高等植物有性生殖的两个主要参与者之一。相对于不动的雌配子体,花粉能随多种媒介在植物个体乃至物种间移动,这对植物的基因交流和变异积累异常重要。花粉的发育调控需要众多基因参与,这些基因有的在花药壁中特异表达,有的在花粉中特异表达,有的在花药壁和花粉中都表达。在花粉中特异表达的基因的启动子为花粉特异启动子。
[0004]
在植物杂种优势利用中,雄性不育系是一种重要的遗传材料,在自花繁殖作物的杂种优势利用中更是极为必要。雄性不育包括孢子体不育和配子体不育,其中的孢子体不育又包括孢子体核不育和孢子体质不育。孢子体核不育比孢子体质不育有更大的杂种优势利用潜力,但其固有的遗传特点使得其难以繁殖出全不育群体,必须开发相应的孢子体核不育全不育群体的繁殖方法。
[0005]
花粉特异启动子可驱动影响花粉发育的一些基因在花粉中表达,影响花粉的正常发育,可用于繁殖孢子体雄性核不育系(wu等,2015;chang等2016)。目前发现的花粉特异启动子不多,主要有番茄花粉特异启动子lat52,水稻花粉特异ra8基因启动子、玉米zmc5基因启动子、zm13和pg47启动子等等。
[0006]
随着各种植物基因组测序完成和各种基因表达谱数据的公开,借助公共数据库可以筛选到在水稻花粉中特异性高效表达的功能基因。这些功能基因的上游dna序列成为花粉特异性启动子的候选者。
[0007]
本发明希望提供一种雄配子体特异性启动子,为防止转基因随花粉漂移,为雄性不育植物基因工程和杂交水稻研究提供遗传资源。
技术实现要素:
[0008]
有鉴于此,本发明的目的之一在于提供植物花粉中特异表达的启动子;本发明的目的之二在于提供植物花粉特异性启动子posptd1的互补核酸序列;本发明的目的之三在于提供含有所述植物花粉特异性启动子posptd1或互补核苷酸序列的重组表达载体;本发
明的目的之四在于提供含有所述植物花粉特异性启动子posptd1或互补核苷酸序列的宿主细胞;本发明的目的之五在于提供所述植物花粉特异表达启动子posptd1在植物花粉中特异驱动目的基因表达中的应用;本发明的目的之六在于提供在植物花粉中驱动特定目的片段的方法;本发明的目的之七在于提供植物花粉特异表达启动子posptd1的应用。
[0009]
为达到上述目的,本发明提供如下技术方案:
[0010]
1、植物花粉特异表达启动子posptd1,所述植物花粉特异表达启动子posptd1至少包含:
[0011]
(a)seq id no.1中所示的核苷酸序列;
[0012]
(b)或者由seq id no.1所示的核苷酸序列经取代、缺失或添加至少一个核苷酸而衍生的且具有同等花粉特异性启动功能的核苷酸序列。
[0013]
2、所述植物花粉特异性启动子posptd1的互补核酸序列。
[0014]
3、含有所述植物花粉特异性启动子posptd1或互补核苷酸序列的重组表达载体。
[0015]
4、含有所述植物花粉特异性启动子posptd1或互补核苷酸序列的宿主细胞。
[0016]
5、所述植物花粉特异表达启动子posptd1在植物花粉中特异驱动目的基因表达的应用。
[0017]
本发明中,所述植物为开花植物,如禾本科植物,也可以为其他开花植物;优选为水稻、玉米、高粱、小麦、大麦、燕麦、黑麦或油菜。
[0018]
6、在植物花粉中驱动特定目的片段表达的方法,具体步骤如下:
[0019]
(1)将权利要求1所述的植物花粉特异表达启动子posptd1与目的片段连接,使posptd1在目的片段上游,然后载入植物表达载体;
[0020]
(2)将步骤(1)所得植物表达载体通过根癌农杆菌介导转化植物,筛选转基因植株,获得花粉中驱动特定目的片段表达的植物。
[0021]
本发明中,所述植物为开花植物,如禾本科植物,也可以为其他开花植物;优选为水稻、玉米、高粱、小麦、大麦、燕麦、黑麦或油菜。
[0022]
优选的,所述目的片段为结构基因或调节基因,或者结构基因或调节基因的突变基因,或者结构基因或调节基因的下调表达元件。
[0023]
7、所述植物花粉特异表达启动子posptd1的应用为以下任意一种:
[0024]
(1)培育转基因成分不随花粉漂移的植物品种;
[0025]
(2)培育雄性配子体不育植物材料;
[0026]
(3)繁殖雄性孢子体核不育系。
[0027]
本发明的有益效果在于:本发明所涉及的启动子及干涉片段均来自水稻,可减少外源转基因成分,降低转基因风险和减少人们对转基因产品的担忧;本发明所获得的水稻启动子posptd1能够调控基因在植株雄配子体中特异表达,通过该启动子驱动能影响花粉育性的基因或dna片段的表达,破坏含转基因成分的花粉的育性,可以制备雄性配子体不育,用于繁殖孢子体核不育系;本发明对于研究雄配子体不育机理,阻断花粉介导基因漂移,繁殖孢子体核不育等方面具有重要价值。
附图说明
[0028]
为了使本发明的目的、技术方案和有益效果更加清楚,本发明提供如下附图进行
说明:
[0029]
图1为雄配子体发育相关基因表达谱分析(a为loc_os05g40740基因在野生型各器官和花发育各时期和在干涉纯合后代花发育后期表达的qpcr分析;b为启动子posptd1驱动gus基因在花粉中特异表达的gus染色检测,显示转基因杂合体花粉的gus染色;c为启动子posptd1驱动gus基因在花粉中特异表达的gus染色检测,显示转基因纯合体花粉的gus染色;黑箭头示gus阳性花粉,白箭头示gus阴性花粉)。
[0030]
图2为载体posmyb76r多克隆位点改造后的酶切位点示意图;
[0031]
图3为转基因t
0
代阳性株杂交f
1
中紫线的传递情况:绝大多数无紫线,箭头示极少数紫线株
具体实施方式
[0032]
下面结合附图和具体实施例对本发明作进一步说明,以使本领域的技术人员可以更好的理解本发明并能予以实施,但所举实施例不作为对本发明的限定。
[0033]
实施例1、水稻雄配子体特异启动子posptd1的克隆
[0034]
雄配子体发育相关基因往往在中后期的花/花粉中表达。为了找到一些配子体发育相关基因,对bar水稻基因表达数据库(http://bar.utoronto.ca/efprice/cgi-bin/efpweb.cgi)进行分析,发现很多在中后期花中表达的基因,经qpcr验证,选出一批可能同雄配子体发育相关的基因。其中,水稻loc_os05g40740基因在后期花中特异高表达,其拟南芥的同源基因在花粉中特异表达,初步将loc_os05g40740基因列为雄配子体发育相关基因,将其启动子列为花粉特异启动子。随后,用qpcr对该基因的时空表达模式进行了分析,所用loc_os05g40740基因的qpcr引物为:上游引物mqf2:5'-ggcgtcttcaggtacaaccag-3'(seq id no.4),下游引物mqr25'-ggtacacctggaccgagtgc-3'(seq id no.5)。所用内参引物为上游引物osactin2f:5'-ctctgtatgccagtggtcgt-3'(seq id no.6),下游引物osactin2r:5'-ccgttgtggtgaatgagtaac-3'(seq id no.7)。qpcr结果表明:loc_os05g40740基因在根、茎、叶、叶鞘和种子中均不表达,在发育中后期的花中高表达(图1)。进一步分析干涉纯合植物花发育后期该基因的表达,发现干涉造成该基因的表达严重降低。
[0035]
随后,为了确定该基因是否在花粉中特异表达,克隆了该基因的启动子,将其置于pcambia1301的gus基因前面。具体操作为:
[0036]
用启动子上游引物p5f1:5'-gcgtcgacgtcccatgtcaccgacagtact-3
’
(seq id no.8,带酶切位点sali)和下游引物p5r1:5'-catgccatggcgtggaaatgtgatcgctaggct-3
’
(seq id no.9,带酶切位点ncoi)从中花11中扩出loc_os05g40740的启动子,其核苷酸序列如seq id no.1所示。用sali和ncoi(不完全酶切)将其载入载体pcambia1301的gus基因前面得p posptd1-gus。
[0037]
实施例2、水稻花粉特异启动子posptd1的花粉特异性的gus染色鉴定
[0038]
将posptd1-gus转入农杆菌lba4404,再将阳性农杆菌转染中花11。开花期对转基因阳性后代的根、叶和花进行gus染色观察,结果表明:根、叶、颖壳、花丝、雌蕊等均不染色,而花药染成蓝色。进一步捣碎花药,在显微镜下观察发现:转基因杂合的植株的gus染色花粉仅有50%左右,纯合的植株的花粉几乎全部能被gus染色(图1)。上述结果说明启动子
posptd1确实为花粉特异启动子。
[0039]
实施例3、通过posptd1驱动的rna干涉制备雄性配子体不育及鉴定
[0040]
为了确定posptd1启动子能否驱动目的基因或dna片段破坏花粉育性,制备和鉴定雄性配子体不育材料。以loc_os05g40740为对象,用posptd1启动子驱动loc_os05g40740基因的干涉片段,实施了雄性配子体不育创造和鉴定:
[0041]
1、为了便于载入相关目的基因或dna片段,为了准确和直接进行转基因阳性株以及雄配子体不育的鉴定,我们用水稻花青素合成相关基因的改造基因osmyb76r替换gus基因,并对载体pcambia1301的多克隆位点进行了改造,具体操作为:
[0042]
(1)osmyb76基因改造并代替gus:osmyb76基因在水稻中为花青素合成必须基因,可用作内源性报告基因;为了避免osmyb76基因本身带有的酶切位点影响载入目的片段,通过碱基替换,除去osmyb76基因中的常用酶切位点,获得osmyb76基因改造后的osmyb76r基因的cds序列并人工合成,其序列如seq id no.10所示。以合成的osmyb76r基因为模板,用引物osmyb76rf:5'-ggactcttgaccatgatgggacgcagggcttgct-3'(seq id no.11,带pcambia1301的bglii附近同源片段)和osmyb76rr:5'-attcgagctggtcactcacgcacacaggttccaag-3'(seq id no.12,带pcambia1301的bsteii附近同源片段)扩出目的片段,用同源重组方式将目的片段连入pcambia1301,得载体posmyb76r。
[0043]
(2)多克隆位点改造:对前述载体posmyb76r的多克隆位点的酶切位点种类和顺序进行改造。先人工合成新的多克隆位点序列,其核苷酸序列如seq id no.13所示。然后通过同源重组将合成序列替换原有多克隆位点的序列,新的多克隆酶切位点如图2所示。多克隆位点改造后的载体命名为posmyb76rm。
[0044]
2、用茎环结构上游引物lf2:5'-gaagatctacgagctggtgagctagcta-3
’
(seq id no.14,带酶切位点bglii)和下游引物lr2:5'-ctggtcacccctcaaacctgaaaattcag-3
’
(seq id no.15,带酶切位点bsteii)从r25中扩出osmyb76基因的第一内含子作干涉茎环(seq id no.2),载入posmyb76rm得载体posmyb76rl。
[0045]
3、用干涉左臂上游引物i5f1:5'-cggaattcgcagaaggtgatcctgatcaac-3
’
(seq id no.16,带酶切位点ecori)和下游引物i5r1:5'-gaagatctgcaggcggctgacgacgctgat-3
’
(seq id no.17,带酶切位点bglii)从中花11中扩出loc_os05g40740的干涉左臂;用干涉右臂上游引物i5f2:5'-gacctgcagggcagaaggtgatcctgatcaac-3
’
(seq id no.18,带酶切位点sbfi)和下游引物i5r2:5'-ctggtcaccgcaggcggctgacgacgctgat-3
’
(seq id no.19,带酶切位点bsteii)从中花11中扩出loc_os05g40740的干涉右臂。将左右臂分别载入载体posmyb76rl中的茎环两边,并用sbfi和asci将tnos终止子载入干涉右臂后面,得载体posmyb76rli,干涉片段的核苷酸序列为seq id no.3所示。
[0046]
4、用启动子上游引物p5f1:5'-gcgtcgacgtcccatgtcaccgacagtact-3
’
(seq id no.8,带酶切位点sali)和下游引物p5r2:5'-cggaattccgtggaaatgtgatcgctaggct-3
’
(seq id no.20,带酶切位点ecori)从中花11中扩出loc_os05g40740的启动子,其核苷酸序列如seq id no.1所示。将其载入载体posmyb76rli得雄性配子体不育载体posmyb76r-gm。
[0047]
5、将posmyb76r-gm转入农杆菌lba4404,再将阳性农杆菌转染所有器官无色的中九b。
[0048]
6、在t
0
代根据茎基部紫色有无选出阳性株,让其自交,并用阳性株作父本,给前述
未转基因的中九b授粉。对自交和杂交种子进行紫线调查,发现自交f
2
种子紫线有无分离比例符合1:1,不符合3:1,96.4%的杂交f
1
种子表现无紫线(图3),这说明转基因成分难以随花粉在代间进行传递,但随雌配子在代间进行传递,这正是雄性配子体不育的特征。因此,loc_os05g40740与雄配子体花粉发育有关,通过对其进行干涉,借助连锁的紫线性状的遗传调查,获得了雄性配子体不育材料并成功进行了遗传鉴定。
[0049]
对上述创造雄性配子体不育材料的遗传分析所用的f
1
种子,也可通过调查紫线个体的比例,实现鉴定花粉传递转基因成分的几率,通过剔除带紫线的个体,实现筛除由花粉传递的转基因污染。
[0050]
以上所述实施例仅是为充分说明本发明而所举的较佳的实施例,本发明的保护范围不限于此。本技术领域的技术人员在本发明基础上所作的等同替代或变换,均在本发明的保护范围之内。本发明的保护范围以权利要求书为准。
起点商标作为专业知识产权交易平台,可以帮助大家解决很多问题,如果大家想要了解更多知产交易信息请点击 【在线咨询】或添加微信 【19522093243】 与客服一对一沟通,为大家解决相关问题。
与客服一对一沟通,为大家解决相关问题。
此文章来源于网络,如有侵权,请联系删除

 热门咨询
热门咨询




tips