
 商标分类
商标分类  商标转让
商标转让 
使用融合蛋白进行T细胞受体重编程的组合物和方法与流程
 2021-02-02 03:02:57|
2021-02-02 03:02:57| 358|
358| 起点商标网
起点商标网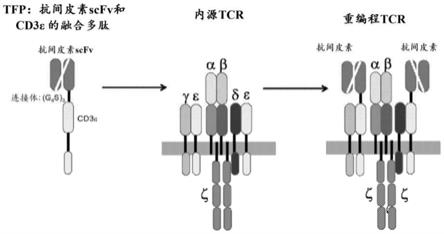
使用融合蛋白进行t细胞受体重编程的组合物和方法
本申请是申请日为2017年10月06日、申请号为201780039623.1、 发明名称为“使用融合蛋白进行t细胞受体重编程的组合物和方法”的 中国专利申请(其对应pct申请的申请日为2017年10月06日、申请 号为pct/us2017/055628)的分案申请。交叉引用
[0001]
本申请要求于2016年10月7日提交的美国临时申请号 62/405,551和于2017年5月23日提交的美国临时申请号62/510,108 的权益,这些临时申请各自通过引用整体并入本文。发明背景
[0002]
大多数晚期实体瘤患者无法用标准疗法治愈。此外,传统的治 疗选择通常会产生严重的副作用。已经进行了许多尝试以使患者的免 疫系统排斥癌细胞,这种方法统称为癌症免疫疗法。然而,一些障碍 使得实现临床有效性相当困难。尽管已经鉴定了数百种所谓的肿瘤抗 原,但这些抗原通常来源于自身,因此可引导癌症免疫疗法针对健康 组织,或者具有较差的免疫原性。此外,癌细胞使用多种机制使其自 身对癌症免疫疗法的免疫攻击而言不可见或抵抗其起始和传播。
[0003]
最近的发展使用嵌合抗原受体(car)修饰的自体t细胞疗法, 其依赖于将基因工程化t细胞重定向至癌细胞上合适的细胞表面分 子,这在利用免疫系统的能力治疗b细胞恶性肿瘤方面显示出有希望 的结果(参见例如,sadelain等人,cancer discovery 3:388-398(2013))。 cd-19特异性car t细胞(称为ctl019)的临床结果显示患有慢性 淋巴细胞白血病(cll)以及儿童急性淋巴母细胞白血病(all)的 患者完全缓解(参见例如,kalos等人,sci transl med 3:95ra73(2011),porte等人,nejm 365:725-733(2011),grupp等人,nejm 368:1509
-ꢀ
1518(2013))。替代方法是使用针对肿瘤相关肽抗原选择的t细胞受 体(tcr)α链和β链对自体t细胞进行基因工程化。这些tcr链将 形成完整的tcr复合物,并为t细胞提供tcr以具有第二限定特异 性。在滑膜癌患者中用表达ny-eso-1特异性tcrα链和β链的工程 化自体t细胞获得了令人鼓舞的结果。
[0004]
除了表达car或第二tcr的遗传修饰的t细胞在体外/离体识 别和破坏相应靶细胞的能力之外,用工程化t细胞进行成功的患者治 疗还需要t细胞能够随时间强劲激活、扩充、持续,并且在疾病复发 的情况下能够实现“记忆”反应。car t细胞较高且可管理的临床功效 目前仅限于bcma阳性和cd-19阳性b细胞恶性肿瘤以及表达hla
-ꢀ
a2的表达ny-eso-1-肽的滑膜肉瘤患者。对于改进基因工程化t细 胞以更广泛地对抗各种人类恶性肿瘤存在明确需求。本文描述了包括 cd3ε、cd3γ和cd3δ在内的tcr亚单位的新型融合蛋白,以及具有 对细胞表面抗原有特异性的结合结构域的tcrα和tcrβ链的新型融 合蛋白,其具有克服现有方法的局限性的潜力。本文描述了相比car 更有效地杀伤靶细胞,但释放相当或更低水平的促炎细胞因子的新型 融合蛋白。这些融合蛋白及其使用方法表现出t细胞受体(tcr)融 合蛋白(tfp)相对于car的优势,因为这些细胞因子升高的水平与 过继car-t疗法的剂量限制性毒性相关。
技术实现要素:
[0005]
本文提供了t细胞受体(tcr)融合蛋白(tfp)、被工程化为表 达一种或多种tfp的t细胞,及其用于治疗疾病的使用方法。
[0006]
在一个方面,本文提供了编码t细胞受体(tcr)融合蛋白(tfp) 的分离的重组核酸分子,所述tfp包含:tcr亚单位以及包含抗间皮素 结合结构域的人或人源化抗体结构域。
[0007]
在一个方面,本文提供了编码t细胞受体(tcr)融合蛋白(tfp) 的分离的重组核酸分子,所述tfp包含:tcr亚单位,所述tcr亚单 位包含tcr细胞外结构域的至少一部分和包含来自cd3ε的细胞内信号 传导结构域的刺激结构域的tcr细胞内结构域;以及包含抗原结合结构 域的人或人源化抗体结构域,其中所述tcr亚单位和所述抗体结构域可 操作地连接,并且其中所述tfp当在t细胞中表达时并入tcr中。
[0008]
在一个方面,本文提供了编码t细胞受体(tcr)融合蛋白(tfp) 的分离的重组核酸分子,所述tfp包含:tcr亚单位,所述tcr亚单 位包含tcr细胞外结构域的至少一部分和包含来自cd3γ的细胞内信 号传导结构域的刺激结构域的tcr细胞内结构域;以及包含抗原结合结 构域的人或人源化抗体结构域,其中所述tcr亚单位和所述抗体结构域 可操作地连接,并且其中所述tfp当在t细胞中表达时并入tcr中。
[0009]
在一个方面,本文提供了编码t细胞受体(tcr)融合蛋白(tfp) 的分离的重组核酸分子,所述tfp包含:tcr亚单位,所述tcr亚单 位包含tcr细胞外结构域的至少一部分和包含来自cd3δ的细胞内信 号传导结构域的刺激结构域的tcr细胞内结构域;以及包含抗原结合结 构域的人或人源化抗体结构域,其中所述tcr亚单位和所述抗体结构域 可操作地连接,并且其中所述tfp当在t细胞中表达时并入tcr中。
[0010]
在一个方面,本文提供了编码t细胞受体(tcr)融合蛋白(tfp) 的分离的重组核酸分子,所述tfp包含:tcr亚单位,所述tcr亚单 位包含tcr细胞外结构域的至少一部分和包含来自tcrα的细胞内信 号传导结构域的刺激结构域的tcr细胞内结构域;以及包含抗原结合结 构域的人或人源化抗体结构域,其中所述tcr亚单位和所述抗体结构域 可操作地连接,并且其中所述tfp当在t细胞中表达时并入tcr中。
[0011]
在一个方面,本文提供了编码t细胞受体(tcr)融合蛋白(tfp) 的分离的重组核酸分子,所述tfp包含:tcr亚单位,所述tcr亚单 位包含tcr细胞外结构域的至少一部分和包含来自tcrβ的细胞内信 号传导结构域的刺激结构域的tcr细胞内结构域;以及包含抗原结合结 构域的人或人源化抗体结构域,其中所述tcr亚单位和所述抗体结构域 可操作地连接,并且其中所述tfp当在t细胞中表达时并入tcr中。
[0012]
在一个方面,本文提供了编码t细胞受体(tcr)融合蛋白(tfp) 的分离的重组核酸分子,所述tfp包含:tcr亚单位以及包含抗原结合 结构域的人或人源化抗体结构域,所述抗原结合结构域是抗间皮素结合 结构域。
[0013]
在一些情况下,所述tcr亚单位和所述抗体结构域可操作地连接。 在一些情况下,所述tfp当在t细胞中表达时并入tcr中。在一些情 况下,所编码的抗原结合结构域通过连接体序列与所述tcr细胞外结构 域连接。在一些情况下,所编码的连接体序列包括(g4s)
n
,其中n=1至 4。在一些情况下,所述tcr亚单位包含tcr细胞外结构域。在一些情 况下,所述tcr亚单位包含tcr跨膜结构域。在一些情况下,所述tcr 亚单位包含tcr细胞内结构域。在一些情况下,所述tcr亚单位包含 (i)tcr细胞外结构域,(ii)tcr跨膜结构域,和(iii)tcr
细胞内 结构域,其中(i)、(ii)和(iii)中的至少两个来自相同的tcr亚单 位。在一些情况下,所述tcr亚单位包含tcr细胞内结构域,所述tcr 细胞内结构域包含选自cd3ε、cd3γ或cd3δ的细胞内信号传导结构域 的刺激结构域,或其具有至少一个、两个或三个修饰的氨基酸序列。在 一些情况下,所述tcr亚单位包含细胞内结构域,所述细胞内结构域包 含选自4-1bb的功能性信号传导结构域和/或cd3ζ的功能性信号传导 结构域的刺激结构域,或其具有至少一个修饰的氨基酸序列。在一些情 况下,所述人或人源化抗体结构域包含抗体片段。在一些情况下,所述 人或人源化抗体结构域包含scfv或v
h
结构域。在一些情况下,所述分 离的核酸分子编码(i)抗间皮素轻链结合结构域氨基酸序列的轻链(lc) cdr1、lc cdr2和lc cdr3,其分别与本文提供的抗间皮素轻链结合 结构域的轻链(lc)cdr1、lc cdr2和lc cdr3具有70-100%的序 列同一性,和/或(ii)抗间皮素重链结合结构域氨基酸序列的重链(hc) cdr1、hc cdr2和hc cdr3,其分别与本文提供的抗间皮素重链结合 结构域的重链(hc)cdr1、hc cdr2和hc cdr3具有70-100%的序 列同一性。在一些情况下,所述分离的核酸分子编码轻链可变区,其中 所述轻链可变区包含相对于本文提供的轻链可变区的轻链可变区氨基酸 序列具有至少一个但不多于30个修饰的氨基酸序列,或与本文提供的 轻链可变区的轻链可变区氨基酸序列具有95-99%同一性的序列。在一些 情况下,所述分离的核酸分子编码重链可变区,其中所述重链可变区包 含相对于本文提供的重链可变区的重链可变区氨基酸序列具有至少一个 但不多于30个修饰的氨基酸序列,或与本文提供的重链可变区的重链 可变区氨基酸序列具有95-99%同一性的序列。在一些情况下,所述tfp 包含tcr亚单位的细胞外结构域,所述细胞外结构域包括选自下组的蛋 白质的细胞外结构域或其部分:tcrα链、tcrβ链、cd3εtcr亚单位、 cd3γtcr亚单位、cd3δtcr亚单位、其功能片段及其具有至少一个 但不多于20个修饰的氨基酸序列。在一些情况下,所编码的tfp包含 跨膜结构域,所述跨膜结构域包括选自下组的蛋白质的跨膜结构域: tcrα链、tcrβ链、cd3εtcr亚单位、cd3γtcr亚单位、cd3δtcr 亚单位、其功能片段及其具有至少一个但不多于20个修饰的氨基酸序 列。在一些情况下,所编码的tfp包含跨膜结构域,所述跨膜结构域包 括选自下组的蛋白质的跨膜结构域:tcrα链、tcrβ链、tcrζ链、cd3ε tcr亚单位、cd3γtcr亚单位、cd3δtcr亚单位、cd45、cd2、cd4、 cd5、cd8、cd9、cd16、cd22、cd33、cd28、cd37、cd64、cd80、 cd86、cd134、cd137、cd154、其功能片段及其具有至少一个但不多 于20个修饰的氨基酸序列。在一些情况下,所述分离的核酸分子进一步 包含编码共刺激结构域的序列。在一些情况下,所述共刺激结构域是从 选自下组的蛋白质获得的功能性信号传导结构域:dap10、dap12、cd30、 light、ox40、cd2、cd27、cd28、cds、icam-1、lfa-1(cd11a/cd18)、 icos(cd278)和4-1bb(cd137)及其具有至少一个但不多于20个修 饰的氨基酸序列。在一些情况下,所述分离的核酸分子进一步包含前导 序列。在一些情况下,所述分离的核酸分子为mrna。
[0014]
在一些情况下,所述tfp包含tcr亚单位的基于免疫受体酪氨 酸的活化基序(itam),所述itam包括选自下组的蛋白质的itam或 其部分:cd3ζtcr亚单位、cd3εtcr亚单位、cd3γtcr亚单位、 cd3δtcr亚单位、tcrζ链、fcε受体1链、fcε受体2链、fcγ受体1 链、fcγ受体2a链、fcγ受体2b1链、fcγ受体2b2链、fcγ受体3a链、fcγ受体3b链、fcβ受体1链、tyrobp(dap12)、cd5、cd16a、 cd16b、cd22、cd23、cd32、cd64、cd79a、cd79b、cd89、cd278、 cd66d、其功能片段及其具有至少一个但不多于20个修饰的氨基酸序 列。在一些情况下,所
述itam取代cd3γ、cd3δ或cd3ε的itam。 在一些情况下,所述itam选自cd3ζtcr亚单位、cd3εtcr亚单位、 cd3γtcr亚单位和cd3δtcr亚单位,并取代选自cd3ζtcr亚单位、 cd3εtcr亚单位、cd3γtcr亚单位和cd3δtcr亚单位的不同itam。
[0015]
在一些情况下,所述核酸包含核苷酸类似物。在一些情况下,所 述核苷酸类似物选自2
’-
o-甲基、2
’-
o-甲氧基乙基(2
’-
o-moe)、2
’-
o
-ꢀ
氨基丙基、2
’-
脱氧、t-脱氧-2
’-
氟代、2
’-
o-氨基丙基(2
’-
o-ap)、2
’-
o
-ꢀ
二甲基氨基乙基(2
’-
o-dmaoe)、2
’-
o-二甲基氨基丙基(2
’-
o-dmap)、 t-o-二甲基氨基乙氧基乙基(2
’-
o-dmaeoe)、2
’-
o-n-甲基乙酰氨基 (2
’-
o-nma)修饰的、锁定核酸(lna)、乙烯核酸(ena)、肽核酸 (pna)、1
’
,5
’-
脱水己糖醇核酸(hna)、吗啉代、甲基膦酸酯核苷酸、 硫醇膦酸酯核苷酸和2
’-
氟代n3-p5
’-
亚磷酰胺。
[0016]
在一个方面,本文提供了由本文提供的核酸分子编码的分离的多 肽分子。
[0017]
在一个方面,本文提供了分离的tfp分子,其包含人或人源化抗 间皮素结合结构域、tcr细胞外结构域、跨膜结构域和细胞内结构域。
[0018]
在一个方面,本文提供了分离的tfp分子,其包含人或人源化抗 间皮素结合结构域、tcr细胞外结构域、跨膜结构域和细胞内信号传导 结构域,其中所述tfp分子能够与内源tcr复合物和/或至少一种内源 tcr多肽在功能上相互作用。
[0019]
在一些实施方案中,包含作为由所述核酸编码的抗间皮素结合结 构域的抗原结合结构域的所述人或人源化抗体结构域,或包含所述抗间 皮素结合结构域的抗体,或表达由所述核酸编码的抗间皮素结合结构域 的细胞,具有以下亲和力值:至多约200nm、100nm、75nm、50nm、 25nm、20nm、15nm、14nm、13nm、12nm、11nm、10nm、9nm、 8nm、7nm、6nm、5nm、4nm、3nm、2nm、1nm、0.9nm、0.8nm、0.7nm、0.6nm、0.5nm、0.4nm、0.3nm、0.2nm、0.1nm、0.09nm、 0.08nm、0.07nm、0.06nm、0.05nm、0.04nm、0.03nm、0.02nm或 0.01nm;和/或至少约100nm、75nm、50nm、25nm、20nm、15nm、 14nm、13nm、12nm、11nm、10nm、9nm、8nm、7nm、6nm、5 nm、4nm、3nm、2nm、1nm、0.9nm、0.8nm、0.7nm、0.6nm、0.5 nm、0.4nm、0.3nm、0.2nm、0.1nm、0.09nm、0.08nm、0.07nm、 0.06nm、0.05nm、0.04nm、0.03nm、0.02nm或0.01nm;和/或约200 nm、100nm、75nm、50nm、25nm、20nm、15nm、14nm、13nm、 12nm、11nm、10nm、9nm、8nm、7nm、6nm、5nm、4nm、3nm、 2nm、1nm、0.9nm、0.8nm、0.7nm、0.6nm、0.5nm、0.4nm、0.3 nm、0.2nm、0.1nm、0.09nm、0.08nm、0.07nm、0.06nm、0.05nm、 0.04nm、0.03nm、0.02nm或0.01nm。
[0020]
在一个方面,本文提供了分离的tfp分子,其包含人或人源化抗 间皮素结合结构域、tcr细胞外结构域、跨膜结构域和细胞内信号传导 结构域,其中所述tfp分子能够在功能上整合到内源tcr复合物中。
[0021]
在一些情况下,所述分离的tfp分子包含抗体或抗体片段,所述 抗体或抗体片段包含人或人源化抗间皮素结合结构域、tcr细胞外结构 域、跨膜结构域和细胞内结构域。在一些情况下,所述抗间皮素结合结 构域是scfv、v
h
结构域或骆驼科动物v
hh
结构域。在一些情况下,所述 抗间皮素结合结构域包含与本文提供的重链的氨基酸序列、其功能片段 或其具有至少一个但不多于30个修饰的氨基酸序列具有95-100%同一 性的重链。在一些情况下,所述抗间皮素结合结构域包含与本文提供的 轻链的氨基酸序列、其功能片段或其具有至少一个但不多于30个修饰 的氨基酸序列具有95-100%同一性的轻链。在一些情况下,所述分离的 tfp分子包含tcr细胞外结构域,所述tcr细胞外结构域包括选自下 组的蛋白质的细胞外结构域或其部分:tcrα链、tcrβ链、cd3εtcr 亚单位、cd3γtcr亚单位、cd3δtcr
亚单位、其功能片段及其具有至 少一个但不多于20个修饰的氨基酸序列。在一些情况下,所述抗间皮素 结合结构域通过连接体序列与所述tcr细胞外结构域连接。在一些情况 下,所述连接体区包括(g4s)
n
,其中n=1至4。
[0022]
在一些情况下,所述分离的tfp分子进一步包含编码共刺激结构 域的序列。在一些情况下,所述分离的tfp分子进一步包含编码细胞内 信号传导结构域的序列。在一些情况下,所述分离的tfp分子进一步包 含前导序列。
[0023]
在一个方面,本文提供了包含编码本文提供的tfp的核酸分子的 载体。在一些情况下,所述载体选自dna、rna、质粒、慢病毒载体、 腺病毒载体、劳斯肉瘤病毒(rsv)载体或逆转录病毒载体。在一些情 况下,所述载体进一步包含启动子。在一些情况下,所述载体是体外转 录的载体。在一些情况下,所述载体中的核酸序列进一步包含聚(a)尾。 在一些情况下,所述载体中的核酸序列进一步包含3
’
utr。
[0024]
在一个方面,本文提供了包含本文提供的载体的细胞。在一些情 况下,所述细胞是人t细胞。在一些情况下,所述t细胞是cd8+或 cd4+ t细胞。在一些情况下,所述t细胞是γδt细胞。在一些情况下, 所述t细胞是nk-t细胞。在一些情况下,所述细胞进一步包含编码抑 制性分子的核酸,所述抑制性分子包含第一多肽,所述第一多肽包含抑 制性分子的至少一部分,与包含来自细胞内信号传导结构域的正信号的 第二多肽缔合。在一些情况下,所述抑制性分子包含第一多肽和第二多 肽,所述第一多肽包含pd1的至少一部分,所述第二多肽包含共刺激结 构域和初级信号传导结构域。
[0025]
在一个方面,本文提供了包含至少两个tfp分子的人cd8+或 cd4+ t细胞,所述tfp分子包含人或人源化抗间皮素结合结构域、tcr 细胞外结构域、跨膜结构域和细胞内结构域,其中所述tfp分子能够与 所述人cd8+或cd4+ t细胞的表面之中、之处和/或之上的内源tcr复 合物和/或至少一种内源tcr多肽在功能上相互作用。
[0026]
在一个方面,本文提供了一种蛋白质复合物,其包含:tfp分子, 其包含人或人源化抗间皮素结合结构域、tcr细胞外结构域、跨膜结构 域和细胞内结构域;以及至少一种内源tcr复合物。
[0027]
在一些情况下,所述tcr包含选自下组的蛋白质的细胞外结构域 或其部分:tcrα链、tcrβ链、cd3εtcr亚单位、cd3γtcr亚单位 和cd3δtcr亚单位。在一些情况下,所述抗间皮素结合结构域通过连 接体序列与所述tcr细胞外结构域连接。在一些情况下,所述连接体区 包括(g4s)
n
,其中n=1至4。
[0028]
在一个方面,本文提供了人cd8+或cd4+ t细胞,其中每个本文 提供的蛋白质复合物包含至少两个不同的tfp蛋白。
[0029]
在一个方面,本文提供了制备细胞的方法,该方法包括用本文提 供的载体转导t细胞。
[0030]
在一个方面,本文提供了产生rna工程化细胞群体的方法,该方 法包括将体外转录的rna或合成rna引入细胞中,其中所述rna包 含编码本文提供的tfp分子的核酸。
[0031]
在一个方面,本文提供了在哺乳动物中提供抗肿瘤免疫的方法, 该方法包括向所述哺乳动物施用有效量的表达本文提供的tfp分子或 表达本文提供的多肽分子的细胞。
[0032]
在一些情况下,所述细胞是自体t细胞。在一些情况下,所述细 胞是同种异体t细胞。在一些情况下,所述哺乳动物是人。
[0033]
在一个方面,本文提供了治疗患有与间皮素表达相关的疾病的哺 乳动物的方法,该方法包括向所述哺乳动物施用有效量的本文提供的 tfp分子、本文提供的细胞或本文提供的多肽分子。在一些情况下,所 述与间皮素表达相关的疾病选自增生性疾病、癌症、恶性肿瘤和与间皮 素表达相关的非癌症相关适应症。在一些情况下,所述疾病是选自间皮 瘤、肾细胞癌、胃癌、乳腺癌、肺癌、卵巢癌、前列腺癌、结肠癌、宫 颈癌、脑癌、肝癌、胰腺癌、甲状腺癌、膀胱癌、输尿管癌、肾癌、子 宫内膜癌、食管癌、胃癌、胸腺癌、胆管癌和胃癌的癌症。
[0034]
在一些情况下,所述疾病是癌症。在一些情况下,所述疾病选自 间皮瘤、乳头状浆液性卵巢腺癌、透明细胞卵巢癌、混合型米勒氏卵巢 癌(mullerian ovarian carcinoma)、子宫内膜粘液性卵巢癌、恶性胸膜疾 病、胰腺腺癌、胰腺导管腺癌、子宫浆液性癌、肺腺癌、肝外胆管癌、 胃腺癌、食管腺癌、结直肠腺癌、乳腺腺癌、与间皮素表达相关的疾病、 与间皮素表达相关的疾病、非粘液性卵巢癌、侵袭性导管腺癌、肺腺癌、 胃/食管腺癌、结直肠腺癌、白血病、小儿急性髓样白血病、侵袭性导管 内乳头状粘液性肿瘤(ipmn)、子宫内膜腺癌、胃/食管腺癌、肺腺癌、 乳腺腺癌及其组合。
[0035]
在一些情况下,所述表达tfp分子的细胞与提高表达tfp分子的 细胞的功效的药剂联合施用。在一些情况下,与施用有效量的表达抗间 皮素嵌合抗原受体(car)的t细胞的哺乳动物相比,所述哺乳动物中 释放的细胞因子较少。在一些情况下,所述表达tfp分子的细胞与改善 与施用表达tfp分子的细胞相关的一种或多种副作用的药剂联合施用。 在一些情况下,所述表达tfp分子的细胞与治疗与间皮素相关的疾病的 药剂联合施用。
[0036]
在一个方面,本文提供的分离的核酸分子、本文提供的分离的多 肽分子、本文提供的分离的tfp、本文提供的复合物、本文提供的载体 或本文提供的细胞用作药物。
[0037]
在一个方面,本文提供了治疗患有与间皮素表达相关的疾病的哺 乳动物的方法,该方法包括向所述哺乳动物施用有效量的本文提供的 tfp分子、本文提供的细胞或本文提供的多肽分子,其中与施用有效量 的表达抗间皮素嵌合抗原受体(car)的t细胞的哺乳动物相比,所述 哺乳动物中释放的细胞因子较少。援引并入
[0038]
本说明书中提到的所有出版物、专利和专利申请均通过引用并入 本文,其程度如同特别地且单独地指出每一个单独的出版物、专利或专 利申请均通过引用而并入。
附图说明
[0039]
本发明的新颖特征在所附的权利要求书中具体阐述。通过参考以 下对利用本发明原理的说明性实施方式加以阐述的详细描述以及附图, 将会获得对本发明的特征和优点的更好的理解,在这些附图中:
[0040]
图1是展示了本发明的t细胞受体融合多肽(tfp)的使用的 示意图。示例性tfp含有经由(g4s)3连接体序列融合的抗间皮素scfv 和全长cd3ε多肽。当由t细胞产生或引入t细胞时,tfp与内源t 细胞受体(tcr)的其他多肽缔合(显示包括两个cd3ε多肽、一个 cd3γ多肽、一个cd3δ多肽、两个cd3ζ多肽、一个tcrα亚单位和 一个tcrβ亚单位,其中水平灰色段代表质膜)以形成重编程的tcr, 其中一个或两个内源cd3ε多肽被tfp置换。
[0041]
图2表示展示了本发明的重编程t细胞受体融合多肽(tfp) 的示例性变化的示意
图。表示为scfv:tcr-vα的图示示出了含有tfp 的示例性重编程tcr,该tfp含有经由(g4s)3连接体序列融合的抗间 皮素scfv和全长tcr-vα多肽。表示为scfv:tcr-vα:tcr-vβ的图 示示出了含有多个tfp的示例性重编程tcr,该tfp包含i)经由 (g4s)3连接体序列融合的抗间皮素scfv和全长tcr-vα多肽,和ii) 经由(g4s)3连接体序列融合的抗间皮素scfv和全长tcr-vβ多肽。表 示为scfv:δtcr-vα:cd3ε的图示示出了含有多个tfp的示例性重编 程tcr,该tfp包含i)经由(g4s)3连接体序列融合的抗间皮素scfv 和截短的(δ)tcr多肽,和ii)经由(g4s)3连接体序列融合的抗间皮素 scfv和全长cd3ε多肽。截短的(δ)tcr多肽通过缺失vα而截短。表 示为scfv:δtcr-vα:δtcr-vβ的图示示出了含有多个tfp的示例性 重编程tcr,该tfp包含i)经由(g4s)3连接体序列融合的抗间皮素 scfv和截短的(δ)tcr vα多肽,和ii)经由(g4s)3连接体序列融合的 抗间皮素scfv和截短的(δ)tcr vβ多肽。截短的(δ)tcr多肽通过缺 失vβ而截短。
[0042]
图3是展示了本发明的t细胞受体融合多肽(tfp)的使用的 示意图。示例性tfp含有经由(g4s)3连接体序列融合的抗间皮素v
h
结构域和全长cd3ε多肽。当由t细胞产生或引入t细胞时,tfp与 内源t细胞受体(tcr)的其他多肽缔合(显示包括两个cd3ε多肽、 一个cd3γ多肽、一个cd3δ多肽、两个cd3ζ多肽、一个tcrα亚 单位和一个tcrβ亚单位,其中水平灰色段代表质膜)以形成重编程 的tcr,其中一个或两个内源cd3ε多肽被tfp置换。
[0043]
图4是展示了编码各种tfp的dna构建体的一系列示意图。
[0044]
图5a描绘了活化的pbmc细胞上tfp的示例性表面表达分 析,并显示了用msln tfp激活并针对cd8(抗cd8 apccy7,y轴) 和间皮素(“msln”)(r-藻红蛋白标记的hmsln igg,x轴) 进行染色的cd3
+
细胞(抗cd3 apc,门(gate))。从左到右显示了 未转导的或用抗msln-cd3εtfp、抗msln-cd28ζcar和抗msln
-ꢀ
41bbζcar构建体转导的细胞。
[0045]
图5b描绘了活化的pbmc细胞上tfp的示例性表面表达分 析,并显示了用内部单结构域tfp激活并针对msln fc进行染色并 针对gfp进行分析的细胞。第一行显示(从左到右)未转导的细胞和 用对照抗msln-cd3εtfp(“ss1”)转导的细胞。第2-4行分别显示 用gfp标记的(从左到右)cd3εtfp、cd3γtfp、tcrβtfp和cd28ζ car构建体转导的细胞中的抗msln结合物sd1、sd4和sd6。
[0046]
图6a是描绘抗msln-tfp构建体随时间对间皮素(msln) 阳性hela(宫颈腺癌,ccl-2
tm
)靶细胞的杀伤的示例性图 示。活化的pbmc未经处理(迹线#1)、未转导(迹线#2)或者用空 载体(迹线#3)、抗msln-cd3εtfp(迹线#4)、抗msln-cd28ζ car或抗msln-41bbζcar转导,并扩充8天,然后与1x104个 msln阳性hela靶细胞一起温育。
[0047]
图6b是描绘抗msln-tfp构建体随时间对msln阴性hela (宫颈腺癌,ccl-2
tm
)靶细胞的杀伤的示例性图示。活化的 pbmc未经处理(迹线#1)、未转导(迹线#2)或者用空载体(迹线 #3)、抗msln-cd3εtfp(迹线#4)、抗msln-cd28ζcar或抗 msln-41bbζcar转导,并扩充8天,然后与1x104个msln阳性 hela靶细胞一起温育。
[0048]
图6c示出了使用来自两个不同人供体(顶部和底部)的t细 胞在高msln表达细胞系(hela细胞)中对msln阳性细胞的杀伤。 示出了具有内部抗msln结合物sd1(图7a)、sd4(中)和sd6 (右)的tfp t细胞的细胞杀伤迹线。活化的pbmc未经转导(迹线 #1),或用cd3εtfp(迹线#2)、cd3γtfp(迹线#3)、tcrβtfp (迹线#4)或cd28ζcar转导。在实时细胞分析仪
(rtca)测定中 确定指示细胞毒性的归一化细胞指数。
[0049]
图7是显示出抗msln car t细胞和tfp t细胞针对表达高 水平间皮素的靶细胞系(hela-luc(
msln高
))的结合活性的一系列图示。 显示出在无t细胞(“仅靶标”)、空载体转导的(“nt”)、抗msln (阳性对照)或者具有内部抗间皮素结合物sd1(图7a)、sd4(图 7b)和sd6(图7c)的抗间皮素tfp t细胞的样品中的%细胞杀伤, 其各自为cd3εtfp、cd3γtfp、tcrβtfp和cd28ζcar的形式。 在每个图中,黑色条代表t细胞与靶细胞的比率为1:1,灰色条代表 t细胞与靶细胞的比率为1:5。对于第二t细胞供体观察到类似的结 果。
[0050]
图8是显示出抗msln car t细胞和tfp t细胞针对表达低 水平间皮素的靶细胞系(pc3-msln(-/低
))的活性的一系列图示。显示 出在无t细胞(“仅靶标”)、空载体转导的(“nt”)、抗msln(阳 性对照,“ss1”)或者tfp形式为cd3ε(图8a)、cd3γ(图8b)、 tcrβ(图8c)和cd28ζcar(图8d)的内部抗间皮素构建体sd1、 sd4和sd6的样品中的%细胞杀伤。在每个图中,黑色条代表t细胞 与靶细胞的比率为1:1,灰色条代表t细胞与靶细胞的比率为1:5。对 于第二t细胞供体观察到类似的结果。
[0051]
图9显示了facs分析的结果,证明表达抗msln car和tfp 构建体的t细胞当与msln+细胞共培养时的活化。如图9a所示, 从左到右,t细胞为未转导的、用空载体转导的、用抗msln-cd3ε tfp、抗msln-28ζcar或抗msln-41bbζcar转导的。与msln
-ꢀ
细胞共培养的细胞显示在上面一行,与msln+靶细胞共培养的细胞 显示在下面一行。然后用对表面活化标志物cd69和cd25或细胞溶 解颗粒组分粒酶b(grb)具有特异性的抗体对细胞进行染色。用抗 cd69染色的细胞数对应于x轴,用抗cd25染色的细胞数对应于y 轴。如图所示,相对于与msln-细胞共培养,通过与msln+细胞培 养激活了表达抗间皮素car和tfp构建体的t细胞,如通过升高的 cd69和cd25表达水平所证明的(图9b)。显示了msln-(白色 条)和msln+(黑色条)细胞中每种构建体的cd25+细胞的百分比。 使用k562 msln-细胞(圆形)和k562-msln+细胞(正方形)在未 转导的t细胞或用抗msln阳性对照结合物(“510-ss1-cd3ε”)转导 的t细胞中进行类似的实验(图9c)。数据表示cd25+、cd69+和 cd25+/cd69+细胞的总和。在图9d中,显示了与具有tfp形式cd3ε、 cd3γ、tcrβ和cd28ζcar的供体t细胞组合的k562 msln-靶细 胞(左图)和k562 msln+细胞(右图)中内部抗msln结合物sd1 (正方形)、sd4(圆形)和sd6(三角形)的数据。使用来自第二 t细胞供体的细胞观察到类似的结果。
[0052]
图10显示了facs分析的结果,证明表达抗msln car和 tfp构建体的t细胞当与msln+细胞共培养时的活化。用抗cd3 apc(门)和抗cd8 apccy7(y轴)针对表面抗原对细胞进行染色, 然后固定、透化,并用抗粒酶b alexafluor700(x轴)进行染色。如 图10a所示,从左到右,t细胞为未转导的、用空载体转导的、用抗 msln-cd3εtfp、抗msln-28ζcar或抗msln-41bbζcar转导 的。与msln-细胞共培养的细胞显示在上面一行,与msln+靶细胞 共培养的细胞显示在下面一行。相对于与msln-细胞共培养,通过与 msln+细胞培养激活了表达抗间皮素car和tfp构建体的cd8 t 细胞,如通过升高的细胞内grb水平所示(图10b)。显示了与msln
-ꢀ
(白色条)或msln+(黑色条)细胞共培养时每种构建体的粒酶b (“grb+”)细胞的百分比。
[0053]
图11显示了当与过表达msln的k562细胞共培养时,表达抗 msln car和tfp构建体的活化t细胞中细胞因子产生的elisa分 析结果。使用过表达bcma的k562细胞作为阴性对照。24小时后, 通过elisa分析细胞的ifn-γ(图11a)和il-2(图11b)表达。在 每个图中,从
左到右,t细胞为未转导的、用空载体转导的、用抗msln-cd3εtfp、抗msln-28ζcar或抗msln-41bbζcar转导 的。与msln-细胞共培养的细胞用白色条表示,与msln+靶细胞共 培养的细胞用黑色条表示。
[0054]
图12是显示出msln特异性sdab tfp t细胞在间皮瘤异种移 植小鼠模型中的体内功效的一系列图示。用萤光素酶标记的msto
-ꢀ
211h-fl-msln-luc以每只小鼠1x106个细胞对小鼠进行接种,并使 肿瘤生长到直至肿瘤体积约为300mm3,将1x107个t细胞静脉内注 射到每只动物中。图12a显示了注射t细胞后的肿瘤体积,该t细 胞从左到右包括无t细胞对照、sd1 cd3ε-tfp和sd4 cd3ε-tfp。 图12b显示了具有sd1和sd4结合物以及sd1 cd28ζcar的cd3γ
-ꢀ
tfp。图12c-图12d显示了来自图12a-图12b的存活小鼠的结果, 该小鼠用肿瘤细胞再次攻击以确定小鼠在没有第二次t细胞注射的 情况下是否会维持其抗msln免疫。已施用sd1 cd3ε-tfp t细胞 (图12c)和sd1 cd3γ-tfp t细胞(图12d)并且已预先清除肿瘤 的小鼠用msln+(msto)或msln-(raji)肿瘤细胞系重新接种。 测量肿瘤体积并示于x轴上。
[0055]
图13示出了来自卵巢癌患者的msln-tfp t细胞的产生和功 能分析。图13a是实验设计的示意图。图13b-图13c显示患者的 sd1ε-tfp t细胞对msto-msln-luc肿瘤细胞的体外杀伤。确认 msto-msln-luc肿瘤细胞(靶细胞)的间皮素表达(图13b);sd1ε
-ꢀ
tfp t细胞(效应细胞)和匹配的未转导对照以5:1、1:1或1:5的e:t (效应物比靶标)比添加,持续24小时。通过m5读板 仪(molecular devices)测量靶细胞的发光的相对发光单位(rlu)。 图中的每条线代表3次重复的平均值(图13c)。图13d-图13l显示 了来自卵巢癌患者的sd1ε-tfp t细胞的细胞因子谱的测量,包括 ifnγ(图13d)、gm-csf(图13e)、粒酶a(图13f)、粒酶b(图 13g)、il-2(图13h)、mip-1α(图13i)、mip-1β(图13j)、tnfα (图13k)和穿孔蛋白(图13l)。将msto-msln-luc肿瘤细胞(靶 细胞)以10000个细胞/孔接种在96孔平底板中。以1:1的比率添加sd1ε-tfp t细胞(效应细胞)和匹配的未转导对照,持续24小时。 收集细胞上清液并使用测定来测量细胞因子。
[0056]
图14显示了来源于患者的sd1 cd3ε-tfp t细胞在msln-高 异种移植肿瘤小鼠模型中的体内功效。以每只小鼠1x106个细胞皮下 接种msto-211h-fl msln-luc细胞。肿瘤注射后10天(肿瘤体积 ~200-300mm3),将5x106个t细胞静脉内注射到每只动物中。图中 的每条线代表一只动物。显示来自nd12(图14a)、患者1(图14b)、 患者2(图14c)、患者3(图14d)和患者4(图14e)的t细胞的 数据。圆形表示用未转导的t细胞接种的小鼠的肿瘤大小;正方形表 示接种tfp t细胞的小鼠的肿瘤大小。
具体实施方式
[0057]
在一个方面,本文描述了编码t细胞受体(tcr)融合蛋白(tfp) 的分离的核酸分子,该tfp包含tcr亚单位以及包含抗间皮素结合结 构域的人或人源化抗体结构域。在一些实施方案中,该tcr亚单位包含 tcr细胞外结构域。在其他实施方案中,该tcr亚单位包含tcr跨膜 结构域。在其他实施方案中,该tcr亚单位包含tcr细胞内结构域。 在其他实施方案中,该tcr亚单位包含(i)tcr细胞外结构域,(ii) tcr跨膜结构域,和(iii)tcr细胞内结构域,其中(i)、(ii)和(iii) 中的至少两个来自相同的tcr亚单位。在其他实施方案中,该tcr亚 单位包含tcr细胞内结构域,该细胞内结构域包含选自cd3ε、cd3γ或 cd3δ的细胞内信号传导结构域的刺激结构域,或其具有至少一个、两个 或三个修饰的氨基酸序列。在其
他实施方案中,该tcr亚单位包含细胞 内结构域,该细胞内结构域包含选自4-1bb的功能性信号传导结构域和 /或cd3ζ的功能性信号传导结构域的刺激结构域,或其具有至少一个、 两个或三个修饰的氨基酸序列。
[0058]
在一些实施方案中,所述分离的核酸分子包含(i)本文提供的任 何抗间皮素轻链结合结构域氨基酸序列的轻链(lc)cdr1、lc cdr2 和lc cdr3,和/或(ii)本文提供的任何抗间皮素重链结合结构域氨基 酸序列的重链(hc)cdr1,hc cdr2和hc cdr3。
[0059]
在一些实施方案中,所述轻链可变区包含相对于本文提供的轻链 可变区的氨基酸序列具有至少一个、两个或三个修饰但不多于30、20或 10个修饰的氨基酸序列,或与本文提供的氨基酸序列具有95-99%同一 性的序列。在其他实施方案中,所述重链可变区包含相对于本文提供的 重链可变区的氨基酸序列具有至少一个、两个或三个修饰但不多于30、 20或10个修饰的氨基酸序列,或与本文提供的氨基酸序列具有95-99% 同一性的序列。
[0060]
在一些实施方案中,所述tfp包含tcr亚单位的细胞外结构域, 该细胞外结构域包括选自下组的蛋白质的细胞外结构域或其部分:t细 胞受体的α或β链、cd3δ、cd3ε或cd3γ,或其功能片段,或其具有至 少一个、两个或三个修饰但不多于20、10或5个修饰的氨基酸序列。在 其他实施方案中,所编码的tfp包含跨膜结构域,该跨膜结构域包括选 自下组的蛋白质的跨膜结构域:tcr的α链、β链,或tcr亚单位cd3ε、 cd3γ和cd3δ,或其功能片段,或其具有至少一个、两个或三个修饰但 不多于20、10或5个修饰的氨基酸序列。
[0061]
在一些实施方案中,所编码的tfp包含跨膜结构域,该跨膜结构 域包括选自下组的蛋白质的跨膜结构域:tcr的α链、β链或ζ链,或 cd3ε、cd3γ和cd3δ、cd45、cd2、cd4、cd5、cd8、cd9、cd16、 cd22、cd33、cd28、cd37、cd64、cd80、cd86、cd134、cd137和 cd154,或其功能片段,或其具有至少一个、两个或三个修饰但不多于 20、10或5个修饰的氨基酸序列。
[0062]
在一些实施方案中,所编码的抗间皮素结合结构域通过连接体序 列与所述tcr细胞外结构域连接。在一些情况下,所编码的连接体序列 包括(g4s)
n
,其中n=1至4。在一些情况下,所编码的连接体序列包括长 连接体(ll)序列。在一些情况下,所编码的长连接体序列包括(g4s)
n
, 其中n=2至4。在一些情况下,所编码的连接体序列包括短连接体(sl) 序列。在一些情况下,所编码的短连接体序列包括(g4s)
n
,其中n=1至 3。
[0063]
在一些实施方案中,所述分离的核酸分子进一步包含编码共刺激 结构域的序列。在一些情况下,该共刺激结构域是从选自下组的蛋白质 获得的功能性信号传导结构域:dap10、dap12、cd30、light、ox40、 cd2、cd27、cd28、cds、icam-1、lfa-1(cd11a/cd18)、icos(cd278) 和4-1bb(cd137),或其具有至少一个、两个或三个修饰但不多于20、 10或5个修饰的氨基酸序列。
[0064]
在一些实施方案中,所述分离的核酸分子进一步包含前导序列。
[0065]
本文还提供了由任何前述核酸分子编码的分离的多肽分子。
[0066]
在另一个方面,本文还提供了分离的t细胞受体融合蛋白(tfp) 分子,其包含人或人源化抗间皮素结合结构域、tcr细胞外结构域、跨 膜结构域和细胞内结构域。在一些实施方案中,所述分离的tfp分子包 含抗体或抗体片段,该抗体或抗体片段包含人或人源化抗间皮素结合结 构域、tcr细胞外结构域、跨膜结构域和细胞内结构域。
[0067]
在一些实施方案中,所述人或人源化抗体结构域包含抗体片段。 在一些实施方案
中,所述人或人源化抗体结构域包含scfv或v
h
结构域。
[0068]
在一些实施方案中,所述抗间皮素结合结构域是scfv或v
h
结构 域。在其他实施方案中,所述抗间皮素结合结构域包含以下序列的轻链 和重链:本文提供的氨基酸序列,或其功能片段,或相对于本文提供的 轻链可变区的氨基酸序列具有至少一个、两个或三个修饰但不多于30、 20或10个修饰的氨基酸序列,或与本文提供的氨基酸序列具有95-99% 同一性的序列。
[0069]
在一些实施方案中,所述分离的tfp分子包含tcr细胞外结构 域,该细胞外结构域包括选自下组的蛋白质的细胞外结构域或其部分: t细胞受体的α或β链、cd3δ、cd3ε或cd3γ,或其具有至少一个、两 个或三个修饰但不多于20、10或5个修饰的氨基酸序列。
[0070]
在一些实施方案中,所述抗间皮素结合结构域通过连接体序列与 所述tcr细胞外结构域连接。在一些情况下,连接体区包括(g4s)
n
,其 中n=1至4。在一些情况下,该连接体序列包括长连接体(ll)序列。 在一些情况下,该长连接体序列包括(g4s)
n
,其中n=2至4。在一些情况 下,该连接体序列包括短连接体(sl)序列。在一些情况下,该短连接 体序列包括(g4s)
n
,其中n=1至3。
[0071]
在一些实施方案中,所述分离的tfp分子进一步包含编码共刺激 结构域的序列。在其他实施方案中,所述分离的tfp分子进一步包含编 码细胞内信号传导结构域的序列。在其他实施方案中,所述分离的tfp 分子进一步包含前导序列。
[0072]
本文还提供了包含编码任何前述tfp分子的核酸分子的载体。在 一些实施方案中,该载体选自dna、rna、质粒、慢病毒载体、腺病毒 载体或逆转录病毒载体。在一些实施方案中,该载体进一步包含启动子。 在一些实施方案中,该载体是体外转录的载体。在一些实施方案中,该 载体中的核酸序列进一步包含聚(a)尾。在一些实施方案中,该载体中的 核酸序列进一步包含3
’
utr。
[0073]
本文还提供了包含任何所述载体的细胞。在一些实施方案中,该 细胞是人t细胞。在一些实施方案中,该细胞是cd8+或cd4+ t细胞。 在一些实施方案中,该细胞进一步包含编码抑制性分子的核酸,该抑制 性分子包含与第二多肽缔合的第一多肽,第一多肽包含抑制性分子的至 少一部分,第二多肽包含来自细胞内信号传导结构域的正信号。在一些 情况下,该抑制性分子包含第一多肽和第二多肽,第一多肽包含pd1的 至少一部分,第二多肽包含共刺激结构域和初级信号传导结构域。
[0074]
在另一个方面,本文提供了分离的tfp分子,其包含人或人源化 抗间皮素结合结构域、tcr细胞外结构域、跨膜结构域和细胞内信号传 导结构域,其中该tfp分子能够与内源tcr复合物和/或至少一种内源 tcr多肽在功能上相互作用。
[0075]
在另一个方面,本文提供了分离的tfp分子,其包含人或人源化 抗间皮素结合结构域、tcr细胞外结构域、跨膜结构域和细胞内信号传 导结构域,其中该tfp分子能够在功能上整合到内源tcr复合物中。
[0076]
在另一个方面,本文提供了包含至少两个tfp分子的人cd8+或 cd4+ t细胞,该tfp分子包含人或人源化抗间皮素结合结构域、tcr 细胞外结构域、跨膜结构域和细胞内结构域,其中该tfp分子能够与该 人cd8+或cd4+ t细胞的表面之中、之处和/或之上的内源tcr复合物 和/或至少一种内源tcr多肽在功能上相互作用。
[0077]
在另一个方面,本文提供了蛋白质复合物,其包含i)tfp分子, 其包含人或人源化
抗间皮素结合结构域、tcr细胞外结构域、跨膜结构 域和细胞内结构域;以及ii)至少一种内源tcr复合物。
[0078]
在一些实施方案中,所述tcr包含选自下组的蛋白质的细胞外结 构域或其部分:t细胞受体的α或β链、cd3δ、cd3ε或cd3γ。在一些 实施方案中,抗间皮素结合结构域通过连接体序列与所述tcr细胞外结 构域连接。在一些情况下,连接体区包括(g4s)
n
,其中n=1至4。在一些 情况下,该连接体序列包括长连接体(ll)序列。在一些情况下,该长 连接体序列包括(g4s)
n
,其中n=2至4。在一些情况下,该连接体序列包 括短连接体(sl)序列。在一些情况下,该短连接体序列包括(g4s)
n
,其 中n=1至3。
[0079]
本文还提供了人cd8+或cd4+ t细胞,其中每个任何所述的蛋白 质复合物包含至少两个不同的tfp蛋白。
[0080]
在另一个方面,本文提供了人cd8+或cd4+ t细胞群体,其中该 群体的t细胞单独或共同地包含至少两个tfp分子,该tfp分子包含 人或人源化抗间皮素结合结构域、tcr细胞外结构域、跨膜结构域和细 胞内结构域,其中该tfp分子能够与该人cd8+或cd4+ t细胞的表面 之中、之处和/或之上的内源tcr复合物和/或至少一种内源tcr多肽 在功能上相互作用。
[0081]
在另一个方面,本文提供了人cd8+或cd4+ t细胞群体,其中该 群体的t细胞单独或共同地包含至少两个由本文提供的分离的核酸分子 编码的tfp分子。
[0082]
在另一个方面,本文提供了制备细胞的方法,其包括用任何所述 载体转导t细胞。
[0083]
在另一个方面,本文提供了产生rna工程化细胞群体的方法,其 包括将体外转录的rna或合成rna引入细胞中,其中该rna包含编 码任何所述tfp分子的核酸。
[0084]
在另一个方面,本文提供了在哺乳动物中提供抗肿瘤免疫的方法, 其包括向哺乳动物施用有效量的表达任何所述tfp分子的细胞。在一些 实施方案中,该细胞是自体t细胞。在一些实施方案中,该细胞是同种 异体t细胞。在一些实施方案中,该哺乳动物是人。
[0085]
在另一个方面,本文提供了治疗患有与间皮素表达相关的疾病的 哺乳动物的方法,其包括向哺乳动物施用有效量的包含任何所述tfp分 子的细胞。在一些实施方案中,与间皮素表达相关的疾病选自增生性疾 病,诸如癌症或恶性肿瘤或癌前病况,诸如胰腺癌、卵巢癌、胃癌、肺 癌或子宫内膜癌,或是与间皮素表达相关的非癌症相关适应症。
[0086]
在一些实施方案中,表达任何所述tfp分子的细胞与改善与施用 表达tfp分子的细胞相关的一种或多种副作用的药剂联合施用。在一些 实施方案中,表达任何所述tfp分子的细胞与治疗与间皮素相关的疾病 的药剂联合施用。
[0087]
本文还提供了用作药物的任何所述分离的核酸分子、任何所述分 离的多肽分子、任何所述分离的tfp、任何所述蛋白质复合物、任何所 述载体或任何所述细胞。定义
[0088]
除非另有定义,否则本文使用的所有技术和科学术语具有与本发 明所属领域的普通技术人员所一般理解的相同的含义。
[0089]
术语“一个”和“一种”是指一个或多于一个(即,至少一个)该物 品的语法对象。举例而言,“一种元素”意指一种元素或多于一种元素。
[0090]
如本文所用,“约”可表示取决于具体情况并且由本领域技术人 员已知或可知的,加或减小于1%或1%、2%、3%、4%、5%、6%、 7%、8%、9%、10%、11%、12%、13%、14%、
15%、16%、17%、18%、 19%、20%、25%、30%或大于30%。
[0091]
如本说明书中所用,“受试者”或“个体”可包括但不限于哺乳动 物如人或非人哺乳动物,例如,家养的、农业的或野生的动物,以及 鸟类和水生动物。“患者”是患有疾病、病症或病况或者有患疾病、病 症或病况的风险或者在其他方面需要本文提供的组合物和方法的受 试者。
[0092]
如本文所用,“治疗”或“处理”是指治疗或改善疾病或病况的任 何成功标志。治疗可包括例如减少、延迟或缓解疾病或病况的一种或 多种症状的严重程度,或者其可包括降低患者经历疾病、缺陷、病症 或不良病况等的症状的频率。如本文所用,“治疗或预防”在本文中有 时用于指示导致疾病或病况的某种程度的治疗或改善的方法,并考虑 到针对该目的的一系列结果,包括但不限于完全预防该病况。
[0093]
如本文所用,“预防”是指预防患者中的疾病或病况,例如肿瘤 形成。例如,如果具有患肿瘤或其他形式的癌症的风险的个体用本发 明的方法治疗并且后来不患该肿瘤或其他形式的癌症,则该疾病至少 在一段时间内在该个体中得以预防。
[0094]
如本文所用,“治疗有效量”是足以对施用组合物的个体提供有 益效果或以其他方式减少有害的非有益事件的组合物或其活性组分 的量。本文的“治疗有效剂量”意指针对施用产生一种或多种所需或期 望(例如,有益)效果的剂量,这样的施用在给定的时间段内进行一 次或多次。确切的剂量将取决于治疗的目的,并且将可由本领域技术 人员使用已知技术确定(参见例如lieberman,pharmaceutical dosageforms(第1-3卷,1992);lloyd,the art,science and technology ofpharmaceutical compounding(1999);以及pickar,dosage calculations (1999))。
[0095]
如本文所用,“t细胞受体(tcr)融合蛋白”或“tfp”包括衍生 自各种包含tcr的多肽的重组多肽,其通常能够i)与靶细胞上的表 面抗原结合,并且ii)与完整tcr复合物的其他多肽组分相互作用, 通常在共同位于t细胞表面之中或之上时。“tfp t细胞”是已被转导 (例如,根据本文公开的方法)和表达tfp的t细胞,例如引入天然 tcr中。在一些实施方案中,该t细胞是cd4+ t细胞、cd8+ t细 胞或cd4+/cd8+ t细胞。在一些实施方案中,该tfp t细胞是nk 细胞。在一些实施方案中,该tfp t细胞是γδt细胞。
[0096]
如本文所用,术语“间皮素”也称msln或cak1抗原或前幼巨 核细胞(pre-pro-megakaryocyte)增强因子,是指人类中由msln(或 巨核细胞增强因子(mpf))基因编码的蛋白质。间皮素是存在于正 常间皮细胞上的40kda蛋白质,并且在包括间皮瘤和卵巢癌和胰腺 腺癌在内的几种人类肿瘤中过表达。间皮素基因编码前体蛋白,该前 体蛋白被加工以产生间皮素,间皮素通过糖基磷脂酰肌醇连接和被称 为巨核细胞增强因子(mpf)的31kda脱落片段附着于细胞膜。间皮 素可能参与细胞粘附,但其生物学功能尚不清楚。间皮素是肿瘤分化 抗原,其通常存在于胸膜、腹膜和心包膜的内衬间皮细胞上。间皮素 是在间皮瘤细胞、卵巢癌细胞、胰腺癌细胞和一些鳞状细胞癌上可检 测的抗原决定簇(参见例如,kojima等人,j.biol.chem.270:21984
-ꢀ
21990(1995)和onda等人,clin.cancer res.12:4225-4231(2006))。 间皮素与ca125/muc16相互作用(参见例如,rump等人,j.biol. chem.279:9190-9198(2004)和ma等人,j.biol.chem.287:33123
-ꢀ
33131(2012))。
[0097]
人和鼠氨基酸和核酸序列可见于公共数据库,诸如genbank、 uniprot和swiss-prot。例如,人间皮素的氨基酸序列可作为 uniprot/swiss-prot登录号q13421找到。人间
皮素多肽规范序列是 uniprot登录号q13421(或q13421-1): malptarpllgscgtpalgsllfllfslgwvqpsrtlagetgqeaa pldgvlanppnisslsprqllgfpcaevsglstervrelavalaqk nvklsteqlrclahrlseppedldalpldlllflnpdafsgpqact rffsritkanvdllprgaperqrllpaalacwgvrgsllseadvr algglacdlpgrfvaesaevllprlvscpgpldqdqqeaaraalq gggppygppstwsvstmdalrgllpvlgqpiirsipqgivaawrqr ssrdpswrqpertilrprfrrevektacpsgkkareideslifykkw eleacvdaallatqmdrvnaipftyeqldvlkhkldelypqgype sviqhlgylflkmspedirkwnvtsletlkallevnkghemspqa prrplpqvatlidrfvkgrgqldkdtldtltafypgylcslspeels svppssiwavrpqdldtcdprqldvlypkarlafqnmngseyfvk iqsflggaptedlkalsqqnvsmdlatfmklrtdavlpltvaevq kllgphveglkaeerhrpvrdwilrqrqddldtlglglqggipng ylvldlsmqealsgtpcllgpgpvltvlalllastla(seq id no: 15)。
[0098]
编码人间皮素转录物变体1的核苷酸序列可以以登录号 nm005823找到。编码人间皮素转录物变体2的核苷酸序列可以以登 录号nm013404找到。编码人间皮素转录物变体3的核苷酸序列可以 以登录号nm001177355找到。间皮素在间皮瘤细胞、卵巢癌细胞、 胰腺腺癌细胞和鳞状细胞癌上表达(参见例如,kojima等人,j.biol. chem.270:21984-21990(1995)和onda等人,clin.cancer res.12:4225
-ꢀ
4231(2006))。表达间皮素的其他细胞在下文“与间皮素表达相关的疾 病”的定义中提供。间皮素还与ca125/muc16相互作用(参见例如, rump等人,j.biol.chem.279:9190-9198(2004)和ma等人,j.biol. chem.287:33123-33131(2012))。在一个实例中,tfp的抗原结合部 分识别并结合在正常或恶性间皮瘤细胞、卵巢癌细胞、胰腺腺癌细胞 或鳞状细胞癌细胞上表达的间皮素蛋白的细胞外结构域内的表位。
[0099]
如本文所用,术语“抗体”是指衍生自免疫球蛋白分子的蛋白质 或多肽序列,其与抗原特异性结合。抗体可以是多克隆或单克隆来源 的完整免疫球蛋白或其片段,并且可衍生自天然来源或来自重组来源。
[0100]
术语“抗体片段”或“抗体结合结构域”是指抗体或其重组变体的 至少一部分,其含有抗原结合结构域,即完整抗体的抗原决定可变区, 其足以赋予抗体片段对靶标如抗原及其定义的表位的识别和特异性 结合。抗体片段的实例包括但不限于fab、fab
’
、f(ab
’
)2和fv片段、 单链(sc)fv(“scfv”)抗体片段、线性抗体、单结构域抗体(缩写为
ꢀ“
sdab”(v
l
或v
h
)、骆驼科动物v
hh
结构域和由抗体片段形成的多 特异性抗体。
[0101]
术语“scfv”是指这样的融合蛋白,其包含至少一个包含轻链可 变区的抗体片段和至少一个包含重链可变区的抗体片段,其中该轻链 和重链可变区经由短的柔性多肽连接体连续连接,并且能够表达为单 一多肽链,并且其中scfv保留其衍生自的完整抗体的特异性。
[0102]
关于抗体的“重链可变区”或“v
h”“(或在单结构域抗体例如纳米 抗体的情况下为”v
hh“)是指含有插入到被称为构架区的侧翼序列段 之间的三个cdr的重链片段,这些构架区通常比cdr更高度保守并 形成支架以支撑cdr。
[0103]
除非另有说明,否则如本文所用,scfv可以以任一顺序具有v
l
和v
h
可变区,例如,相对于多肽的n末端和c末端,scfv可包含 v
l-连接体-v
h
或可包含v
h-连接体-v
l
。
[0104]
包含抗体或其抗体片段的本发明tfp组合物的部分可以以其中 抗原结合结构域表达为连续多肽链的部分的多种形式存在,包括例如 单结构域抗体片段(sdab)或重链抗
体hcab,衍生自鼠、人源化或 人抗体的单链抗体(scfv)(harlow等人,1999,in:using antibodies: a laboratory manual,cold spring harbor laboratory press,n.y.;harlow 等人,1989,in:antibodies:a laboratory manual,cold spring harbor, n.y.;houston等人,1988,proc.natl.acad.sci.usa 85:5879-5883; bird等人,1988,science 242:423-426)。在一个方面,本发明tfp组 合物的抗原结合结构域包含抗体片段。在另一个方面,tfp包含包括 scfv或sdab在内的抗体片段。
[0105]
术语“抗体重链”是指处于天然构象的抗体分子中存在的两种类 型的多肽链中较大的那种,并且通常决定抗体所属的类别。
[0106]
术语“抗体轻链”是指处于天然构象的抗体分子中存在的两种类 型的多肽链中较小的那种。kappa(“κ”)和lambda(“λ”)轻链是指两 种主要的抗体轻链同种型。
[0107]
术语“重组抗体”是指使用重组dna技术产生的抗体,例如由噬 菌体或酵母表达系统表达的抗体。该术语还应解释为意指通过合成编 码抗体的dna分子和表达抗体蛋白质的dna分子或具体说明抗体 的氨基酸序列所产生的抗体,其中该dna或氨基酸序列已使用本领 域可获得且公知的重组dna或氨基酸序列技术获得。
[0108]
术语“抗原”或“ag”是指能够被抗体特异性结合或以其他方式引 发免疫应答的分子。该免疫应答可涉及抗体产生或特异性免疫感受态 细胞的活化或两者。
[0109]
本领域技术人员将会理解,包括几乎所有蛋白质或肽在内的任 何大分子都可用作抗原。此外,抗原可衍生自重组或基因组dna。本 领域技术人员将会理解,包含编码引发免疫应答的蛋白质的核苷酸序 列或部分核苷酸序列的任何dna因此编码“抗原”(在本文使用该术 语时)。此外,本领域技术人员将会理解,抗原不必仅由基因的全长 核苷酸序列编码。显而易见的是,本发明包括但不限于使用多于一种 基因的部分核苷酸序列,并且这些核苷酸序列以各种组合排列以编码 引发所需免疫应答的多肽。此外,本领域技术人员将会理解,抗原根 本不必由“基因”编码。显而易见的是,抗原可通过合成产生,或可衍 生自生物样品,或者可以是除多肽之外的大分子。这样的生物样品可 包括但不限于组织样品、肿瘤样品、细胞或具有其他生物组分的流体。
[0110]
术语“抗肿瘤作用”是指可通过各种手段显现的生物效果,包括 但不限于例如肿瘤体积减小、肿瘤细胞数目减少、转移的数目减少、 预期寿命延长、肿瘤细胞增殖减少、肿瘤细胞存活率降低或与癌性病 况相关的各种生理症状改善。“抗肿瘤作用”还可以首先通过本发明的 肽、多核苷酸、细胞和抗体预防肿瘤发生的能力来证明。
[0111]
术语“自体”是指来源于与其随后将重新引入的个体相同的个体 的任何物质。
[0112]
术语“同种异体”是指来源于与物质将引入的个体为相同物种的 不同动物或者不同患者的任何物质。当一个或多个基因座处的基因不 相同时,两个或更多个个体被称为彼此同种异体。在一些方面,来自 相同物种的个体的同种异体材料可能在遗传学上充分不同,从而发生 抗原性相互作用。
[0113]
术语“异种”是指来源于不同物种的动物的移植物。
[0114]
术语“癌症”是指特征在于异常细胞的快速且不受控制的生长的 疾病。癌细胞可局部扩散或通过血流和淋巴系统扩散到身体的其他部 位。本文描述了各种癌症的实例,包括但不限于乳腺癌、前列腺癌、 卵巢癌、宫颈癌、皮肤癌、胰腺癌、结直肠癌、肾癌、肝癌、脑癌、 肺癌等。
[0115]
短语“与间皮素表达相关的疾病”包括但不限于与间皮素表达相 关的疾病或与表达间皮素的细胞相关的病况,包括例如增生性疾病, 如癌症或恶性肿瘤或癌前病况。在一个方面,该癌症是间皮瘤。在一 个方面,该癌症是胰腺癌。在一个方面,该癌症是卵巢癌。在一个方 面,该癌症是胃癌。在一个方面,该癌症是肺癌。在一个方面,该癌 症是子宫内膜癌。与间皮素表达相关的非癌症相关适应症包括但不限 于例如自身免疫病(例如,狼疮、类风湿性关节炎、结肠炎)、炎性 病症(变态反应和哮喘)和移植。
[0116]
术语“保守序列修饰”是指不显著影响或改变含有氨基酸序列的 抗体或抗体片段的结合特性的氨基酸修饰。此类保守修饰包括氨基酸 置换、添加和缺失。可通过本领域已知的标准技术如定点诱变和pcr 介导的诱变将修饰引入本发明的抗体或抗体片段中。保守氨基酸置换 是其中氨基酸残基被具有相似侧链的氨基酸残基取代的氨基酸置换。 本领域已经定义了具有相似侧链的氨基酸残基家族。这些家族包括具 有以下侧链的氨基酸:碱性侧链(例如,赖氨酸、精氨酸、组氨酸)、 酸性侧链(例如,天冬氨酸、谷氨酸)、不带电荷的极性侧链(例如, 甘氨酸、天冬酰胺、谷氨酰胺、丝氨酸、苏氨酸、酪氨酸、半胱氨酸、 色氨酸)、非极性侧链(例如,丙氨酸、缬氨酸、亮氨酸、异亮氨酸、 脯氨酸、苯丙氨酸、甲硫氨酸)、β-支链侧链(例如,苏氨酸、缬氨 酸、异亮氨酸)和芳香族侧链(例如,酪氨酸、苯丙氨酸、色氨酸、 组氨酸)。因此,本发明的tfp内的一个或多个氨基酸残基可用来自 相同侧链家族的其他氨基酸残基取代,并且可使用本文所述的功能测 定来测试改变的tfp。
[0117]
术语“刺激”是指通过刺激结构域或刺激分子(例如,tcr/cd3 复合物)与其同源配体结合而诱导的初级应答,从而介导信号转导事 件,诸如但不限于,经由tcr/cd3复合物的信号转导。刺激可介导 某些分子的表达改变和/或细胞骨架结构的重组等。
[0118]
术语“刺激分子”或“刺激结构域”是指由t细胞表达的分子或其 部分,其提供以针对t细胞信号传导途径的至少一些方面的刺激方式 调节tcr复合物的初级活化的初级细胞质信号传导序列。在一个方 面,初级信号通过例如tcr/cd3复合物与装载有肽的mhc分子的 结合而起始,并且其导致包括但不限于增殖、活化、分化等t细胞应 答的介导。以刺激方式起作用的初级细胞质信号传导序列(也称为“初 级信号传导结构域”)可含有信号传导基序,其被称为基于免疫受体酪 氨酸的活化基序或“itam”。在本发明中特别有用的含有itam的初 级细胞质信号传导序列的实例包括但不限于衍生自tcrζ、fcrγ、fcrβ、 cd3γ、cd3δ、cd3ε、cd5、cd22、cd79a、cd79b、cd278(也称 为“icos”)和cd66d的那些序列。
[0119]
术语“抗原呈递细胞”或“apc”是指在其表面上展示与主要组织 相容性复合物(mhc)复合的外来抗原的免疫系统细胞,诸如佐细胞 (例如,b细胞、树突细胞等)。t细胞可使用它们的t细胞受体 (tcr)识别这些复合物。apc加工抗原并将它们呈递给t细胞。
[0120]
如本文所用,术语“细胞内信号传导结构域”是指分子的细胞内 部分。细胞内信号传导结构域产生促进含tfp细胞例如表达tfp的 t细胞的免疫效应物功能的信号。例如在表达tfp的t细胞中的免疫 效应物功能的实例包括细胞溶解活性和t辅助细胞活性,包括细胞因 子的分泌。在一个实施方案中,细胞内信号传导结构域可包含初级细 胞内信号传导结构域。示例性的初级细胞内信号传导结构域包括衍生 自负责初级刺激或抗原依赖性刺激的分子的细胞内信号传导结构域。 在一个实施方案中,细胞内信号传导结构域可包含共刺激细胞内结构 域。示例性的共刺激细胞内信号传导结构域包括衍生自负责共刺激信 号或抗原非依赖性刺激的分子的细胞内信号传导结构域。
[0121]
初级细胞内信号传导结构域可包含itam(“基于免疫受体酪氨 酸的活化基序”)。含有itam的初级细胞质信号传导序列的实例包 括但不限于衍生自cd3ζ、fcrγ、fcrβ、cd3γ、cd3δ、cd3ε、cd5、 cd22、cd79a、cd79b和cd66d dap10和dap12的那些序列。
[0122]
术语“共刺激分子”是指t细胞上的同源结合配偶体,其与共刺 激配体特异性结合,从而介导t细胞的共刺激反应,诸如但不限于增 殖。共刺激分子是有效免疫应答所需的抗原受体或其配体之外的细胞 表面分子。共刺激分子包括但不限于mhc 1类分子、btla和toll 配体受体,以及dap10、dap12、cd30、light、ox40、cd2、cd27、 cd28、cds、icam-1、lfa-1(cd11a/cd18)和4-1bb(cd137)。 共刺激细胞内信号传导结构域可以是共刺激分子的细胞内部分。共刺 激分子可在以下蛋白质家族中表示:tnf受体蛋白质、免疫球蛋白样 蛋白质、细胞因子受体、整联蛋白、信号传导性淋巴细胞活化分子 (slam蛋白)和激活性nk细胞受体。此类分子的实例包括cd27、 cd28、4-1bb(cd137)、ox40、gitr、cd30、cd40、icos、baffr、 hvem、淋巴细胞功能相关抗原1(lfa-1)、cd2、cd7、light、 nkg2c、slamf7、nkp80、cd160、b7-h3以及与cd83特异性结 合的配体等。细胞内信号传导结构域可包含其衍生自的分子的整个细 胞内部分或整个天然细胞内信号传导结构域,或其功能片段。术语“4
-ꢀ
1bb”是指tnfr超家族的成员,其具有以genbank登录号 aaa62478.2提供的氨基酸序列,或来自非人物种如小鼠、啮齿动物、 猴、猿等的等同残基;“4-1bb共刺激结构域”被定义为genbank登录 号aaa62478.2的氨基酸残基214-255,或来自非人物种如小鼠、啮 齿动物、猴、猿等的等同残基。
[0123]
术语“编码”是指多核苷酸如基因、cdna或mrna中的特定核 苷酸序列用作在生物过程中合成其他聚合物和大分子的模板的固有 性质,该聚合物和大分子具有确定的核苷酸序列(例如,rrna、trna 和mrna)或确定的氨基酸序列以及由此产生的生物学性质。因此, 如果对应于基因的mrna的转录和翻译在细胞或其他生物系统中产 生蛋白质,则该基因、cdna或rna编码蛋白质。编码链及其核苷酸 序列均与mrna序列相同并且通常在序列表中提供,而用作基因或 cdna转录的模板的非编码链可以称为编码蛋白质或者该基因或 cdna的其他产物。
[0124]
除非另有说明,否则“编码氨基酸序列的核苷酸序列”包括彼此 为简并形式且编码相同氨基酸序列的所有核苷酸序列。短语编码蛋白 质或rna的“核苷酸序列”也可包括内含子,其程度为编码蛋白质的 核苷酸序列在一些形式中可含有一个或多个内含子。
[0125]
术语“有效量”或“治疗有效量”在本文中可互换使用,并且是指 有效实现特定生物学或治疗结果的如本文所述的化合物、制剂、物质 或组合物的量。
[0126]
术语“内源”是指来自生物体、细胞、组织或系统内或者在生物 体、细胞、组织或系统内产生的任何物质。
[0127]
术语“外源”是指从生物体、细胞、组织或系统外引入或者在生 物体、细胞、组织或系统外产生的任何物质。
[0128]
术语“表达”是指由启动子驱动的特定核苷酸序列的转录和/或 翻译。
[0129]
术语“转移载体”是指包含分离的核酸并且可用于将所分离的核 酸递送至细胞内部的物质的组合物。许多载体是本领域已知的,包括 但不限于线性多核苷酸、与离子或两亲化合物缔合的多核苷酸、质粒 和病毒。因此,术语“转移载体”包括自主复制的质粒或病毒。该术语 还应解释为进一步包含促进将核酸转移到细胞中的非质粒和非病毒 化合物,
例如聚赖氨酸化合物、脂质体等。病毒转移载体的实例包括 但不限于腺病毒载体、腺伴随病毒载体、逆转录病毒载体、慢病毒载 体等。
[0130]
术语“表达载体”是指包含重组多核苷酸的载体,该重组多核苷 酸包含与待表达的核苷酸序列可操作地连接的表达控制序列。表达载 体包含足以用于表达的顺式作用元件;用于表达的其他元件可由宿主 细胞或在体外表达系统中提供。表达载体包括本领域已知的所有表达 载体,包括粘粒、质粒(例如,裸露的或包含在脂质体中)和掺有重 组多核苷酸的病毒(例如,慢病毒、逆转录病毒、腺病毒和腺伴随病 毒)。
[0131]
术语“慢病毒”是指逆转录病毒科的一个属。慢病毒在逆转录病 毒中在能够感染非分裂细胞方面是独特的;它们可以将大量的遗传信 息递送到宿主细胞的dna中,因此它们是基因递送载体的最有效的 方法之一。hiv、siv和fiv均为慢病毒的实例。
[0132]
术语“慢病毒载体”是指衍生自慢病毒基因组的至少一部分的载 体,特别包括milone等人,mol.ther.17(8):1453-1464(2009)中提供 的自失活慢病毒载体。可以在临床中使用的慢病毒载体的其他实例包 括但不限于例如来自oxford biomedica的lentivector
tm
基因递 送技术、来自lentigen的lentimax
tm
载体系统等。非临床类型的 慢病毒载体也是可获得的并且是本领域技术人员已知的。
[0133]
术语“同源”或“同一性”是指两个聚合物分子之间,例如,两个 核酸分子如两个dna分子或两个rna分子之间,或两个多肽分子之 间的亚单位序列同一性。当两个分子中的亚单位位置被相同的单体亚 单位占据时;例如,如果两个dna分子中的每一个的位置被腺嘌呤 占据,则它们在该位置是同源的或相同的。两个序列之间的同源性是 匹配或同源位置的数目的直接函数;例如,如果两个序列中的一半位 置(例如,长度为10个亚单位的聚合物中的5个位置)是同源的, 则这两个序列是50%同源的;如果90%的位置(例如,10个中的9 个)是匹配的或同源的,则这两个序列是90%同源的。
[0134]“人源化”形式的非人(例如,鼠)抗体是含有衍生自非人免疫 球蛋白的最小序列的嵌合免疫球蛋白、免疫球蛋白链或其片段(诸如 fv、fab、fab
’
、f(ab
’
)2或抗体的其他抗原结合子序列)。在大多数情 况下,人源化抗体及其抗体片段是人免疫球蛋白(接受者抗体或抗体 片段),其中来自接受者的互补决定区(cdr)的残基被来自非人物 种如小鼠、大鼠或兔的具有所需特异性、亲和力和容量的cdr(供体 抗体)的残基取代。在一些情况下,人免疫球蛋白的fv构架区(fr) 残基被相应的非人残基所取代。此外,人源化抗体/抗体片段可包含既 不存在于接受者抗体中也不存在于输入的cdr或构架序列中的残基。 这些修饰可以进一步改进和优化抗体或抗体片段性能。通常,人源化 抗体或其抗体片段将会包含基本上全部或至少一个,通常为两个可变 域中,其中全部或基本上全部cdr区对应于非人免疫球蛋白的cdr 区,并且全部或大部分fr区是人免疫球蛋白序列的fr区。人源化 抗体或抗体片段还可包含免疫球蛋白恒定区(fc)的至少一部分,通 常是人免疫球蛋白的fc的至少一部分。进一步的细节参见jones等 人,nature,321:522-525,1986;reichman等人,nature,332:323-329, 1988;presta,curr.op.struct.biol.,2:593-596,1992。
[0135]“人”或“完全人”是指这样的免疫球蛋白,如抗体或抗体片段, 其中整个分子是人源的或由与人形式的抗体或免疫球蛋白相同的氨 基酸序列组成。
[0136]
术语“分离的”意指从天然状态改变或移出的。例如,天然存在 于活动物中的核酸或肽不是“分离的”,但与其天然状态下共同存在的 物质部分或完全分离的相同核酸或肽
是“分离的”。分离的核酸或蛋白 质可以以基本上纯化的形式存在,或者可存在于非天然环境如宿主细 胞中。
[0137]
在本发明的上下文中,对常见的核酸碱基使用以下缩写。“a”是 指腺苷,“c”是指胞嘧啶,“g”是指鸟苷,“t”是指胸苷,“u”是指尿苷。
[0138]
术语“可操作地连接”或“转录控制”是指调节序列与异源核酸序 列之间的功能性连接,其导致后者的表达。例如,当第一核酸序列与 第二核酸序列以功能性关系布置时,第一核酸序列与第二核酸序列可 操作地连接。例如,如果启动子影响编码序列的转录或表达,则该启 动子与该编码序列可操作地连接。可操作地连接的dna序列可彼此 邻接,并且例如,在需要连接两个蛋白质编码区的情况下,该dna序 列在相同的阅读框中。
[0139]
术语“肠胃外”施用免疫原性组合物包括例如皮下(s.c.)、静脉 内(i.v.)、肌肉内(i.m.)或胸骨内注射,肿瘤内或输注技术。
[0140]
术语“核酸”或“多核苷酸”是指单链或双链形式的脱氧核糖核酸 (dna)或核糖核酸(rna)及其聚合物。除非明确限定,否则该术 语包括含有天然核苷酸的已知类似物的核酸,其具有与参考核酸相似 的结合特性,并以与天然存在的核苷酸相似的方式代谢。除非另有说 明,否则特定核酸序列还隐含地包括其保守修饰的变体(例如,简并 密码子置换)、等位基因、直向同源物、snp和互补序列以及明确指 出的序列。具体地,简并密码子置换可通过产生其中一个或多个选定 的(或所有)密码子的第三位置被混合碱基和/或脱氧肌苷残基置换的 序列来实现(batzer等人,nucleic acid res.19:5081(1991);ohtsuka 等人,j.biol.chem.260:2605-2608(1985);和rossolini等人,mol.cell. probes 8:91-98(1994))。
[0141]
术语“肽”、“多肽”和“蛋白质”可互换使用,并且是指由通过肽键 共价连接的氨基酸残基组成的化合物。蛋白质或肽必须含有至少两个 氨基酸,并且对可构成蛋白质或肽序列的氨基酸的最大数目没有限制。 多肽包括包含通过肽键彼此连接的两个或更多个氨基酸的任何肽或 蛋白质。如本文所用,该术语是指短链和长链两者,短链在本领域中 通常也称为例如肽、寡肽和寡聚体,长链在本领域中通常称为蛋白质, 其存在很多类型。“多肽”包括例如生物活性片段、基本上同源的多肽、 寡肽、同源二聚体、异源二聚体、多肽的变体、修饰的多肽、衍生物、 类似物、融合蛋白等。多肽包括天然肽、重组肽或其组合。
[0142]
术语“启动子”是指被细胞转录机制或引入的合成机制识别的、 启动多核苷酸序列的特定转录所需的dna序列。
[0143]
术语“启动子/调节序列”是指表达与启动子/调节序列可操作地 连接的基因产物所需的核酸序列。在一些情况下,该序列可以是核心 启动子序列,而在其他情况下,该序列还可包含增强子序列和表达基 因产物所需的其他调节元件。启动子/调节序列可以是例如以组织特异 性方式表达基因产物的启动子/调节序列。
[0144]
术语“组成型”启动子是指当与编码或指定基因产物的多核苷酸 可操作地连接时,在细胞的大多数或所有生理条件下导致在细胞中产 生基因产物的核苷酸序列。
[0145]
术语“诱导型”启动子是指当与编码或指定基因产物的多核苷酸 可操作地连接时,基本上仅当对应于该启动子的诱导物存在于细胞中 时才导致在细胞中产生基因产物的核苷酸序列。。
[0146]
术语“组织特异性”启动子是指当与基因编码或指定的多核苷酸 可操作地连接
时,基本上仅在细胞是对应于该启动子的组织类型的细 胞的情况下才导致在细胞中产生基因产物的核苷酸序列。
[0147]
在scfv的上下文中使用的术语“连接体”和“柔性多肽连接体”是 指由单独或组合使用的氨基酸如甘氨酸和/或丝氨酸残基组成的肽连 接体,以将可变重链区和可变轻链区连接在一起。在一个实施方案中, 柔性多肽连接体是gly/ser连接体,并且包含氨基酸序列(gly-gly-gly
-ꢀ
ser)
n
,其中n是等于或大于1的正整数。例如,n=1、n=2、n=3、 n=4、n=5、n=6、n=7、n=8、n=9、n=10。在一个实施方案中, 柔性多肽连接体包括但不限于(gly4ser)4或(gly4ser)3。在另一个实施 方案中,连接体包括(gly2ser)、(glyser)或(gly3ser)的多个重复。本发 明的范围内还包括描述于wo2012/138475(通过引用并入本文)中的 连接体。在一些情况下,连接体序列包括长连接体(ll)序列。在一些 情况下,长连接体序列包括(g4s)
n
,其中n=2至4。在一些情况下,连接 体序列包括短连接体(sl)序列。在一些情况下,短连接体序列包括(g4s)
n
, 其中n=1至3。
[0148]
如本文所用,5
’
帽(也称为rna帽、rna 7-甲基鸟苷帽或rnam7g帽)是在转录开始后不久已添加至真核信使rna的“前”端或5
’ꢀ
端的经修饰的鸟嘌呤核苷酸。5
’
帽由与第一转录核苷酸连接的末端基 团组成。其存在对于核糖体的识别和保护免受rna酶作用至关重要。 帽的添加与转录偶合,并且共同转录地发生,使得每一个都影响另一 个。在转录开始后不久,合成的mrna的5
’
端被与rna聚合酶缔合 的帽合成复合物结合。该酶促复合物催化mrna加帽所需的化学反 应。合成作为多步生化反应进行。可修饰加帽部分以调节mrna的功 能,诸如其稳定性或翻译效率。
[0149]
如本文所用,“体外转录的rna”是指已在体外合成的rna,优 选mrna。通常,体外转录的rna由体外转录载体产生。体外转录 载体包含用于产生体外转录的rna的模板。
[0150]
如本文所用,“聚(a)”是通过对mrna的聚腺苷酸化而附接的 一系列腺苷。在用于瞬时表达的构建体的优选实施方案中,聚a为50 至5000个,优选多于64个,更优选多于100个,最优选多于300或 400个。聚(a)序列可被化学或酶促修饰以调节mrna功能,诸如定 位、稳定性或翻译效率。
[0151]
如本文所用,“聚腺苷酸化”是指聚腺苷酰基部分或其修饰变体 与信使rna分子的共价连接。在真核生物中,大多数信使rna (mrna)分子在3
’
末端被聚腺苷酸化。3
’
聚(a)尾是通过聚腺苷酸聚 合酶这种酶的作用添加到前mrna的长腺嘌呤核苷酸序列(通常数 百个)。在高等真核生物中,将聚(a)尾添加到含有特定序列聚腺苷酸 化信号的转录物上。聚(a)尾和与其结合的蛋白质有助于保护mrna 不被核酸外切酶降解。聚腺苷酸化对于转录终止、从细胞核输出 mrna和翻译也是重要的。聚腺苷酸化在dna转录为rna后立即 在细胞核中发生,但另外还可稍后在细胞质中发生。在转录终止后, 通过与rna聚合酶缔合的内切核酸酶复合物的作用切割mrna链。 切割位点的特征通常在于切割位点附近存在碱基序列aauaaa。在 mrna被切割后,腺苷残基被添加到切割位点处的游离3
’
端。
[0152]
如本文所用,“瞬时”是指非整合的转基因表达数小时、数天或 数周的时段,其中表达的时间段短于当基因整合到基因组中或包含在 宿主细胞中的稳定质粒复制子内时基因表达的时间段。
[0153]
术语“信号转导途径”是指多个信号转导分子之间的生物化学关 系,它们在信号从细胞的一部分到细胞的另一部分的传递中起作用。 短语“细胞表面受体”包括能够接收
信号并跨细胞膜传送信号的分子 和分子复合物。
[0154]
术语“受试者”旨在包括其中可引发免疫应答的活生物体(例如, 哺乳动物、人)。
[0155]
术语“基本上纯化的”细胞是指基本上不含其他细胞类型的细胞。 基本上纯化的细胞还指已经与其在天然存在状态下通常关联的其他 细胞类型分离的细胞。在一些情况下,基本上纯化的细胞群体是指均 质的细胞群体。在其他情况下,该术语仅指已经与其在天然状态下天 然关联的细胞分离的细胞。在一些方面,细胞在体外培养。在其他方 面,细胞不在体外培养。
[0156]
如本文所用,术语“治疗性”意指治疗。治疗效果通过减少、抑 制、缓解或根除疾病状态而获得。
[0157]
如本文所用,术语“预防”是指对疾病或疾病状态的预防或保护 性处理。
[0158]
在本发明的上下文中,“肿瘤抗原”或“过度增生性病症抗原”或
ꢀ“
与过度增生性病症相关的抗原”是指特定过度增生性病症共有的抗 原。在某些方面,本发明的过度增生性病症抗原来源于癌症,包括但 不限于原发性或转移性黑素瘤、
[0159]
间皮瘤、肾细胞癌、胃癌、乳腺癌、肺癌、卵巢癌、前列腺癌、 结肠癌、宫颈癌、脑癌、肝癌、胰腺癌、肾癌、子宫内膜癌和胃癌。
[0160]
在一些情况下,所述疾病是选自下组的癌症:间皮瘤、乳头状 浆液性卵巢腺癌、透明细胞卵巢癌、混合型米勒氏卵巢癌、子宫内膜 粘液性卵巢癌、恶性胸膜疾病、胰腺腺癌、胰腺导管腺癌、子宫浆液 性癌、肺腺癌、肝外胆管癌、胃腺癌、食管腺癌、结直肠腺癌、乳腺 腺癌、与间皮素表达相关的疾病及其组合、与间皮素表达相关的疾病 及其组合。
[0161]
术语“转染”或“转化”或“转导”是指借以将外源核酸转移或引入 宿主细胞的过程。“转染的”或“转化的”或“转导的”细胞是已经用外源 核酸转染、转化或转导的细胞。该细胞包括原代受试细胞及其后代。
[0162]
术语“特异性结合”是指抗体、抗体片段或特异性配体,其识别 并结合样品中存在的同源结合配偶体(例如,间皮素),但不一定且 基本上不识别或结合样品中的其他分子。
[0163]
范围:在整个本公开内容中,本发明的各个方面可以以范围形 式呈现。应当理解,范围形式的描述仅为了方便和简洁起见,而不应 解释为对本发明范围的僵化限制。因此,应当认为范围的描述具体公 开了所有可能的子范围以及该范围内的单个数值。例如,应当认为范 围的描述如1至6具有具体公开的子范围,如1至3、1至4、1至5、2至4、2至6、3至6等,以及该范围内的单个数字,例如1、2、2.7、 3、4、5、5.3和6。作为另一实例,诸如95-99%同一性的范围包括具 有95%、96%、97%、98%或99%同一性的某物,并且包括诸如96
-ꢀ
99%、96-98%、96-97%、97-99%、97-98%和98-99%同一性等子范围。 无论范围的广度如何,这都适用。描述
[0164]
本文提供了使用t细胞受体(tcr)融合蛋白治疗疾病如癌症 的物质组合物和使用方法。如本文所用,“t细胞受体(tcr)融合蛋 白”或“tfp”包括衍生自包含tcr的各种多肽的重组多肽,其通常能 够i)与靶细胞上的表面抗原结合,并且ii)与完整tcr复合物的其 他多肽组分相互作用,通常在共同位于t细胞表面之中或之上时。如 本文提供的,与嵌合抗原受体相比,tfp提供了实质性益处。术语“嵌 合抗原受体”或替代地“car”是指包含scfv形式的细胞外抗原结合结 构域、跨膜结构域和细胞质信号传导结构域(本文也称为“细胞内信
号 传导结构域”)的重组多肽,该细胞质信号传导结构域包含衍生自如下 定义的刺激分子的功能性信号传导结构域。通常,car的中心细胞内 信号传导结构域衍生自通常发现与tcr复合物缔合的cd3ζ链。 cd3ζa信号传导结构域可与衍生自至少一种共刺激分子如4-1bb(即 cd137)、cd27和/或cd28的一个或多个功能性信号传导结构域融 合。t细胞受体(tcr)融合蛋白(tfp)
[0165]
本发明包括编码tfp的重组dna构建体,其中该tfp包含与 间皮素例如人间皮素特异性结合的抗体片段,其中该抗体片段的序列 与编码tcr亚单位或其部分的核酸序列邻接并在同一阅读框中。本 文提供的tfp能够与一个或多个内源(或替代地,一个或多个外源, 或者内源与外源的组合)tcr亚单位缔合,以便形成功能性tcr复 合物。
[0166]
在一个方面,本发明的tfp包含靶标特异性结合元件,其另外 被称为抗原结合结构域。部分的选择取决于限定靶细胞表面的靶抗原 的类型和数目。例如,可选择抗原结合结构域以识别作为靶细胞上与 特定疾病状态相关的细胞表面标志物的靶抗原。因此,可作为本发明 tfp中抗原结合结构域的靶抗原的细胞表面标志物的实例包括与病 毒、细菌和寄生物感染、自身免疫病和癌性疾病(例如,恶性疾病) 相关的标志物。
[0167]
在一个方面,通过将抗原结合结构域工程化到特异性结合所需 抗原的tfp中,可使tfp介导的t细胞应答针对感兴趣的抗原。
[0168]
在一个方面,包含抗原结合结构域的tfp部分包含靶向间皮素 的抗原结合结构域。在一个方面,抗原结合结构域靶向人间皮素。
[0169]
抗原结合结构域可以是与抗原结合的任何结构域,包括但不限 于单克隆抗体、多克隆抗体、重组抗体、人抗体、人源化抗体及其功 能片段,包括但不限于单结构域抗体,如重链可变域(v
h
)、轻链可 变域(v
l
)和骆驼科动物来源的纳米抗体的可变域(v
hh
),以及本 领域已知作为抗原结合结构域的替代性支架,诸如重组纤连蛋白结构 域、anticalin、darpin等。同样,特异性识别并结合靶抗原的天然或 合成配体可用作tfp的抗原结合结构域。在一些情况下,抗原结合结 构域来源于tfp将会最终在其中使用的相同的物种是有益的。例如, 对于在人类中使用,tfp的抗原结合结构域包含抗体或抗体片段的抗 原结合结构域的人或人源化残基可能是有益的。
[0170]
因此,在一个方面,抗原结合结构域包含人源化或人抗体或抗 体片段,或者鼠抗体或抗体片段。在一个实施方案中,人源化或人抗 间皮素结合结构域包含本文所述的人源化或人抗间皮素结合结构域 的轻链互补决定区1(lc cdr1)、轻链互补决定区2(lc cdr2) 和轻链互补决定区3(lc cdr3)中的一个或多个(例如,全部三个), 和/或本文所述的人源化或人抗间皮素结合结构域的重链互补决定区 1(hc cdr1)、重链互补决定区2(hc cdr2)和重链互补决定区3 (hc cdr3)中的一个或多个(例如,全部三个),例如,包含一个 或多个,例如全部三个lc cdr以及一个或多个,例如全部三个hccdr的人源化或人抗间皮素结合结构域。在一个实施方案中,人源化 或人抗间皮素结合结构域包含本文所述的人源化或人抗间皮素结合 结构域的重链互补决定区1(hc cdr1)、重链互补决定区2(hccdr2)和重链互补决定区3(hc cdr3)中的一个或多个(例如,全 部三个),例如,人源化或人抗间皮素结合结构域具有两个可变重链 区,每个包含本文所述的hc cdr1、hc cdr2和hc cdr3。在一个 实施方案中,人源化或人抗间皮素结合结构域包含本文所述的人源化 或人轻链可变区和/或本文所述的人源化或人重链可变区。在一个实施 方案中,人源化或人抗间皮素结合
结构域包含本文所述的人源化重链 可变区,例如本文所述的人源化或人重链可变区中的至少两个。在一 个实施方案中,抗间皮素结合结构域是scfv,其包含本文提供的氨基 酸序列的轻链和重链。在一个实施方案中,抗间皮素结合结构域(例 如,scfv)包含:轻链可变区,其包含相对于本文提供的轻链可变区 的氨基酸序列具有至少一个、两个或三个修饰(例如,置换)但不多 于30、20或10个修饰(例如,置换)的氨基酸序列,或与本文提供 的氨基酸序列具有95-99%同一性的序列;和/或重链可变区,其包含 相对于本文提供的重链可变区的氨基酸序列具有至少一个、两个或三 个修饰(例如,置换)但不多于30、20或10个修饰(例如,置换) 的氨基酸序列,或与本文提供的氨基酸序列具有95-99%同一性的序 列。在一个实施方案中,人源化或人抗间皮素结合结构域是scfv,并 且包含本文所述的氨基酸序列的轻链可变区经由连接体例如本文所 述的连接体与包含本文所述的氨基酸序列的重链可变区连接。在一个 实施方案中,人源化抗间皮素结合结构域包括(gly
4-ser)
n
连接体,其 中n为1、2、3、4、5或6,优选3或4。scfv的轻链可变区和重链 可变区可以例如处于任何以下方向:轻链可变区-连接体-重链可变区 或重链可变区-连接体-轻链可变区。在一些情况下,连接体序列包括长 连接体(ll)序列。在一些情况下,长连接体序列包括(g4s)
n
,其中n=2 至4。在一些情况下,连接体序列包括短连接体(sl)序列。在一些情 况下,短连接体序列包括(g4s)
n
,其中n=1至3。
[0171]
在一些方面,对非人抗体进行人源化,其中该抗体的特定序列 或区域被修饰以增加与人类中天然产生的抗体或其片段的相似性。在 一个方面,对抗原结合结构域进行人源化。
[0172]
可使用本领域已知的多种技术产生人源化抗体,包括但不限于 cdr移植(参见例如,欧洲专利号ep 239,400;国际公开号wo 91/09967;以及美国专利号5,225,539、5,530,101和5,585,089,其各 自通过引用整体并入本文)、镶饰(veneering)或表面再塑(resurfacing) (参见例如,欧洲专利号ep 592,106和ep 519,596;padlan,1991, molecular immunology,28(4/5):489-498;studnicka等人,1994,proteinengineering,7(6):805-814;以及roguska等人,1994,pnas,91:969
-ꢀ
973,其各自通过引用整体并入本文)、链改组(参见例如,美国专利 号5,565,332,其通过引用整体并入本文)以及公开于以下文献的技术: 例如,美国专利申请公开号us2005/0042664、美国专利申请公开号 us2005/0048617、美国专利号6,407,213、美国专利号5,766,886、国 际公开号wo 9317105、tan等人,j.immunol.,169:1119-25(2002)、 caldas等人,protein eng.,13(5):353-60(2000)、morea等人,methods, 20(3):267-79(2000)、baca等人,j.biol.chem.,272(16):10678-84(1997)、 roguska等人,protein eng.,9(10):895-904(1996)、couto等人,cancerres.,55(23增刊):5973s-5977s(1995)、couto等人,cancer res., 55(8):1717-22(1995),sandhu j s,gene,150(2):409-10(1994)和 pedersen等人,j.mol.biol.,235(3):959-73(1994),其各自通过引用整 体并入本文。通常,构架区中的构架残基将被来自cdr供体抗体的 相应残基置换,以改变(例如改善)抗原结合。这些构架置换通过本 领域公知的方法鉴定,例如,通过模拟cdr与构架残基的相互作用 以鉴定对抗原结合至关重要的构架残基,以及序列比较以鉴定特定位 置处的异常构架残基(参见例如,queen等人,美国专利号5,585,089; 和riechmann等人,1988,nature,332:323,其通过引用整体并入本文)。
[0173]
人源化抗体或抗体片段具有保持其所来自的非人来源的一个或 多个氨基酸残
基。这些非人氨基酸残基通常被称为“输入”残基,其通 常取自“输入”可变域。如本文提供的,人源化抗体或抗体片段包含来 自非人免疫球蛋白分子和构架区的一个或多个cdr,其中构成构架的 氨基酸残基完全或主要来源于人种系。用于抗体或抗体片段的人源化 的多种技术是本领域公知的,并且基本上可按照winter及其同事的 方法(jones等人,nature,321:522-525(1986);riechmann等人,nature, 332:323-327(1988);verhoeyen等人,science,239:1534-1536(1988)) 进行,其中用啮齿动物的cdr或cdr序列置换人抗体的相应序列, 即cdr移植(ep 239,400;pct公开号wo 91/09967;以及美国专利 号4,816,567;6,331,415;5,225,539;5,530,101;5,585,089;6,548,640, 其内容通过引用整体并入本文)。在这样的人源化抗体和抗体片段中, 基本上小于完整的人可变域被来自非人物种的相应序列置换。人源化 抗体通常是这样的人抗体,其中一些cdr残基和可能的一些构架(fr) 残基被来自啮齿动物抗体中类似位点的残基置换。抗体和抗体片段的 人源化还可通过镶饰或表面再塑(ep 592,106;ep 519,596;padlan, 1991,molecular immunology,28(4/5):489-498;studnicka等人,proteinengineering,7(6):805-814(1994);和roguska等人,pnas,91:969-973 (1994))或者链改组(美国专利号5,565,332)来实现,其内容通过引 用整体并入本文。
[0174]
用于制备人源化抗体的人可变域(轻链和重链)的选择是为了 降低抗原性。根据所谓的“最佳适配”方法,在已知人可变域序列的整 个文库中筛选啮齿动物抗体的可变域的序列。然后接受最接近啮齿动 物序列的人序列作为人源化抗体的人构架(fr)(sims等人,j. immunol.,151:2296(1993);chothia等人,j.mol.biol.,196:901(1987), 其内容通过引用整体并入本文。另一种方法使用衍生自所有人抗体的 特定轻链或重链亚组的共有序列的特定构架。相同的构架可用于几种 不同的人源化抗体(参见例如,nicholson等人mol.immun.34(16
-ꢀ
17):1157-1165(1997);carter等人,proc.natl.acad.sci.usa,89:4285 (1992);presta等人,j.immunol.,151:2623(1993),其内容通过引用整 体并入本文)。在一些实施方案中,重链可变区的构架区,例如所有 四个构架区均衍生自v
h
4-4-59种系序列。在一个实施方案中,构架区 可包含一个、两个、三个、四个或五个修饰,例如置换,例如来自相 应鼠序列的氨基酸的修饰。在一个实施方案中,轻链可变区的构架区, 例如所有四个构架区均衍生自vk3-2.25种系序列。在一个实施方案 中,构架区可包含一个、两个、三个、四个或五个修饰,例如置换, 例如来自相应鼠序列的氨基酸的修饰。
[0175]
在一些方面,包含抗体片段的本发明tfp组合物的部分被人源 化,保留对靶抗原的高亲和力和其他有利的生物学性质。根据本发明 的一个方面,通过使用亲本和人源化序列的三维模型分析亲本序列和 各种概念性人源化产物的过程制备人源化抗体和抗体片段。三维免疫 球蛋白模型通常是可获得的并且是本领域技术人员熟知的。可获得说 明并展示所选择的候选免疫球蛋白序列的可能的三维构象结构的计 算机程序。检查这些展示允许分析残基在候选免疫球蛋白序列的功能 中的可能作用,例如分析影响候选免疫球蛋白与靶抗原结合的能力的 残基。以这种方式,可从接受者和输入序列中选择并组合fr残基, 从而实现所需的抗体或抗体片段特性,诸如对靶抗原增加的亲和力。 通常,cdr残基直接且最大程度地参与影响抗原结合。
[0176]
人源化抗体或抗体片段可保留与原始抗体相似的抗原特异性, 例如,在本发明中为与人间皮素结合的能力。在一些实施方案中,人 源化抗体或抗体片段可具有改善的与人
间皮素结合的亲和力和/或特 异性。
[0177]
在一个方面,抗间皮素结合结构域的特征在于抗体或抗体片段 的特定功能特征或性质。例如,在一个方面,包含抗原结合结构域的 本发明tfp组合物的部分与人间皮素特异性结合。在一个方面,抗原 结合结构域与nicholson等人mol.immun.34(16-17):1157-1165 (1997)描述的fmc63 scfv具有相同或相似的对人间皮素的结合特异 性。在一个方面,本发明涉及包含抗体或抗体片段的抗原结合结构域, 其中该抗体结合结构域与间皮素蛋白质或其片段特异性结合,其中该 抗体或抗体片段包括包含本文提供的氨基酸序列的可变轻链和/或可 变重链。在某些方面,scfv与前导序列邻接并在同一阅读框中。
[0178]
在一个方面,抗间皮素结合结构域是片段,例如单链可变片段 (scfv)。在一个方面,抗间皮素结合结构域是fv、fab、(fab
’
)2或 双功能(例如双特异性)杂合抗体(例如,lanzavecchia等人,eur.j. immunol.17,105(1987))。在一个方面,本文公开的抗体及其片段以 野生型或增强的亲和力与间皮素蛋白质结合。
[0179]
本文还提供了用于获得对靶抗原(例如,间皮素或本文其他地 方针对融合部分结合结构域的靶标所描述的任何靶抗原)具有特异性 的抗体抗原结合结构域的方法,该方法包括通过在本文所示v
h
结构 域的氨基酸序列中添加、缺失、置换或插入一个或多个氨基酸的方式 提供作为该v
h
结构域的氨基酸序列变体的v
h
结构域,任选地将由此 提供的v
h
结构域与一个或多个v
l
结构域组合,并测试v
h
结构域或 一个或多个v
h
/v
l
组合,以鉴定对感兴趣的靶抗原(例如,间皮素) 具有特异性并任选地具有一种或多种所需性质的特异性结合成员或 抗体抗原结合结构域。
[0180]
在一些情况下,v
h
结构域和scfv可根据本领域已知的方法制 备(参见例如,bird等人,(1988)science 242:423-426和huston等人, (1988)proc.natl.acad.sci.usa 85:5879-5883)。scfv分子可通过使 用柔性多肽连接体将v
h
和v
l
区连接在一起来产生。scfv分子包含 具有优化的长度和/或氨基酸组成的连接体(例如,ser-gly连接体)。 连接体长度可极大地影响scfv的可变区的折叠和相互作用的方式。 事实上,如果采用短多肽连接体(例如,5-10个氨基酸),则防止了 链内折叠。还需要链间折叠以将两个可变区组合在一起以形成功能性 表位结合位点。在一些情况下,连接体序列包括长连接体(ll)序列。 在一些情况下,长连接体序列包括(g4s)
n
,其中n=2至4。在一些情况下, 连接体序列包括短连接体(sl)序列。在一些情况下,短连接体序列包 括(g4s)
n
,其中n=1至3。连接体取向和大小的实例参见例如hollinger 等人1993proc natl acad.sci.u.s.a.90:6444-6448、美国专利号 7,695,936、美国专利申请公开号20050100543和20050175606以及 pct公开号wo2006/020258和wo2007/024715,其全部通过引用并 入本文。
[0181]
scfv在其v
h
和v
l
区之间可包含约10、11、12、13、14、15个 或多于15个残基的连接体。连接体序列可包含任何天然存在的氨基 酸。在一些实施方案中,连接体序列包含氨基酸甘氨酸和丝氨酸。在 另一个实施方案中,连接体序列包含成组的甘氨酸和丝氨酸重复,诸 如(gly4ser)
n
,其中n是等于或大于1的正整数。在一个实施方案中, 连接体可以是(gly4ser)4或(gly4ser)3。连接体长度的变化可保留或增 强活性,从而在活性研究中产生优异的功效。在一些情况下,连接体 序列包括长连接体(ll)序列。在一些情况下,长连接体序列包括(g4s)
n
, 其中n=2至4。在一些情况下,连接体序列包括短连接体(sl)序列。 在一些情况下,短连接体序列包括(g4s)
n
,其中n=1至3。
稳定性和突变
[0182]
可参考常规对照scfv分子或全长抗体的生物物理性质(例如, 热稳定性)来评价抗间皮素结合结构域例如scfv分子(例如,可溶性 scfv)的稳定性。在一个实施方案中,人源化或人scfv在所述测定中 具有比亲本scfv高约0.1、约0.25、约0.5、约0.75、约1、约1.25、 约1.5、约1.75、约2、约2.5、约3、约3.5、约4、约4.5、约5、约 5.5、约6、约6.5、约7、约7.5、约8、约8.5、约9、约9.5、约10 摄氏度、约11摄氏度、约12摄氏度、约13摄氏度、约14摄氏度或 约15摄氏度的热稳定性。
[0183]
随后将抗间皮素结合结构域例如scfv的提高的热稳定性赋予 整个间皮素-tfp构建体,从而改善抗间皮素tfp构建体的治疗性质。 与常规抗体相比,抗间皮素结合结构域例如scfv的热稳定性可提高 至少约2℃或3℃。在一个实施方案中,与常规抗体相比,抗间皮素 结合结构域例如scfv具有1℃的提高的热稳定性。在另一个实施方 案中,与常规抗体相比,抗间皮素结合结构域例如scfv具有2℃的 提高的热稳定性。在另一个实施方案中,与常规抗体相比,scfv具有 4℃、5℃、6℃、7℃、8℃、9℃、10℃、11℃、12℃、13℃、14℃或 15℃的提高的热稳定性。例如,可在本文公开的scfv分子与scfv v
h
和v
l
所衍生自的抗体的scfv分子或fab片段之间进行比较。可使用 本领域已知的方法测量热稳定性。例如,在一个实施方案中,可测量 t
m
。用于测量t
m
的方法和确定蛋白质稳定性的其他方法在下文描述。
[0184]
scfv中的突变(通过可溶性scfv的人源化或诱变产生)改变 scfv的稳定性并提高scfv和抗间皮素tfp构建体的总体稳定性。使 用诸如t
m
、温度变性和温度聚集等测量将人源化scfv的稳定性与鼠 scfv进行比较。在一个实施方案中,抗间皮素结合结构域例如scfv包 含由人源化过程产生的至少一个突变,使得突变的scfv赋予抗间皮 素tfp构建体提高的稳定性。在另一个实施方案中,抗间皮素结合结 构域例如scfv包含由人源化过程产生的至少1、2、3、4、5、6、7、 8、9、10个突变,使得突变的scfv赋予间皮素-tfp构建体提高的稳 定性。
[0185]
在一个方面,tfp的抗原结合结构域包含与本文所述的抗原结 合结构域氨基酸序列同源的氨基酸序列,并且该抗原结合结构域保留 本文所述的抗间皮素抗体片段所需的功能性质。在一个具体方面,本 发明的tfp组合物包含抗体片段。在另一个方面,该抗体片段包含 scfv。
[0186]
在各个方面,通过修饰一个或两个可变区(例如,v
h
和/或v
l
) 内,例如一个或多个cdr区内和/或一个或多个构架区内的一个或多 个氨基酸来工程化tfp的抗原结合结构域。在一个具体方面,本发明 的tfp组合物包含抗体片段。在另一个方面,该抗体片段包含scfv。
[0187]
本领域普通技术人员将会理解,可进一步修饰本发明的抗体或 抗体片段,使得它们在氨基酸序列上(例如,相对于野生型)有所变 化,但在所需活性上没有变化。例如,可对蛋白质进行另外的核苷酸 置换,导致“非必需”氨基酸残基处的氨基酸置换。例如,分子中的非 必需氨基酸残基可被来自相同侧链家族的另一个氨基酸残基取代。在 另一个实施方案中,氨基酸串可被结构相似但在顺序和/或组成上与侧 链家族成员不同的串取代,例如,可进行保守置换,其中氨基酸残基 被具有相似侧链的氨基酸残基所取代。
[0188]
本领域已经定义了具有相似侧链的氨基酸残基家族,包括碱性 侧链(例如,赖氨酸、精氨酸、组氨酸)、酸性侧链(例如,天冬氨 酸、谷氨酸)、不带电荷的极性侧链(例如,甘
氨酸、天冬酰胺、谷 氨酰胺、丝氨酸、苏氨酸、酪氨酸、半胱氨酸)、非极性侧链(例如, 丙氨酸、缬氨酸、亮氨酸、异亮氨酸、脯氨酸、苯丙氨酸、甲硫氨酸、 色氨酸)、β-支链侧链(例如,苏氨酸、缬氨酸、异亮氨酸)和芳香 族侧链(例如,酪氨酸、苯丙氨酸、色氨酸、组氨酸)。
[0189]
在两个或更多个核酸或多肽序列的上下文中,同一性百分比是 指相同的两个或更多个序列。当在比较窗口或指定区域上针对最大对 应性进行比较和比对时,如使用以下序列比较算法中的一种或者通过 手动比对和目视检查所测量的,如果两个序列具有指定百分比的相同 的氨基酸残基或核苷酸(例如,在指定区域上,或者在未指定的情况 下在整个序列上60%的同一性,任选地70%、71%、72%、73%、74%、 75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、 86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、 97%、98%或99%的同一性),则这两个序列“基本上相同”。任选地, 同一性存在于长度为至少约50个核苷酸(或10个氨基酸)的区域上, 或更优选地存在于长度为100至500个或1000个或更多个核苷酸(或 者20、50、200个或更多个氨基酸)的区域上。
[0190]
对于序列比较,通常一个序列充当参考序列,测试序列与之进 行比较。当使用序列比较算法时,将测试序列和参考序列输入计算机, 在需要的情况下指定子序列坐标,并指定序列算法程序参数。可使用 默认程序参数,或得可指定替代参数。随后,序列比较算法基于程序 参数计算测试序列相对于参考序列的序列同一性百分比。用于比较的 序列比对方法是本领域公知的。可通过以下方法进行用于比较的最佳 序列比对:例如smith和waterman,(1970)adv.appl.math.2:482c的 局部同源性算法;needleman和wunsch,(1970)j.mol.biol.48:443的 同源性比对算法;pearson和lipman,(1988)proc.nat
’
l.acad.sci.usa 85:2444的相似性检索法;这些算法(wisconsin genetics software package,genetics computer group,575science dr.,madison,wis.中的 gap、bestfit、fasta和tfasta)的计算机化实现;或手动比对 和目视检查(参见例如,brent等人,(2003)current protocols inmolecular biology)。适用于确定序列同一性百分比和序列相似性的 算法的两个实例是blast和blast 2.0算法,其分别描述于altschul 等人(1977)nuc.acids res.25:3389-3402;和altschul等人(1990)j. mol.biol.215:403-410中。用于进行blast分析的软件可通过国家 生物技术信息中心公开获得。
[0191]
在一个方面,本发明考虑到产生功能上等同的分子的起始抗体 或片段(例如,scfv)氨基酸序列的修饰。例如,可修饰tfp中包含 的抗间皮素结合结构域例如scfv的v
h
或v
l
,以保留抗间皮素结合 结构域例如scfv的起始v
h
或v
l
构架区的至少约70%、71%、72%、 73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、 84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、 95%、96%、97%、98%、99%的同一性。本发明考虑到整个tfp构建 体的修饰,例如,tfp构建体的各个结构域的一个或多个氨基酸序列 的修饰,以便产生功能上等同的分子。可修饰tfp构建体以保留起始 tfp构建体的至少约70%、71%、72%、73%、74%、75%、76%、77%、 78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、 89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99% 的同一性。细胞外结构域
[0192]
细胞外结构域可衍生自天然来源或来自重组来源。在天然来源 的情况下,该结构域可衍生自任何蛋白质,但特别是膜结合蛋白质或 跨膜蛋白质。在一个方面,细胞外结构
域能够与跨膜结构域缔合。在 本发明中特别有用的细胞外结构域可包括至少以下细胞外区域:例如 t细胞受体的α、β或ζ链,或者cd3ε、cd3γ或cd3δ,或者在替代 实施方案中,包括cd28、cd45、cd4、cd5、cd8、cd9、cd16、 cd22、cd33、cd37、cd64、cd80、cd86、cd134、cd137、cd154。跨膜结构域
[0193]
通常,tfp序列含有由单个基因组序列编码的细胞外结构域和 跨膜结构域。在替代实施方案中,tfp可被设计成包含与tfp的细胞 外结构域异源的跨膜结构域。跨膜结构域可包含与跨膜区域相邻的一 个或多个另外的氨基酸,例如,与跨膜结构域所衍生自的蛋白质的细 胞外区域缔合的一个或多个氨基酸(例如,细胞外区域的1、2、3、 4、5、6、7、8、9、10个或至多15个氨基酸),和/或与跨膜蛋白质 所衍生自的蛋白质的细胞内区域缔合的一个或多个另外的氨基酸(例 如,细胞内区域的1、2、3、4、5、6、7、8、9、10个或至多15个氨 基酸)。在一个方面,跨膜结构域是与所用tfp的其他结构域之一缔 合的跨膜结构域。在一些情况下,可通过氨基酸置换来选择或修饰跨 膜结构域,以避免这样的结构域与相同或不同表面膜蛋白的跨膜结构 域结合,例如,以使与受体复合物的其他成员的相互作用最小化。在 一个方面,跨膜结构域能够与tfp-t细胞表面上的另一个tfp同源 二聚化。在不同方面,可修饰或置换跨膜结构域的氨基酸序列,以便 使与相同tfp中存在的天然结合配偶体的结合结构域的相互作用最 小化。
[0194]
跨膜结构域可衍生自天然来源或来自重组来源。在天然来源的 情况下,该结构域可衍生自任何膜结合蛋白质或跨膜蛋白质。在一个 方面,每当tfp与靶标结合时,跨膜结构域能够向细胞内结构域发信 号。在本发明中特别有用的跨膜结构域可包括至少以下跨膜区域:例 如t细胞受体的α、β或ζ链,或者cd28、cd3ε、cd45、cd4、cd5、 cd8、cd9、cd16、cd22、cd33、cd37、cd64、cd80、cd86、cd134、 cd137、cd154。
[0195]
在一些情况下,跨膜结构域可经由铰链(例如来自人类蛋白质 的铰链)与tfp的细胞外区域(例如tfp的抗原结合结构域)连接。 例如,在一个实施方案中,该铰链可以是人免疫球蛋白(ig)铰链, 例如igg4铰链或cd8a铰链。连接体
[0196]
任选地,长度为2至10个氨基酸的短寡肽或多肽连接体可形成 tfp的跨膜结构域与细胞质区域之间的连接。甘氨酸-丝氨酸双联体 提供了特别合适的连接体。例如,在一个方面,该连接体包含 ggggsggggs的氨基酸序列。在一些实施方案中,该连接体由 ggtggcggaggttctggaggtggaggttcc的核苷酸序列编码。细胞质结构域
[0197]
如果tfp含有cd3γ、δ或ε多肽,则tfp的细胞质结构域可 包括细胞内信号传导结构域;tcrα和tcrβ亚单位通常缺乏信号传 导结构域。细胞内信号传导结构域通常负责激活其中已引入tfp的免 疫细胞的至少一种正常效应物功能。术语“效应物功能”是指细胞的特 化功能。例如,t细胞的效应物功能可以是细胞溶解活性或辅助活性, 包括细胞因子的分泌。因此,术语“细胞内信号传导结构域”是指蛋白 质的部分,其转导效应物功能信号并引导细胞执行特化功能。虽然通 常可采用整个细胞内信号传导结构域,但在许多情况下不必使用整条 链。就使用细胞内信号传导结构域的截短部分来说,可使用这样的截 短部分代替完整链,只要其转导效应物功能信号即可。因此,术语“细 胞内信号传导结构域”意在包
括足以转导效应物功能信号的细胞内信 号传导结构域的任何截短部分。
[0198]
用于本发明tfp的细胞内信号传导结构域的实例包括t细胞受 体(tcr)的细胞质序列和协同作用以在抗原受体接合后启动信号转 导的共同受体,以及任何这些序列的衍生物或变体以及具有相同功能 能力的任何重组序列。
[0199]
已知仅通过tcr产生的信号不足以完全激活幼稚t细胞,并 且需要次级和/或共刺激信号。因此,幼稚t细胞活化可以说是由两 种不同类别的细胞质信号传导序列介导的:通过tcr启动抗原依赖 性初级激活的信号传导序列(初级细胞内信号传导结构域)和以非抗 原依赖性方式起作用以提供次级或共刺激信号的信号传导序列(次级 细胞质结构域,例如共刺激结构域)。
[0200]
初级信号传导结构域以刺激方式或以抑制方式调节tcr复合 物的初级激活。以刺激方式起作用的初级细胞内信号传导结构域可含 有信号传导基序,其被称为基于免疫受体酪氨酸的活化基序(itam)。
[0201]
在本发明中特别有用的含有itam的初级细胞内信号传导结构 域的实例包括cd3ζ、fcrγ、fcrβ、cd3γ、cd3δ、cd3ε、cd5、cd22、 cd79a、cd79b和cd66d的细胞内信号传导结构域。在一个实施方案 中,本发明的tfp包含细胞内信号传导结构域,例如cd3ε的初级信 号传导结构域。在一个实施方案中,初级信号传导结构域包含修饰的 itam结构域,例如与天然itam结构域相比具有改变的(例如,升 高或降低的)活性的突变itam结构域。在一个实施方案中,初级信 号传导结构域包括含有修饰的itam的初级细胞内信号传导结构域, 例如,含有优化的和/或截短的itam的初级细胞内信号传导结构域。 在一个实施方案中,初级信号传导结构域包含一个、两个、三个、四 个或更多个itam基序。
[0202]
tfp的细胞内信号传导结构域可包含单独的cd3ζ信号传导结 构域,或者其可与在本发明tfp的背景下有用的任何其他所需的细胞 内信号传导结构域组合。例如,tfp的细胞内信号传导结构域可包含 cd3ε链部分和共刺激信号传导结构域。共刺激信号传导结构域是指 包含共刺激分子的细胞内结构域的tfp的一部分。共刺激分子是除了 抗原受体或其配体之外的细胞表面分子,其为淋巴细胞对抗原的有效 应答所需的。此类分子的实例包括cd27、cd28、4-1bb(cd137)、 ox40、dap10、dap12、cd30、cd40、pd1、icos、淋巴细胞功能 相关抗原-1(lfa-1)、cd2、cd7、light、nkg2c、b7-h3以及与 cd83特异性结合的配体等。例如,已证明cd27共刺激增强人tfp
-ꢀ
t细胞在体外的扩充、效应物功能和存活,并增强人t细胞在体内的 持久性和抗肿瘤活性(song等人blood.2012;119(3):696-706)。
[0203]
本发明tfp的细胞质部分内的细胞内信号传导序列可以以随机 或指定的顺序彼此连接。任选地,短的寡肽或多肽连接体,例如,长 度为2至10个氨基酸(例如,2、3、4、5、6、7、8、9或10个氨基 酸),可在细胞内信号传导序列之间形成连接。
[0204]
在一个实施方案中,甘氨酸-丝氨酸双联体可用作合适的连接体。 在一个实施方案中,单个氨基酸,例如丙氨酸、甘氨酸,可用作合适 的连接体。
[0205]
在一个方面,本文所述的表达tfp的细胞可进一步包含第二 tfp,例如,包含不同的抗原结合结构域的第二tfp,该抗原结合结 构域例如针对相同的靶标(间皮素)或不同的靶标(例如,cd123)。 在一个实施方案中,当表达tfp的细胞包含两种或更多种不同的tfp 时,不同tfp的抗原结合结构域可使得抗原结合结构域不彼此相互作 用。例如,表达第一和第二tfp的细胞可具有第一tfp的抗原结合 结构域,例如片段,例如scfv,其不与第二tfp的
抗原结合结构域缔 合,例如,第二tfp的抗原结合结构域是v
hh
。
[0206]
在另一个方面,本文所述的表达tfp的细胞可进一步表达另一 种药剂,例如增强表达tfp的细胞的活性的药剂。例如,在一个实施 方案中,该药剂可以是抑制抑制性分子的药剂。在一些实施方案中, 抑制性分子,例如pd1,可降低表达tfp的细胞产生免疫效应物应答 的能力。抑制性分子的实例包括pd1、pd-l1、ctla4、tim3、lag3、 vista、btla、tigit、lair1、cd160、2b4和tgfrβ。在一个实 施方案中,抑制抑制性分子的药剂包含第一多肽,例如抑制性分子, 其与向细胞提供正信号的第二多肽例如本文所述的细胞内信号传导 结构域缔合。在一个实施方案中,所述药剂包含第一多肽,例如抑制 性分子如pd1、lag3、ctla4、cd160、btla、lair1、tim3、2b4 和tigit的多肽,或其中任一种的片段(例如,其中任一种的细胞外 结构域的至少一部分),以及第二多肽,其为本文所述的细胞内信号 传导结构域(例如,包含共刺激结构域(例如,4-1bb、cd27或cd28, 例如,如本文所述)和/或初级信号传导结构域(例如,本文所述的cd3ζ 信号传导结构域))。在一个实施方案中,所述药剂包含pd1或其片 段(例如,pd1的细胞外结构域的至少一部分)的第一多肽和本文所 述的细胞内信号传导结构域的第二多肽(例如,本文所述的cd28信 号传导结构域和/或本文所述的cd3ζ信号传导结构域)。pd1是cd28 受体家族的抑制性成员,该家族还包括cd28、ctla-4、icos、和 btla。pd-1在活化的b细胞、t细胞和髓样细胞上表达(agata等 人1996int.immunol 8:765-75)。已经显示pd1的两种配体pd-l1和 pd-l2在与pd1结合后下调t细胞活化(freeman等人2000j expmed 192:1027-34;latchman等人2001nat immunol 2:261-8;carter等 人2002eur j immunol 32:634-43)。pd-l1在人类癌症中是丰富的 (dong等人2003j mol med 81:281-7;blank等人2005cancer immunol. immunother 54:307-314;konishi等人2004clin cancer res 10:5094)。 通过抑制pd1与pd-l1的局部相互作用可逆转免疫抑制。
[0207]
在一个实施方案中,所述药剂包含抑制性分子的细胞外结构域 (ecd),例如,程序性死亡1(pd1)可以与跨膜结构域和可选的细 胞内信号传导结构域如41bb和cd3ζ融合(本文中也称pd1 tfp)。 在一个实施方案中,pd1 tfp当与本文所述的抗间皮素tfp组合使用 时改善t细胞的持久性。在一个实施方案中,该tfp是包含pd1的 细胞外结构域的pd1 tfp。或者,提供了含有抗体或抗体片段如scfv 的tfp,其与程序性死亡配体1(pd-l1)或程序性死亡配体2(pd
-ꢀ
l2)特异性结合。
[0208]
在另一个方面,本发明提供了表达tfp的t细胞例如tfp-t细 胞的群体。在一些实施方案中,表达tfp的t细胞群体包含表达不同 tfp的细胞的混合物。例如,在一个实施方案中,tfp-t细胞群体可 包括表达具有本文所述抗间皮素结合结构域的tfp的第一细胞和表 达具有不同抗间皮素结合结构域(例如,不同于第一细胞所表达的 tfp中的抗间皮素结合结构域的本文所述的抗间皮素结合结构域)的 tfp的第二细胞。作为另一个实例,表达tfp的细胞群体可包含表达 包含例如本文所述抗间皮素结合结构域的tfp的第一细胞和表达包 含针对除间皮素外的靶标(例如,另一种肿瘤相关抗原)的抗原结合 结构域的tfp的第二细胞。
[0209]
在另一个方面,本发明提供一种细胞群体,其中该群体中的至 少一种细胞表达具有本文所述的抗间皮素结构域的tfp,并且第二种 细胞表达另一种药剂,例如,增强表达tfp的细胞的活性的药剂。例 如,在一个实施方案中,该药剂可以是抑制抑制性分子的药剂。在一 些实施方案中,抑制性分子例如可降低表达tfp的细胞产生免疫效应 物应答的能
力。抑制性分子的实例包括pd1、pd-l1、pd-l2、ctla4、 tim3、lag3、vista、btla、tigit、lair1、cd160、2b4和tgfrβ。 在一个实施方案中,抑制抑制性分子的药剂包含第一多肽,例如抑制 性分子,其与向细胞提供正信号的第二多肽例如本文所述的细胞内信 号传导结构域缔合。
[0210]
本文公开了用于产生编码tfp的体外转录的rna的方法。本 发明还包括可直接转染到细胞中的编码tfp的rna构建体。产生用 于转染的mrna的方法可涉及用特别设计的引物对模板进行体外转 录(ivt),然后添加聚a,以产生含有3
’
和5
’
非翻译序列(“utr”)、 5
’
帽和/或内部核糖体进入位点(ires)、待表达的核酸和聚a尾的 构建体,通常长度为50-2000个碱基。如此产生的rna可有效地转 染不同种类的细胞。在一个方面,模板包含tfp的序列。
[0211]
在一个方面,抗间皮素tfp由信使rna(mrna)编码。在一 个方面,将编码抗间皮素tfp的mrna引入t细胞中以产生tfp-t 细胞。在一个实施方案中,体外转录的rna tfp可作为瞬时转染的 形式引入细胞中。使用聚合酶链反应(pcr)产生的模板通过体外转 录产生rna。可使用适当的引物和rna聚合酶将来自任何来源的感 兴趣的dna通过pcr直接转化为用于体外mrna合成的模板。dna 的来源可以是例如基因组dna、质粒dna、噬菌体dna、cdna、 合成dna序列或任何其他适当的dna来源。用于体外转录的所需 模板是本发明的tfp。在一个实施方案中,待用于pcr的dna含有 开放阅读框。该dna可来自生物基因组的天然存在的dna序列。在 一个实施方案中,该核酸可包含5
’
和/或3
’
非翻译区(utr)中的一些 或全部。该核酸可包含外显子和内含子。在一个实施方案中,待用于 pcr的dna是人核酸序列。在另一个实施方案中,待用于pcr的dna是包含5
’
和3
’
utr的人核酸序列。或者,该dna可以是通常 不在天然存在的生物体中表达的人工dna序列。示例性人工dna序 列是包含连接在一起以形成编码融合蛋白的开放阅读框的基因部分 的序列。连接在一起的dna部分可来自单个生物体或来自多于一个 生物体。
[0212]
使用pcr来产生用于体外转录mrna的模板,该mrna用于 转染。进行pcr的方法是本领域公知的。用于pcr的引物被设计成 具有与待用作pcr模板的dna区域基本上互补的区域。如本文所 用,“基本上互补”是指这样的核苷酸序列,其中引物序列中的大部分 或全部碱基是互补的,或者一个或多个碱基是非互补的或错配的。基 本上互补的序列能够在用于pcr的退火条件下与预期的dna靶标退 火或杂交。引物可设计成与dna模板的任何部分基本上互补。例如, 引物可设计用于扩增通常在细胞中转录的核酸部分(开放阅读框), 包括5
’
和3
’
utr。引物还可设计用于扩增编码感兴趣的特定结构域 的核酸部分。在一个实施方案中,引物被设计用于扩增人cdna的编 码区,包括5
’
和3
’
utr的全部或部分。可用于pcr的引物可通过本 领域公知的合成方法产生。“正向引物”是含有与待扩增的dna序列 上游的dna模板上的核苷酸基本上互补的核苷酸区域的引物。“上游
”ꢀ
在本文中用于指相对于编码链,处于待扩增的dna序列的5
’
侧的位 置。“反向引物”是含有与待扩增的dna序列下游的双链dna模板基 本上互补的核苷酸区域的引物。“下游”在本文用于指相对于编码链, 处于待扩增的dna序列的3
’
侧的位置。
[0213]
对于pcr有用的任何dna聚合酶均可在本文公开的方法中使 用。试剂和聚合酶可从许多来源商购。
[0214]
还可使用具有促进稳定性和/或翻译效率的能力的化学结构。 rna优选具有5
’
和
3
’
utr。在一个实施方案中,5
’
utr的长度为1 至3,000个核苷酸。待添加至编码区的5
’
和3
’
utr序列的长度可通 过不同的方法改变,包括但不限于设计用于pcr的、与utr的不同 区域退火的引物。使用该方法,本领域普通技术人员可改变在转染所 转录的rna后实现最佳翻译效率所需的5
’
和3
’
utr长度。
[0215]5’
和3
’
utr可以是感兴趣的核酸的天然存在的内源5
’
和3
’ꢀ
utr。或者,可通过将utr序列掺入正向和反向引物中或通过模板 的任何其他修饰来添加对感兴趣的核酸而言不是内源的utr序列。 使用对感兴趣的核酸而言不是内源的utr序列可用于改变rna的稳 定性和/或翻译效率。例如,已知3
’
utr序列中富含au的元件可降 低mrna的稳定性。因此,可基于本领域公知的utr的性质来选择 或设计3
’
utr,以提高转录的rna的稳定性。
[0216]
在一个实施方案中,5
’
utr可含有内源核酸的kozak序列。或 者,当如上所述通过pcr添加对感兴趣的核酸而言不是内源的5
’
utr 时,可通过添加5
’
utr序列重新设计共有kozak序列。kozak序列 可提高一些rna转录物的翻译效率,但似乎不是所有rna实现有效 翻译所需的。许多mrna对kozak序列的需要是本领域已知的。在 其他实施方案中,5
’
utr可以是rna基因组在细胞中稳定的rna 病毒的5
’
utr。在其他实施方案中,3
’
或5
’
utr中可使用各种核苷 酸类似物以妨碍mrna的外切核酸酶降解。
[0217]
为了实现从dna模板合成rna而无需基因克隆,转录启动子 应连接到待转录的序列上游的dna模板上。当作为rna聚合酶启动 子的序列添加到正向引物的5
’
端时,该rna聚合酶启动子被掺入待 转录的开放阅读框上游的pcr产物中。在一个优选的实施方案中,该 启动子是t7聚合酶启动子,如本文其他地方所述。其他有用的启动 子包括但不限于t3和sp6 rna聚合酶启动子。t7、t3和sp6启动 子的共有核苷酸序列是本领域已知的。
[0218]
在一个优选的实施方案中,mrna具有5
’
端的帽和3
’
聚(a)尾, 其决定了mrna在细胞中的核糖体结合、翻译起始和稳定性。在环状 dna模板例如质粒dna上,rna聚合酶产生长的多联(concatameric) 产物,其不适于在真核细胞中表达。在3
’
utr的末端线性化的质粒 dna的转录产生正常大小的mrna,其在真核转染中无效,即使其 在转录后发生聚腺苷酸化。
[0219]
在线性dna模板上,噬菌体t7 rna聚合酶可使转录物的3
’ꢀ
端延伸超出模板的最后碱基(schenborn和mierendorf,nuc acids res., 13:6223-36(1985);nacheva和berzal-herranz,eur.j.biochem., 270:1485-65(2003))。
[0220]
将聚a/t段整合到dna模板中的常规方法是分子克隆。然而, 整合到质粒dna中的聚a/t序列可导致质粒不稳定,这就是为什么 从细菌细胞获得的质粒dna模板经常高度具有缺失和其他畸变的原 因。这使得克隆程序不仅费力和耗时,而且通常不可靠。这就是为什 么非常需要允许构建具有聚a/t 3
’
段的dna模板而无需克隆的方法 的原因。
[0221]
转录dna模板的聚a/t区段可在pcr期间通过使用含有聚t 尾如100t尾(大小可以是50-5000个t)的反向引物产生,或者在 pcr之后通过任何其他方法产生,该方法包括但不限于dna连接或 体外重组。聚(a)尾也为rna提供稳定性并减少其降解。通常,聚(a) 尾的长度与所转录的rna的稳定性正相关。在一个实施方案中,聚 (a)尾为100至5000个腺苷。
[0222]
在使用聚(a)聚合酶如大肠杆菌聚a聚合酶(e-pap)进行体外 转录后,rna的聚(a)尾可进一步延伸。在一个实施方案中,将聚(a) 尾的长度从100个核苷酸增加到300至400个核苷酸导致rna的翻 译效率增加到约2倍。另外,不同化学基团与3
’
端的连接可增加mrna 稳定性。这样的连接可含有修饰的/人工核苷酸、适体和其他化合物。 例如,可使用聚(a)聚
合酶将atp类似物掺入聚(a)尾中。atp类似物 可进一步提高rna的稳定性。
[0223]5’
帽也为rna分子提供稳定性。在优选的实施方案中,通过本 文公开的方法产生的rna包含5
’
帽。使用本领域已知和本文描述的 技术提供5
’
帽(cougot等人,trends in biochem.sci.,29:436-444(2001); stepinski等人,rna,7:1468-95(2001);elango等人,biochim.biophys. res.commun.,330:958-966(2005))。
[0224]
通过本文公开的方法产生的rna还可含有内部核糖体进入位 点(ires)序列。ires序列可以是任何病毒的、染色体的或人工设计 的序列,其起始不依赖于帽的核糖体与mrna的结合并促进翻译的 起始。可包括适用于细胞电穿孔的任何溶质,其可包含促进细胞渗透 性和活力的因素,诸如糖、肽、脂质、蛋白质、抗氧化剂和表面活性 剂。
[0225]
可使用许多不同方法中的任一种将rna引入靶细胞中,例如 可商购的方法,包括但不限于电穿孔(amaxa nucleofector-ii(amaxabiosystems,cologne,germany))、ecm 830(btx)(harvard instruments, boston,mass.)或gene pulser ii(biorad,denver,colo.)、multiporator (eppendort,hamburg germany)、使用脂质转染的阳离子脂质体介 导的转染、聚合物包封、肽介导的转染或者生物射弹(biolistic)颗粒 递送系统如“基因枪”(参见例如,nishikawa等人hum gene ther., 12(8):861-70(2001))。编码tfp的核酸构建体
[0226]
本发明还提供了编码本文所述的一种或多种tfp构建体的核酸 分子。在一个方面,该核酸分子作为信使rna转录物提供。在一个 方面,该核酸分子作为dna构建体提供。
[0227]
编码所需分子的核酸序列可使用本领域已知的重组方法获得, 例如通过筛查来自表达该基因的细胞的文库,通过从已知包含该基因 的载体衍生该基因,或使用标准技术直接从含有该基因的细胞和组织 中分离。或者,可通过合成方式产生感兴趣的基因,而不是克隆。
[0228]
本发明还提供了本发明dna所插入的载体。衍生自逆转录病 毒如慢病毒的载体是实现长期基因转移的合适工具,因为它们允许转 基因的长期、稳定的整合及其在子细胞中的繁殖。相对于衍生自肿瘤 逆转录病毒如鼠白血病病毒的载体,慢病毒载体具有额外的优势,因 为它们可转导非增殖细胞,如肝细胞。它们还具有低免疫原性的额外 优势。
[0229]
在另一个实施方案中,包含编码本发明所需tfp的核酸的载体 是腺病毒载体(a5/35)。在另一个实施方案中,编码tfp的核酸的 表达可使用诸如sleeping beauty、crisper、cas9和锌指核酸酶等转座 子完成(参见june等人2009nature reviews immunol.9.10:704-716, 通过引用并入本文)。
[0230]
使用标准基因递送方案,本发明的表达构建体也可用于核酸免 疫和基因疗法。用于基因递送的方法是本领域已知的(参见例如,美 国专利号5,399,346、5,580,859、5,589,466,通过引用整体并入本文)。 在另一个实施方案中,本发明提供了基因疗法载体。
[0231]
核酸可克隆到许多类型的载体中。例如,核酸可克隆到包括但 不限于质粒、噬菌粒、噬菌体衍生物、动物病毒和粘粒等载体中。特 别感兴趣的载体包括表达载体、复制载体、探针产生载体和测序载体。
[0232]
此外,表达载体可以以病毒载体的形式提供给细胞。病毒载体 技术是本领域中公知的,并且描述于例如sambrook等人,2012, molecular cloning:a laboratory manual,第1-4卷,cold spring harborpress,ny,以及其他病毒学和分子生物学手册中。可用作载
体的病毒 包括但不限于逆转录病毒、腺病毒、腺伴随病毒、疱疹病毒和慢病毒。 通常,合适的载体含有在至少一种生物体中起作用的复制起点、启动 子序列、方便的限制性内切核酸酶位点和一种或多种选择性标记(例 如,wo 01/96584;wo 01/29058;和美国专利号6,326,193)。
[0233]
已经开发了许多基于病毒的系统用于将基因转移到哺乳动物细 胞中。例如,逆转录病毒为基因递送系统提供了方便的平台。可使用 本领域已知的技术将选定的基因插入载体中并包装在逆转录病毒颗 粒中。然后可分离重组病毒并将其递送至受试者的体内或离体细胞。 许多逆转录病毒系统是本领域已知的。在一些实施方案中,使用腺病 毒载体。许多腺病毒载体是本领域已知的。在一个实施方案中,使用 慢病毒载体。
[0234]
另外的启动子元件,例如增强子,调节转录起始的频率。通常, 这些元件位于起始位点上游30-110bp的区域中,但已经显示许多启 动子也包含起始位点下游的功能元件。启动子元件之间的间隔经常是 灵活的,以便当元件相对于彼此倒置或移动时保持启动子功能。在胸 苷激酶(tk)启动子中,启动子元件之间的间隔可在活性开始下降之 前增加至相距50bp。取决于启动子,看来单个元件可协同或独立地 起作用以激活转录。
[0235]
能够在哺乳动物t细胞中表达tfp转基因的启动子的实例是 ef1a启动子。天然ef1a启动子驱动延伸因子-1复合物的α亚单位的 表达,后者负责将氨酰基trna酶促递送至核糖体。ef1a启动子已广 泛用于哺乳动物表达质粒中,并已显示有效驱动克隆到慢病毒载体中 的转基因的tfp表达(参见例如,milon等人,mol.ther.17(8):1453
-ꢀ
1464(2009))。启动子的另一个实例是即时早期巨细胞病毒(cmv) 启动子序列。该启动子序列是强组成型启动子序列,其能够驱动与其 可操作地连接的任何多核苷酸序列的高水平表达。然而,还可使用其 他组成型启动子序列,包括但不限于猿猴病毒40(sv40)早期启动 子、小鼠乳腺肿瘤病毒(mmtv)、人免疫缺陷病毒(hiv)长末端 重复序列(ltr)启动子、momulv启动子、禽白血病病毒启动子、 eb病毒即时早期启动子、劳斯肉瘤病毒启动子以及人基因启动子, 诸如但不限于肌动蛋白启动子、肌球蛋白启动子、延伸因子-1a启动 子、血红蛋白启动子和肌酸激酶启动子。此外,本发明不应限于使用 组成型启动子。也考虑诱导型启动子作为本发明的一部分。诱导型启 动子的使用提供了分子开关,其能够在需要表达时启动其可操作地连 接的多核苷酸序列的表达,或者当不需要表达时关闭该表达。诱导型 启动子的实例包括但不限于金属硫蛋白启动子、糖皮质激素启动子、 孕酮启动子和四环素调节的启动子。
[0236]
为了评估tfp多肽或其部分的表达,待引入细胞的表达载体还 可含有选择性标记基因或报告基因或两者,以便于从试图通过病毒载 体转染或感染的细胞群体中鉴定和选择表达细胞。在其他方面,选择 性标记可在单独的dna片段上携带并在共转染程序中使用。选择性 标记和报告基因都可侧接适当的调节序列以实现在宿主细胞中的表 达。有用的选择性标记包括例如抗生素抗性基因,诸如neo等。
[0237]
报告基因用于鉴定可能转染的细胞以及用于评价调节序列的功 能。通常,报告基因是不存在于接受者生物体或组织中或不由其表达 的基因,并且编码其表达通过一些易于检测的性质例如酶活性来表现 的多肽。在将dna引入接受者细胞后,在适当的时间测定报告基因 的表达。合适的报告基因可包括编码萤光素酶、β-半乳糖苷酶、氯霉 素乙酰转移酶、分泌的碱性磷酸酶的基因或者绿色荧光蛋白基因(例 如,ui-tei等人,2000febs letters 479:79-82)。合适的表达系统是 公知的,并且可使用已知技术制备或商业获得。
通常,具有显示报告 基因的最高表达水平的最小5
’
侧翼区的构建体被鉴定为启动子。此类 启动子区域可与报告基因连接,并用来评价药剂调节启动子驱动的转 录的能力。
[0238]
将基因引入细胞中并将其表达到细胞中的方法是本领域已知的。 在表达载体的背景下,可通过本领域的任何方法将载体容易地引入宿 主细胞,例如哺乳动物、细菌、酵母或昆虫细胞。例如,可通过物理、 化学或生物学手段将表达载体转移到宿主细胞中。
[0239]
用于将多核苷酸引入宿主细胞中的物理方法包括磷酸钙沉淀、 脂质转染、粒子轰击、显微注射、电穿孔等。用于产生包含载体和/或 外源核酸的细胞的方法是本领域公知的(参见例如,sambrook等人, 2012,molecular cloning:a laboratory manual,第1-4卷,cold spring harbor press,ny)。用于将多核苷酸引入宿主细胞中的一种方法是磷 酸钙转染
[0240]
用于将感兴趣的多核苷酸引入宿主细胞中的生物学方法包括使 用dna和rna载体。病毒载体,尤其是逆转录病毒载体,已成为用 于将基因插入哺乳动物例如人细胞中的最广泛使用的方法。其他病毒 载体可来源于慢病毒、痘病毒、单纯疱疹病毒i、腺病毒和腺伴随病 毒等(参见例如,美国专利号5,350,674和5,585,362)。
[0241]
用于将多核苷酸引入宿主细胞中的化学方法包括胶体分散系统, 诸如大分子复合物、纳米胶囊、微球、珠子和基于脂质的系统,包括 水包油乳液、胶束、混合胶束和脂质体。用作体外和体内递送载体的 示例性胶体系统是脂质体(例如,人造膜囊泡)。可使用最先进的靶 向递送核酸的其他方法,诸如用靶向纳米颗粒或其他合适的亚微米大 小的递送系统递送多核苷酸。
[0242]
在利用非病毒递送系统的情况下,示例性递送载体是脂质体。 考虑使用脂质制剂将核酸引入宿主细胞(体外、离体或体内)。在另 一个方面,核酸可与脂质缔合。与脂质缔合的核酸可包封在脂质体的 水性内部、散布在脂质体的脂双层内、经由与脂质体和寡核苷酸同时 缔合的连接分子附接于脂质体、包埋在脂质体中、与脂质体复合、分 散在含有脂质的溶液中、与脂质混合、与脂质组合、作为悬浮液包含 在脂质中、含有胶束或与胶束复合,或以其他方式与脂质缔合。脂质、 脂质/dna或脂质/表达载体相关的组合物不限于溶液中的任何特定 结构。例如,它们可以以双层结构、胶束或“坍缩”结构存在。它们也 可简单地散布在溶液中,可能形成大小或形状不均匀的聚集体。脂质 是脂肪物质,其可以是天然存在的或合成的脂质。例如,脂质包括天 然存在于细胞质中的脂肪小滴以及含有长链脂肪族烃及其衍生物的 化合物类别,诸如脂肪酸、醇、胺、氨基醇和醛。
[0243]
适合使用的脂质可从商业来源获得。例如,二肉豆蔻基磷脂酰 胆碱(“dmpc”)可从sigma,st.louis,mo获得;双十六烷基磷酸 酯(“dcp”)可从k&k laboratories(plainview,n.y.)获得;胆固 醇(“choi”)可从calbiochem-behring获得;二肉豆蔻基磷脂酰甘油 (“dmpg”)和其他脂质可从avanti polar lipids,inc.(birmingham, ala.)获得。脂质在氯仿或氯仿/甲醇中的原液可以在约-20℃下储存。 氯仿用作唯一的溶剂,因为它比甲醇更容易蒸发。“脂质体”是通用术 语,包括通过产生封闭的脂双层或聚集体而形成的多种单层和多层脂 质媒介物。脂质体的特征可在于具有囊泡结构,该结构具有磷脂双层 膜和内部水性介质。多层脂质体具有被水性介质分隔开的多个脂质层。 它们在磷脂悬浮于过量的水溶液中时自发形成。脂质组分在形成封闭 结构之前经历自重排,并在脂双层之间捕获水和溶解的溶质(ghosh 等人,1991glycobiology 5:505-10)。然而,还包括在溶液中具有与正 常
囊泡结构不同的结构的组合物。例如,脂质可呈现胶束结构或仅作 为脂质分子的不均匀聚集体存在。还考虑到脂质转染胺(lipofectamine)
ꢀ-
核酸复合物。
[0244]
无论用于将外源核酸引入宿主细胞中或以其他方式使细胞暴露 于本发明的抑制剂的方法如何,为了证实重组dna序列在宿主细胞 中的存在,可进行多种测定。此类测定包括,例如,本领域技术人员 公知的“分子生物学”测定,诸如southern和northern印迹法、rt-pcr 和pcr;“生物化学”测定,诸如通过例如免疫学手段(elisa和western 印迹法)或通过本文所述的测定来检测特定肽的存在或不存在,以鉴 定落入本发明范围内的试剂。
[0245]
本发明进一步提供了包含编码tfp的核酸分子的载体。在一个 方面,tfp载体可直接转导到细胞例如t细胞中。在一个方面,该载 体是克隆或表达载体,例如载体,包括但不限于一种或多种质粒(例 如,表达质粒、克隆载体、微环、微型载体、双微染色体)、逆转录 病毒和慢病毒载体构建体。在一个方面,该载体能够在哺乳动物t细 胞中表达tfp构建体。在一个方面,该哺乳动物t细胞是人t细胞。t细胞的来源
[0246]
在扩充和遗传修饰之前,从受试者获得t细胞来源。术语“受试 者”旨在包括其中可引发免疫应答的活生物体(例如,哺乳动物)。受 试者的实例包括人、狗、猫、小鼠、大鼠及其转基因物种。t细胞可 从许多来源获得,包括外周血单个核细胞、骨髓、淋巴结组织、脐带 血、胸腺组织、来自感染部位的组织、腹水、胸腔积液、脾组织和肿 瘤。在本发明的某些方面,可使用本领域可获得的任何数目的t细胞 系。在本发明的某些方面,可使用本领域技术人员已知的任意数量的 技术如ficoll
tm
分离技术从收集自受试者的血液单位中获得t细胞。 在一个优选的方面,通过单采血液成分术获得来自个体的循环血液的 细胞。单采血液成分术产物通常含有淋巴细胞,包括t细胞、单核细 胞、粒细胞、b细胞、其他有核白细胞、红细胞和血小板。在一个方 面,可洗涤通过单采血液成分术收集的细胞以去除血浆部分并将细胞 置于适当的缓冲液或培养基中以供后续处理步骤。在本发明的一个方 面,用磷酸盐缓冲盐水(pbs)洗涤细胞。在另一个方面,洗涤溶液 缺乏钙并且可能缺乏镁,或者可能缺少许多(如果不是全部)二价阳 离子。在不存在钙的情况下的初始活化步骤可以导致放大的活化。如 本领域普通技术人员将容易理解的,洗涤步骤可以通过本领域技术人 员已知的方法完成,诸如通过根据制造商的说明使用半自动“流通”离 心机(例如,cobe 2991细胞处理器、baxter cytomate或haemoneticscell saver 5)。洗涤后,细胞可重新悬浮于多种生物相容性缓冲液, 诸如无ca、无mg的pbs、plasmalyte a,或者具有或不具有缓冲液 的其他盐溶液。或者,可去除单采血液成分术样品中不需要的组分, 并将细胞直接重新悬浮于培养基中。
[0247]
在一个方面,通过溶解红细胞和耗尽单核细胞从外周血淋巴细 胞分离t细胞,例如,通过percolltm梯度离心或通过逆流离心 淘析。可通过正选择或负选择技术进一步分离特定的t细胞亚群,诸 如cd3+、cd28+、cd4+、cd8+、cd45ra+和cd45ro+ t细胞。例 如,在一个方面,通过与抗cd3/抗cd28(例如,3x28)缀合珠如 dynabeads
tm
m-450cd3/cd28 t一起温育足以对所需的t细胞进 行正选择的时间段来分离t细胞。在一个方面,该时间段是约30分 钟。在另一个方面,该时间段的范围为30分钟至36小时或更长,以 及其间的所有整数值。在另一个方面,该时间段为至少1、2、3、4、 5或6小时。在另一个优选的方面,该时间段是10至24小时。在一 个方面,温育时间段是24小时。在与其他细胞类型相比t细胞较少 的任何情
况下,可使用更长的温育时间来分离t细胞,诸如从肿瘤组 织或免疫受损个体分离肿瘤浸润淋巴细胞(til)。此外,使用更长的 温育时间可提高cd8+ t细胞的捕获效率。因此,通过简单地缩短或 延长允许t细胞与cd3/cd28珠结合的时间和/或通过增大或减小珠 子与t细胞的比率(如本文进一步描述的),可在培养起始时或在该 过程中的其他时间点优先选择或不选择t细胞亚群。另外,通过增大 或减小珠子或其他表面上的抗cd3和/或抗cd28抗体的比率,可在 培养起始时或在其他所需时间点优先选择或不选择t细胞亚群。本领 域技术人员将认识到,在本发明的背景下还可使用多轮选择。在某些 方面,可能需要进行选择程序并在激活和扩充过程中使用“未选择的
”ꢀ
细胞。“未选择的”细胞也可以经受其他轮选择。
[0248]
通过负选择来富集t细胞群体可通过针对对于负选择的细胞而 言独特的表面标志物的抗体组合来实现。一种方法是经由负磁性免疫 粘附或流式细胞术的细胞分选和/或选择,其使用针对存在于负选择的 细胞上的细胞表面标志物的单克隆抗体的混合物(cocktail)。例如, 为了通过负选择富集cd4+细胞,单克隆抗体混合物通常包含针对 cd14、cd20、cd11b、cd16、hla-dr和cd8的抗体。在某些方面, 可能需要富集或正选择通常表达cd4+、cd25+、cd62lhi、gitr+和 foxp3+的调节t细胞。或者,在某些方面,t调节细胞被抗c25缀合 的珠子或其他类似的选择方法耗尽。
[0249]
在一个实施方案中,可选择表达ifn-γ、tnf-α、il-17a、il-2、 il-3、il-4、gm-csf、il-10、il-13、粒酶b和穿孔蛋白或其他适当 的分子例如其他细胞因子中的一种或多种的t细胞群体。针对细胞表 达进行筛选的方法可以通过例如pct公开号wo2013/126712中描述 的方法确定。
[0250]
为了通过正选择或负选择分离所需的细胞群体,可改变细胞和 表面(例如颗粒,如珠子)的浓度。在某些方面,可能需要显著减小 珠子和细胞混合在一起的体积(例如,提高细胞浓度),以确保细胞 与珠子的最大接触。例如,在一个方面,使用20亿个细胞/ml的浓 度。在一个方面,使用10亿个细胞/ml的浓度。在另一个方面,使用 大于1亿个细胞/ml。在另一个方面,使用1000万、1500万、2000 万、2500万、3000万、3500万、4000万、4500万或5000万个细胞 /ml的细胞浓度。在又一个方面,使用7500万、8000万、8500万、 9000万、9500万或1亿个细胞/ml的细胞浓度。在其他方面,可使 用1.25亿或1.5亿个细胞/ml的浓度。使用高浓度可导致细胞产率、 细胞活化和细胞扩充的增加。此外,使用高细胞浓度可允许更有效地 捕获可能较弱地表达感兴趣的靶抗原的细胞,诸如cd28阴性t细胞, 或者来自存在许多肿瘤细胞的样品(例如,白血病血液、肿瘤组织等) 的细胞。这样的细胞群体可能具有治疗价值并且会希望获得。例如, 使用高浓度的细胞允许更有效地选择通常具有较弱cd28表达的 cd8+ t细胞。
[0251]
在相关方面,可能需要使用较低浓度的细胞。通过大量稀释t 细胞和表面(例如颗粒,如珠子)的混合物,使颗粒与细胞之间的相 互作用最小化。这选择表达较高量的所需抗原的细胞以与颗粒结合。 例如,cd4+ t细胞表达较高水平的cd28,并且在稀释浓度下比cd8+ t细胞更有效地被捕获。在一个方面,使用的细胞浓度为5x106个/ml。 在其他方面,使用的浓度可以是约1x105个/ml至1x106个/ml,以及 其间的任何整数值。在其他方面,细胞可在2-10℃或室温下在旋转 器上以不同的速度温育不同的时间长度。
[0252]
用于刺激的t细胞也可在洗涤步骤后冷冻。不希望受理论约束, 冷冻和随后的解冻步骤通过去除细胞群体中的粒细胞和一定程度地 去除单核细胞提供更均一的产物。在
与蛋白激酶c活化剂(例如,苔藓抑制素)以及钙离子载体接触。为 了共刺激t细胞表面上的辅助分子,使用与该辅助分子结合的配体。 例如,在适合于刺激t细胞增殖的条件下,可使t细胞群体与抗cd3 抗体和抗cd28抗体接触。为了刺激cd4+ t细胞或cd8+ t细胞的 增殖,使用抗cd3抗体和抗cd28抗体。抗cd28抗体的实例包括 9.3、b-t3、xr-cd28(diaclone,besancon,france),其可如本领域 公知的其他方法使用(berg等人,transplant proc.30(8):3975-3977, 1998;haanen等人,j.exp.med.190(9):13191328,1999;garland等人, j.immunol.meth.227(1-2):53-63,1999)。
[0257]
已暴露不同刺激时间的t细胞可表现出不同的特性。例如,典 型的血液或进行单采血液成分术的(apheresed)外周血单个核细胞产 物具有辅助t细胞群体(th,cd4+),其大于细胞毒性或抑制t细 胞群体(tc,cd8+)。通过刺激cd3和cd28受体的t细胞离体扩 充产生t细胞群体,其在约8-9天之前主要由th细胞组成,但在约 8-9天之后,该t细胞群体包含越来越大的tc细胞群体。因此,取 决于治疗目的,向受试者输注主要包含th细胞的t细胞群体可能是 有利的。类似地,如果已经分离了tc细胞的抗原特异性子集,则将 该子集扩充到更大程度可能是有益的。
[0258]
此外,除了cd4和cd8标志物之外,其他表型标志物也显著 变化,但在很大程度上,在细胞扩充过程中可再现地变化。因此,这 样的再现性实现了针对特定目的定制活化t细胞产物的能力。
[0259]
一旦构建了抗间皮素tfp,就可使用各种测定来评价分子的活 性,例如但不限于在抗原刺激后扩充t细胞的能力、在不存在再刺激 的情况下维持t细胞扩充的能力以及在适当的体外和动物模型中的 抗癌活性。用来评价抗间皮素tfp的作用的测定在下文进一步详细描 述。
[0260]
原代t细胞中tfp表达的western印迹分析可用于检测单体和 二聚体的存在(参见例如,milone等人,molecular therapy 17(8):1453
-ꢀ
1464(2009))。简言之,将表达tfp的t细胞(cd4
+
和cd8
+ t细胞 的1:1混合物)在体外扩充超过10天,然后在还原条件下进行溶解和 sds-page。使用针对tcr链的抗体通过western印迹法检测tfp。 使用相同的t细胞亚组在非还原条件下进行sds-page分析,以允 许评价共价二聚体形成。
[0261]
可通过流式细胞术测量在抗原刺激后tfp
+ t细胞的体外扩充。 例如,用αcd3/αcd28和apc刺激cd4
+
和cd8
+ t细胞的混合物, 然后在待分析的启动子的控制下用表达gfp的慢病毒载体进行转导。 示例性启动子包括cmv ie基因、ef-1α、遍在蛋白c或磷酸甘油激 酶(pgk)启动子。通过流式细胞术在cd4+和/或cd8+ t细胞子集 中在培养的第6天评估gfp荧光(参见例如,milone等人,moleculartherapy 17(8):1453-1464(2009))。或者,在第0天用αcd3/αcd28 包被的磁珠刺激cd4+和cd8+ t细胞的混合物,并使用表达tfp的 双顺反子慢病毒载体和使用2a核糖体跳跃序列的egfp在第1天用 tfp转导。在洗涤后,在抗cd3和抗cd28抗体(k562-bbl-3/28) 的存在下,用间皮素+k562细胞(k562-间皮素)、野生型k562细 胞(k562野生型)或表达hcd32和4-1bbl的k562细胞再刺激培 养物。每隔一天以100iu/ml向培养物添加外源il-2。使用基于珠子 的计数通过流式细胞术对gfp+ t细胞进行计数(参见例如,milone 等人,molecular therapy 17(8):1453-1464(2009))。
[0262]
还可测量在不存在再刺激的情况下持续的tfp+ t细胞扩充(参 见例如,milone等人,molecular therapy 17(8):1453-1464(2009))。 简言之,在第0天用αcd3/αcd28包被的
磁珠刺激,并在第1天用指 定的tfp转导后,使用coulter multisizer iii颗粒计数器在培养的第8天测量平均t细胞体积(fl)。
[0263]
动物模型也可用于测量tfp-t活性。例如,使用人间皮素特异 性tfp+ t细胞来治疗免疫缺陷小鼠中的癌症的异种移植模型(参见 例如,milone等人,molecular therapy 17(8):1453-1464(2009))。简 言之,在建立癌症后,将小鼠随机分配至治疗组。不同数目的工程化 t细胞以1:1的比例共同注射到携带癌症的nod/scid/γ-/-小鼠中。 在t细胞注射后的不同时间评估来自小鼠的脾dna中每种载体的拷 贝数。以每周的间隔评估动物的癌症。在注射了α间皮素-ζtfp+ t细 胞或模拟转导的t细胞的小鼠中测量外周血间皮素+癌细胞计数。使 用对数秩检验比较各组的存活曲线。此外,还可分析在nod/scid/γ
-ꢀ
/-小鼠中注射t细胞后4周的绝对外周血cd4+和cd8+ t细胞计数。 向小鼠注射癌细胞,并在3周后注射t细胞,该t细胞被工程化以通 过编码与egfp连接的tfp的双顺反子慢病毒载体而表达tfp。通过 在注射前与模拟转导的细胞混合,将t细胞归一化为45-50%输入 gfp+ t细胞,并通过流式细胞术确认。以1周的间隔评估动物的癌 症。使用对数秩检验比较tfp+ t细胞组的存活曲线。
[0264]
可评估剂量依赖性tfp治疗响应(参见例如,milone等人, molecular therapy 17(8):1453-1464(2009))。例如,在第21天用tfpt细胞、等量的模拟转导的t细胞或没有t细胞注射的小鼠中建立癌 症后35-70天获得外周血。将来自每组的小鼠在第35天和第49天随 机放血以确定外周血间皮素+癌细胞计数,然后杀死。在第57天和第 70天评估剩余的动物。
[0265]
先前已经描述了细胞增殖和细胞因子产生的评估,例如,milone 等人,molecular therapy 17(8):1453-1464(2009)。简言之,通过将洗 涤的t细胞与表达间皮素或cd32和cd137的细胞(kt32-bbl)混 合,最终的t细胞:表达间皮素的细胞之比为2:1,在微量滴定板中进 行tfp介导的增殖的评估。表达间皮素的细胞在使用前用γ射线照 射。将抗cd3(克隆okt3)和抗cd28(克隆9.3)单克隆抗体添加 至具有kt32-bbl细胞的培养物中,以充当刺激t细胞增殖的阳性对 照,因为这些信号支持离体的长期cd8+ t细胞扩充。如制造商所述, 使用countbright
tm
荧光珠(invitrogen)和流式细胞术对培养物中的t 细胞进行计数。使用用egfp-2a连接的表达tfp的慢病毒载体工程 化的t细胞,通过gfp表达鉴定tfp+ t细胞。对于不表达gfp的 tfp+ t细胞,用生物素化的重组间皮素蛋白质和次级抗生物素蛋白
-ꢀ
pe缀合物检测tfp+ t细胞。还用特异性单克隆抗体(bd biosciences) 同时检测t细胞上的cd4+和cd8+表达。根据制造商的说明,使用 人th1/th2细胞因子细胞计数珠阵列试剂盒(bd biosciences)对再 刺激后24小时收集的上清液进行细胞因子测量。使用facscalibur流 式细胞仪评估荧光,并根据制造商的说明分析数据。
[0266]
可通过标准
51
cr释放测定来评估细胞毒性(参见例如,milone 等人,molecular therapy 17(8):1453-1464(2009))。简言之,在37℃ 下用
51
cr(作为nacro4,new england nuclear)加载靶细胞,在频繁 搅拌下持续2小时,在完全rpmi培养基中洗涤两次,并接种到微量 滴定板中。将效应t细胞与孔中完全rpmi中的靶细胞以不同的效应 细胞:靶细胞(e:t)比率混合。还准备了仅含有培养基(自发释放, sr)或triton-x 100去污剂的1%溶液(总释放,tr)的额外孔。在 37℃下温育4小时后,收获每个孔的上清液。然后使用γ粒子计数器 (packard instrument co.,waltham,mass.)测量释放的
51
cr。每种条 件至少一式三份进行,并使用下式计算溶解百分比:%溶解=(er
-ꢀ
sr)/(tr-sr),其中er表示每种实验条件释
放的平均
51
cr。
[0267]
可使用成像技术评价携带肿瘤的动物模型中tfp的特定运输和 增殖。这样的测定已描述于例如barrett等人,human gene therapy 22:1575-1586(2011)。简言之,向nod/scid/γc-/-(nsg)小鼠静脉 内注射癌细胞,7天后在用tfp构建体电穿孔后4小时注射t细胞。 用慢病毒构建体稳定转染t细胞以表达萤火虫萤光素酶,并对小鼠进 行生物发光成像。或者,可如下测量在癌症异种移植模型中单次注射 tfp+ t细胞的治疗功效和特异性:向nsg小鼠注射被转导以稳定表 达萤火虫萤光素酶的癌细胞,然后在7天后单次尾静脉注射用间皮素tfp电穿孔的t细胞。在注射后的不同时间点对动物进行成像。例如, 可产生第5天(治疗前2天)和第8天(tfp+pbl后24小时)代表 性小鼠中的萤火虫萤光素酶阳性癌症的光子密度热图。
[0268]
其他测定,包括本文实施例部分中描述的测定以及本领域已知 的测定,也可用于评价本发明的抗间皮素tfp构建体。
[0269]
治疗应用间皮素相关疾病和/或病症
[0270]
在一个方面,本发明提供了治疗与间皮素表达相关的疾病的方 法。在一个方面,本发明提供了用于治疗疾病的方法,其中部分肿瘤 对间皮素呈阴性,而部分肿瘤对间皮素呈阳性。例如,本发明的tfp 可用于治疗已经经历与间皮素表达升高相关的疾病的治疗的受试者, 其中已经经历间皮素水平升高的治疗的受试者表现出与间皮素水平 升高相关的疾病。
[0271]
在一个方面,本发明涉及包含抗间皮素tfp的载体,该抗间皮 素tfp与用于在哺乳动物t细胞中表达的启动子可操作地连接。在 一个方面,本发明提供了表达间皮素tfp以用于治疗表达间皮素的肿 瘤的重组t细胞,其中表达间皮素tfp的重组t细胞被称为间皮素 tfp-t。在一个方面,本发明的间皮素tfp-t能够使肿瘤细胞与在其 表面上表达的至少一种本发明的间皮素tfp接触,使得tfp-t靶向 肿瘤细胞并抑制肿瘤的生长。
[0272]
在一个方面,本发明涉及抑制表达间皮素的肿瘤细胞的生长的 方法,其包括使肿瘤细胞与本发明的间皮素tfp t细胞接触,使得 tfp-t响应于抗原被激活并靶向癌细胞,其中肿瘤的生长得到抑制。
[0273]
在一个方面,本发明涉及治疗受试者中的癌症的方法。该方法 包括向受试者施用本发明的间皮素tfp t细胞,使得在受试者中治疗 癌症。可以用本发明的间皮素tfp t细胞治疗的癌症的实例是与间皮 素表达相关的癌症。在一个方面,该癌症是间皮瘤。在一个方面,该 癌症是胰腺癌。在一个方面,该癌症是卵巢癌。在一个方面,该癌症 是胃癌。在一个方面,该癌症是肺癌。在一个方面,该癌症是子宫内 膜癌。在一些实施方案中,间皮素tfp疗法可与一种或多种另外的疗 法联合使用。
[0274]
本发明包括如下类型的细胞疗法,其中t细胞经遗传修饰以表 达tfp,并且将表达tfp的t细胞输注给有需要的接受者。所输注的 细胞能够杀伤接受者中的肿瘤细胞。与抗体疗法不同,表达tfp的t 细胞能够在体内复制,从而产生长期持久性,这可导致持续的肿瘤控 制。在各个方面,在将t细胞施用于患者后,施用于患者的t细胞或 其后代在患者中存留至少一个月、两个月、三个月、四个月、五个月、 六个月、七个月、八个月、九个月、十个月、十一个月、十二个月、 十三个月、十四个月、十五个月、十六个月、十七个月、十八个月、 十九
个月、二十个月、二十一个月、二十二个月、二十三个月、两年、 三年、四年或五年。
[0275]
本发明还包括如下类型的细胞疗法,其中例如用体外转录的 rna修饰t细胞以瞬时表达tfp,并且将表达tfp的t细胞输注给 有需要的接受者。所输注的细胞能够杀伤接受者中的肿瘤细胞。因此, 在各个方面,在将t细胞施用于患者后,施用于患者的t细胞存在少 于一个月,例如三周、两周或一周。
[0276]
不希望受任何特定理论的约束,由表达tfp的t细胞引发的抗 肿瘤免疫应答可以是主动或被动免疫应答,或者可归于直接与间接免 疫应答。在一个方面,tfp转导的t细胞响应于表达间皮素抗原的人 癌细胞表现出特异性促炎细胞因子分泌和有效的细胞溶解活性,抵抗 可溶性间皮素抑制,介导旁观者杀伤,和/或介导已建立的人肿瘤的消 退。例如,表达间皮素的肿瘤的异质区域内的无抗原肿瘤细胞可能易 受间皮素重定向的t细胞的间接破坏,该t细胞先前已经与邻近的抗 原阳性癌细胞反应。
[0277]
在一个方面,本发明的人tfp修饰的t细胞可以是用于哺乳动 物中的离体免疫和/或体内治疗的疫苗类型。在一个方面,该哺乳动物 是人。
[0278]
关于离体免疫,在将细胞施用于哺乳动物之前,在体外发生以 下至少一种:i)细胞扩充,ii)将编码tfp的核酸引入细胞,或iii) 冷冻保存细胞。
[0279]
离体程序是本领域公知的,并在以下更充分地讨论。简言之, 从哺乳动物(例如,人)分离细胞,并用表达本文公开的tfp的载体 对其进行遗传修饰(即,体外转导或转染)。tfp修饰的细胞可施用 于哺乳动物接受者以提供治疗益处。哺乳动物接受者可以是人,并且 tfp修饰的细胞相对于接受者可以是自体的。或者,该细胞相对于接 受者可以是同种异体的、同种同基因的或异种的。
[0280]
用于离体扩充造血干细胞和祖细胞的程序描述于通过引用并入 本文的美国专利号5,199,942中,可应用于本发明的细胞。其他合适 的方法是本领域已知的,因此本发明不限于离体扩充细胞的任何特定 方法。简言之,t细胞的离体培养和扩充包括:(1)从外周血收获物 或骨髓外植体收集来自哺乳动物的cd34+造血干细胞和祖细胞;(2) 离体扩充这类细胞。除了美国专利号5,199,942中描述的细胞生长因 子之外,其他因子如flt3-l、il-1、il-3和c-kit配体也可用于培养和 扩充细胞。
[0281]
除了在离体免疫方面使用基于细胞的疫苗之外,本发明还提供 用于体内免疫的组合物和方法,以在患者中引发针对抗原的免疫应答。
[0282]
通常,如本文所述活化和扩充的细胞可用于治疗和预防免疫受 损个体中出现的疾病。特别地,本发明的tfp修饰的t细胞用于治疗 与间皮素表达相关的疾病、病症和病况。在某些方面,本发明的细胞 用于治疗有患与间皮素表达相关的疾病、病症和病况的风险的患者。 因此,本发明提供了用于治疗或预防与间皮素表达相关的疾病、病症 和病况的方法,其包括向有需要的受试者施用治疗有效量的本发明的 tfp修饰的t细胞。
[0283]
在一个方面,本发明的tfp-t细胞可用于治疗增生性疾病,诸 如癌症或恶性肿瘤或癌前病况。在一个方面,该癌症是间皮瘤。在一 个方面,该癌症是胰腺癌。在一个方面,该癌症是卵巢癌。在一个方 面,该癌症是胃癌。在一个方面,该癌症是肺癌。在一个方面,该癌 症是子宫内膜癌。此外,与间皮素表达相关的疾病包括但不限于例如 非典型和/或非经典癌症、恶性肿瘤、癌前病况或表达间皮素的增生性 疾病。与间皮素表达相关的非癌症相关适应症包括但不限于例如自身 免疫病(例如狼疮)、炎性病症(变态反应和哮喘)和移植。
[0284]
本发明的tfp修饰的t细胞可单独施用,或作为药物组合物与 稀释剂和/或与其他组分如il-2或其他细胞因子或细胞群体联合施用。
[0285]
本发明还提供了用于减少表达间皮素的细胞群体或抑制其增殖 的方法,该方法包括使包含表达间皮素的细胞的细胞群体与本发明的 与表达间皮素的细胞结合的抗间皮素tfp-t细胞接触。在一个具体方 面,本发明提供了用于减少表达间皮素的癌细胞群体或抑制其增殖的 方法,该方法包括使表达间皮素的癌细胞群体与本发明的与表达间皮 素的细胞结合的抗间皮素tfp-t细胞结合。在一个方面,本发明提供 了用于减少表达间皮素的癌细胞群体或抑制其增殖的方法,该方法包 括使表达间皮素的癌细胞群体与本发明的与表达间皮素的细胞结合 的抗间皮素tfp-t细胞结合。在某些方面,相对于阴性对照,在具有 与表达间皮素的细胞相关的癌症的受试者或动物模型中,本发明的抗 间皮素tfp-t细胞使细胞和/或癌细胞的数量、数目、量或百分比减 少至少25%、至少30%、至少40%、至少50%、至少65%、至少75%、 至少85%、至少95%或至少99%。在一个方面,受试者是人。
[0286]
本发明还提供了用于预防、治疗和/或管理与表达间皮素的细胞 (例如,表达间皮素的癌症)相关的疾病的方法,该方法包括向有需 要的受试者施用本发明的与表达间皮素的细胞结合的抗间皮素tfp
-ꢀ
t细胞。在一个方面,受试者是人。与表达间皮素的细胞相关的病症 的非限制性实例包括自身免疫病症(诸如狼疮)、炎性病症(诸如变 态反应和哮喘)和癌症(诸如胰腺癌、卵巢癌、胃癌、肺癌或子宫内 膜癌,或表达间皮素的非典型癌症)。
[0287]
本发明还提供了用于预防、治疗和/或管理与表达间皮素的细胞 相关的疾病的方法,该方法包括向有需要的受试者施用本发明的与表 达间皮素的细胞结合的抗间皮素tfp-t细胞。在一个方面,受试者是 人。
[0288]
本发明提供了用于预防与表达间皮素的细胞相关的癌症的复发 的方法,该方法包括向有需要的受试者施用本发明的与表达间皮素的 细胞结合的抗间皮素tfp-t细胞。在一个方面,该方法包括向有需要 的受试者施用有效量的本文所述的与表达间皮素-bmca的细胞结合的 抗间皮素tfp-t细胞,联合有效量的另一种疗法。联合疗法
[0289]
本文所述的表达tfp的细胞可与其他已知的药剂和疗法联合使 用。如本文所用,“联合”施用意指在受试者罹患病症的过程中向受试 者递送两种(或更多种)不同的治疗,例如,在受试者被诊断为患有 该病症后并在该病症被治愈或消除之前或者由于其他原因停止治疗 之前递送两种或更多种治疗。在一些实施方案中,一种治疗的递送在 第二种治疗的递送开始时仍在进行,因此在施用方面存在重叠。这在 本文中有时称为“同时”或“并行递送”。在其他实施方案中,一种治疗 的递送在另一种治疗的递送开始之前结束。在任一种情况的一些实施 方案中,由于联合施用,治疗更加有效。例如,相比于在不存在第一 种治疗的情况下施用第二种治疗的情况将观察到的,第二种治疗更有 效,例如,使用更少的第二种治疗观察到同等效果,或者第二种治疗 在更大程度上减轻症状,或者对第一种治疗观察到类似情况。在一些 实施方案中,递送使得症状或与病症相关的其他参数的减小大于在不 存在另一种治疗的情况下递送的一种治疗将观察到的。两种治疗的效 果可以是部分相加、完全相加或大于相加。这种递送可使得所递送的 第一种治疗的效果在递送第二种治疗时仍可检测。
[0290]
在一些实施方案中,“至少一种另外的治疗剂”包括表达tfp的 细胞。还提供了表
达多种tfp的t细胞,其与相同或不同的靶抗原或 者相同靶抗原上的相同或不同的表位结合。还提供了t细胞群体,其 中t细胞的第一子集表达第一tfp,t细胞的第二子集表达第二tfp。
[0291]
本文所述的表达tfp的细胞和至少一种另外的治疗剂可在相同 或单独的组合物中同时施用,或者顺序地施用。对于顺序施用,可首 先施用本文所述的表达tfp的细胞,并可其次施用另外的药剂,或者 施用顺序可逆转。
[0292]
在其他方面,本文所述的表达tfp的细胞可在治疗方案中与手 术、化疗、放疗、免疫抑制剂(如环孢菌素、硫唑嘌呤、甲氨蝶呤、 霉酚酸酯和fk506)、抗体或其他免疫消融剂如阿仑珠单抗、抗cd3 抗体或其他抗体疗法、细胞毒素、氟达拉滨、环孢菌素、fk506、雷 帕霉素、霉酚酸、类固醇、fr901228、细胞因子和照射联合使用。本 文所述的表达tfp的细胞还可与肽疫苗联合使用,诸如izumoto等人 2008j neurosurg 108:963-971所述的肽疫苗。在另一个方面,本文所 述的表达tfp的细胞还可与髓样细胞分化的启动子(例如,全反式视 黄酸)、骨髓衍生的抑制细胞(mdsc)扩充的抑制剂(例如,c-kit受 体抑制剂或vegf抑制剂)、mdsc功能的抑制(例如,cox2抑制 剂或磷酸二酯酶-5抑制剂)或mdsc的治疗性消除(例如,用化疗方 案,诸如用多柔比星和环磷酰胺治疗)联合使用。可阻止mdsc扩充 的其他治疗剂包括氨基二膦酸盐、二膦酸盐、西地那非和他达拉非、 硝基阿司匹林、维生素d3和吉西他滨(参见例如,gabrilovich和 nagaraj,nat.rev.immunol,(2009)v9(3):162-174)。
[0293]
在一个实施方案中,可向受试者施用减少或改善与施用tfp表 达细胞相关的副作用的药剂。与施用tfp表达细胞相关的副作用包括 但不限于细胞因子释放综合征(crs)和噬血细胞性淋巴组织细胞增 多症(hlh),也称为巨噬细胞活化综合征(mas)。crs的症状包 括高烧、恶心、短暂性低血压、缺氧等。因此,本文所述的方法可包 括向受试者施用本文所述的表达tfp的细胞,并进一步施用药剂以管 理由采用表达tfp的细胞治疗导致的可溶性因子的水平升高。在一个 实施方案中,受试者中升高的可溶性因子是ifn-γ、tnfα、il-2、il
-ꢀ
6和il8中的一种或多种。因此,为了治疗该副作用而施用的药剂可 以是中和这些可溶性因子中的一种或多种的药剂。此类药剂包括但不 限于类固醇、tnfα抑制剂和il-6抑制剂。tnfα抑制剂的实例是依 那西普(entanercept)。il-6抑制剂的实例是托珠单抗(toc)。
[0294]
在一个实施方案中,可向受试者施用增强表达tfp的细胞的活 性的药剂。例如,在一个实施方案中,该药剂可以是抑制抑制性分子 的药剂。在一些实施方案中,抑制性分子,例如程序性死亡1(pd1), 可降低表达tfp的细胞产生免疫效应物应答的能力。抑制性分子的实 例包括pd1、pd-l1、ctla4、tim3、lag3、vista、btla、tigit、 lair1、cd160、2b4和tgfrβ。抑制性分子的抑制,例如通过在 dna/rna或蛋白质水平上的抑制,可优化表达tfp的细胞的性能。 在实施方案中,抑制性核酸,例如抑制性核酸,例如dsrna,例如 sirna或shrna,可用于抑制表达tfp的细胞中抑制性分子的表达。 在一个实施方案中,抑制剂是shrna。在一个实施方案中,抑制性分 子在表达tfp的细胞内受到抑制。在这些实施方案中,抑制抑制性分 子表达的dsrna分子与编码tfp的组分例如所有组分的核酸连接。 在一个实施方案中,抑制性信号的抑制剂可以是例如与抑制性分子结 合的抗体或抗体片段。例如,该药剂可以是与pd1、pd-l1、pd-l2或 ctla4结合的抗体或抗体片段(例如,伊匹木单抗(ipilimumab)(也 称为mdx-010和mdx-101,并且以yervoy
tm
销售;bristol-myerssquibb;曲美木单抗(tremelimumab)(可从pfizer获得的igg2单克 隆抗体,旧称ticilimumab,cp-675,206)。在
一个实施方案中,该 药剂是与tim3结合的抗体或抗体片段。在一个实施方案中,该药剂 是与lag3结合的抗体或抗体片段。
[0295]
在一些实施方案中,可通过慢病毒(例如,特异性靶向cd4+或 cd8+ t细胞的慢病毒)在体内改变(例如,通过基因转移)t细胞 (参见例如,zhou等人,j.immunol.(2015)195:2493-2501)。
[0296]
在一些实施方案中,增强表达tfp的细胞的活性的试剂可以是 例如包含第一结构域和第二结构域的融合蛋白,其中第一结构域是抑 制性分子或其片段,并且第二结构域是与正信号相关的多肽,例如包 含如本文所述的细胞内信号传导结构域的多肽。在一些实施方案中, 与正信号相关的多肽可包括cd28、cd27、icos的共刺激结构域, 例如cd28、cd27和/或icos的细胞内信号传导结构域,和/或初级 信号传导结构域,例如cd3ζ的初级信号传导结构域,例如本文所述 的初级信号传导结构域。在一个实施方案中,该融合蛋白由表达tfp 的相同细胞表达。在另一个实施方案中,该融合蛋白由细胞表达,例 如由不表达抗间皮素tfp的t细胞表达。
[0297]
在一些实施方案中,包含作为由核酸编码的抗间皮素结合结构 域的抗原结合结构域的人或人源化抗体结构域,或包含抗间皮素结合 结构域的抗体,或表达由核酸编码的抗间皮素结合结构域的细胞,具 有以下亲和力值:至多约200nm、100nm、75nm、50nm、25nm、 20nm、15nm、14nm、13nm、12nm、11nm、10nm、9nm、8nm、 7nm、6nm、5nm、4nm、3nm、2nm、1nm、0.9nm、0.8nm、 0.7nm、0.6nm、0.5nm、0.4nm、0.3nm、0.2nm、0.1nm、0.09nm、 0.08nm、0.07nm、0.06nm、0.05nm、0.04nm、0.03nm、0.02nm 或0.01nm;和/或至少约100nm、75nm、50nm、25nm、20nm、 15nm、14nm、13nm、12nm、11nm、10nm、9nm、8nm、7nm、 6nm、5nm、4nm、3nm、2nm、1nm、0.9nm、0.8nm、0.7nm、 0.6nm、0.5nm、0.4nm、0.3nm、0.2nm、0.1nm、0.09nm、0.08 nm、0.07nm、0.06nm、0.05nm、0.04nm、0.03nm、0.02nm或0.01 nm;和/或约200nm、100nm、75nm、50nm、25nm、20nm、15 nm、14nm、13nm、12nm、11nm、10nm、9nm、8nm、7nm、 6nm、5nm、4nm、3nm、2nm、1nm、0.9nm、0.8nm、0.7nm、 0.6nm、0.5nm、0.4nm、0.3nm、0.2nm、0.1nm、0.09nm、0.08 nm、0.07nm、0.06nm、0.05nm、0.04nm、0.03nm、0.02nm或0.01 nm。药物组合物
[0298]
本发明的药物组合物可包含如本文所述的表达tfp的细胞,例 如,多种表达tfp的细胞,联合一种或多种药学上或生理学上可接受 的载体、稀释剂或赋形剂。此类组合物可包含缓冲液,如中性缓冲盐 水、磷酸盐缓冲盐水等;碳水化合物,如葡萄糖、甘露糖、蔗糖或葡 聚糖、甘露醇;蛋白质;多肽或氨基酸,如甘氨酸;抗氧化剂;螯合 剂,如edta或谷胱甘肽;佐剂(例如,氢氧化铝);以及防腐剂。 在一个方面,本发明的组合物被配制用于静脉内给药。
[0299]
本发明的药物组合物可以以适合于待治疗(或预防)的疾病的 方式施用。施用的量和频率将取决于诸如患者的状况以及患者疾病的 类型和严重程度等因素,然而适当的剂量可通过临床试验确定。
[0300]
在一个实施方案中,所述药物组合物基本上不含(例如,没有 可检测水平的)污染物,例如,选自内毒素、支原体、有复制能力的 慢病毒(rcl)、p24、vsv-g核酸、hiv gag、残留的抗cd3/抗cd28 包被的珠子、小鼠抗体、合并的人血清、牛血清白蛋白、牛血清、培 养基组分、载体包装细胞或质粒组分、细菌和真菌的污染物。在一个 实施方案中,该细菌是选自下
组的至少一种:粪产碱菌(alcaligenesfaecalis)、白色念珠菌(candida albicans)、大肠杆菌(escherichiacoli)、流感嗜血菌(haemophilus influenza)、脑膜炎奈瑟氏球菌 (neisseria meningitides)、铜绿假单胞菌(pseudomonas aeruginosa)、 金黄色葡萄球菌(staphylococcus aureus)、肺炎链球菌(streptococcuspneumonia)和酿脓链球菌(streptococcus pyogenes)组a。
[0301]
当指出“免疫有效量”、“抗肿瘤有效量”、“肿瘤抑制有效量”或
ꢀ“
治疗量”时,待施用的本发明组合物的精确量可由医生在考虑年龄、 体重、肿瘤大小、感染或转移程度以及患者(受试者)状况的个体差 异的情况下确定。通常,可规定包含本文所述t细胞的药物组合物可 以以104至109个细胞/kg体重的剂量施用,在一些情况下为105至106个细胞/kg体重,包括这些范围内的所有整数值。t细胞组合物还可 以以这些剂量多次施用。可通过使用免疫疗法中公知的输注技术来施 用细胞(参见例如,rosenberg等人,new eng.j.of med.319:1676, 1988)。
[0302]
在某些方面,可能需要将活化的t细胞施用于受试者,随后重 新抽取血液(或进行单采血液成分术),根据本发明从其激活t细胞, 并将这些活化和扩充的t细胞重新输注给患者。该过程可每隔几周进 行多次。在某些方面,t细胞可从10cc至400cc的抽取血液活化。在 某些方面,t细胞从20cc、30cc、40cc、50cc、60cc、70cc、80cc、90cc 或100cc的抽取血液活化。
[0303]
主题组合物的施用可以以任何方便的方式进行,包括通过气雾 剂吸入、注射、摄入、输液、植入或移植。本文所述的组合物可经动 脉、皮下、皮内、肿瘤内、结内、髓内、肌肉内、通过静脉内(i.v.) 注射或腹膜内施用于患者。在一个方面,通过皮内或皮下注射将本发 明的t细胞组合物施用于患者。在一个方面,本发明的t细胞组合物 通过静脉内注射施用。可以将t细胞的组合物直接注射到肿瘤、淋巴 结或感染部位中。
[0304]
在特定示例性方面,受试者可经受白细胞去除术,其中离体收 集、富集或耗尽白细胞以选择和/或分离感兴趣的细胞,例如t细胞。 这些t细胞分离物可通过本领域已知的方法扩充,并进行处理,使得 可以引入一种或多种本发明tfp构建体,从而产生本发明的表达tfp 的t细胞。有需要的受试者可随后经受高剂量化疗然后进行外周血干 细胞移植的标准治疗。在某些方面,在移植后或与移植同时,受试者 接受扩充的本发明tfp t细胞的输注。在另一个方面,在手术之前或 之后施用扩充的细胞。
[0305]
待施用于患者的上述治疗的剂量将随所治疗病况的精确性质和 治疗的接受者而变化。可根据本领域公认的实践来进行人类施用的剂 量的缩放。例如,对于成年患者,阿仑珠单抗的剂量将通常为1至约 100mg,通常每日施用持续1至30天。优选的日剂量为每日1至10 mg,但在一些情况下,可以使用至多每日40mg的较大剂量(描述于 美国专利号6,120,766中)。
[0306]
在一个实施方案中,使用例如体外转录将tfp引入t细胞中, 并且受试者(例如,人)接受本发明的tfp t细胞的初始施用,以及 本发明的tfp t细胞的一次或多次后续施用,其中所述一次或多次后 续施用在先前施用后少于15天,例如,14、13、12、11、10、9、8、 7、6、5、4、3或2天进行。在一个实施方案中,每周向受试者(例 如,人)进行本发明的tfp t细胞的多于一次施用,例如,每周进行 本发明的tfp t细胞的2、3或4次施用。在一个实施方案中,受试 者(例如,人类受试者)接受每周多于一次tfp t细胞施用(例如, 每周2、3或4次施
用)(本文中也称为周期),然后一周没有tfp t 细胞施用,然后向受试者施用一次或多次另外的tfp t细胞施用(例 如,每周多于一次tfp t细胞施用)。在另一个实施方案中,受试者 (例如,人类受试者)接受多于一个周期的tfp t细胞,并且每个周 期之间的时间少于10、9、8、7、6、5、4或3天。在一个实施方案 中,tfp t细胞每隔一天施用,每周施用3次。在一个实施方案中, 本发明的tfp t细胞施用至少2、3、4、5、6、7、8周或更多周。
[0307]
在一个方面,使用慢病毒病毒载体如慢病毒产生间皮素tfp t 细胞。以这种方式产生的tfp-t细胞将具有稳定的tfp表达。
[0308]
在一个方面,tfp t细胞在转导后瞬时表达tfp载体,持续4、 5、6、7、8、9、10、11、12、13、14、15天。tfp的瞬时表达可通 过rna tfp载体递送来实现。在一个方面,通过电穿孔将tfp rna 转导到t细胞中。
[0309]
在使用瞬时表达tfp t细胞(特别是携带鼠scfv的tfp t细 胞)治疗的患者中,可出现的潜在问题是多次治疗后的过敏反应。
[0310]
不受该理论约束,据认为这样的过敏反应可能由患者发生体液 抗tfp应答,即具有抗ige同种型的抗tfp抗体引起。据认为,当 暴露于抗原时具有十至十四天中断时,患者产生抗体的细胞经历从 igg同种型(不引起过敏反应)到ige同种型的类别转换。
[0311]
如果患者在瞬时tfp治疗(诸如通过rna转导产生的那些) 过程中具有产生抗tfp抗体应答的高风险,则tfp t细胞输注中断 不应当持续超过十至十四天。本发明提供了包括但不限于以下实施方案:1.一种编码t细胞受体(tcr)融合蛋白(tfp)的分离的重组 核酸分子,所述tfp包含:(a)tcr亚单位,其包含:(i)tcr细胞外结构域的至少一部分,和(ii)包含来自cd3ε的细胞内信号传导结构域的刺激结构域 的tcr细胞内结构域;以及(b)包含抗原结合结构域的人或人源化抗体结构域,所述抗原结 合结构域是抗间皮素结合结构域;其中所述tcr亚单位和所述抗体结构域可操作地连接,并且其中所述tfp当在t细胞中表达时并入tcr中。2.一种编码t细胞受体(tcr)融合蛋白(tfp)的分离的重组 核酸分子,所述tfp包含:(a)tcr亚单位,其包含:(i)tcr细胞外结构域的至少一部分,和(ii)包含来自cd3ε的细胞内信号传导结构域的刺激结构域 的tcr细胞内结构域;以及(b)包含抗原结合结构域的人或人源化抗体结构域,所述抗原结 合结构域是抗间皮素结合结构域;其中所述tcr亚单位和所述抗体结构域可操作地连接,并且其中所述tfp当在t细胞中表达时并入tcr中。3.一种编码t细胞受体(tcr)融合蛋白(tfp)的分离的重组 核酸分子,所述tfp包含:(a)tcr亚单位,其包含:(i)tcr细胞外结构域的至少一部分,和(ii)包含来自cd3γ的细胞内信号传导结构域的刺激结构域 的tcr细胞内结构域;以及
(b)包含抗原结合结构域的人或人源化抗体结构域,所述抗原结 合结构域是抗间皮素结合结构域;其中所述tcr亚单位和所述抗体结构域可操作地连接,并且其中所述tfp当在t细胞中表达时并入tcr中。4.一种编码t细胞受体(tcr)融合蛋白(tfp)的分离的重组 核酸分子,所述tfp包含:(a)tcr亚单位,其包含:(i)tcr细胞外结构域的至少一部分,和(ii)包含来自cd3δ的细胞内信号传导结构域的刺激结构域 的tcr细胞内结构域;以及(b)包含抗原结合结构域的人或人源化抗体结构域,所述抗原结 合结构域是抗间皮素结合结构域;其中所述tcr亚单位和所述抗体结构域可操作地连接,并且其中所述tfp当在t细胞中表达时并入tcr中。5.一种编码t细胞受体(tcr)融合蛋白(tfp)的分离的重组 核酸分子,所述tfp包含:(a)tcr亚单位,其包含:(i)tcr细胞外结构域的至少一部分,和(ii)包含来自tcrα的细胞内信号传导结构域的刺激结构域 的tcr细胞内结构域;以及(b)包含抗原结合结构域的人或人源化抗体结构域,所述抗原结 合结构域是抗间皮素结合结构域;其中所述tcr亚单位和所述抗体结构域可操作地连接,并且其中所述tfp当在t细胞中表达时并入tcr中。6.一种编码t细胞受体(tcr)融合蛋白(tfp)的分离的重组 核酸分子,所述tfp包含:(a)tcr亚单位,其包含:(i)tcr细胞外结构域的至少一部分,和(ii)包含来自tcrβ的细胞内信号传导结构域的刺激结构域 的tcr细胞内结构域;以及(b)包含抗原结合结构域的人或人源化抗体结构域,所述抗原结 合结构域是抗间皮素结合结构域;其中所述tcr亚单位和所述抗体结构域可操作地连接,并且其中所述tfp当在t细胞中表达时并入tcr中。7.一种编码t细胞受体(tcr)融合蛋白(tfp)的分离的重组 核酸分子,所述tfp包含tcr亚单位以及包含抗原结合结构域的人 或人源化抗体结构域,所述抗原结合结构域是抗间皮素结合结构域。8.如实施方案7所述的分离的核酸分子,其中所述tcr亚单位 和所述抗体结构域可操作地连接。9.如实施方案7或8所述的分离的核酸分子,其中所述tfp当 在t细胞中表达时并入tcr中。10.如实施方案1-9中任一项所述的分离的核酸分子,其中所编 码的抗原结合结构域通过连接体序列与所述tcr细胞外结构域连接。11.如实施方案10所述的分离的核酸分子,其中所编码的连接体 序列包括(g4s)
n
,其中n=1至4。
12.如实施方案1-11中任一项所述的分离的核酸分子,其中所述 tcr亚单位包含tcr细胞外结构域。13.如实施方案1-12中任一项所述的分离的核酸分子,其中所述 tcr亚单位包含tcr跨膜结构域。14.如实施方案1-13中任一项所述的分离的核酸分子,其中所述 tcr亚单位包含tcr细胞内结构域。15.如实施方案1-14中任一项所述的分离的核酸分子,其中所述 tcr亚单位包含(i)tcr细胞外结构域,(ii)tcr跨膜结构域,和 (iii)tcr细胞内结构域,其中(i)、(ii)和(iii)中的至少两个 来自相同的tcr亚单位。16.如实施方案1-15中任一项所述的分离的核酸分子,其中所述 tcr亚单位包含tcr细胞内结构域,所述tcr细胞内结构域包含选 自cd3ε、cd3γ或cd3δ的细胞内信号传导结构域的刺激结构域,或 其具有至少一个修饰的氨基酸序列。17.如实施方案1-16中任一项所述的分离的核酸分子,其中所述 tcr亚单位包含细胞内结构域,所述细胞内结构域包含选自4-1bb的 功能性信号传导结构域和/或cd3ζ的功能性信号传导结构域的刺激 结构域,或其具有至少一个修饰的氨基酸序列。18.如实施方案1-17中任一项所述的分离的核酸分子,其中所述 人或人源化抗体结构域包含抗体片段。19.如实施方案1-18中任一项所述的分离的核酸分子,其中所述 人或人源化抗体结构域包含scfv或v
h
结构域。20.如实施方案1-19中任一项所述的分离的核酸分子,其中所述 人或人源化抗体结构域包含sdab或v
hh
结构域。21.如实施方案1-20中任一项所述的分离的核酸分子,其编码(i) 抗间皮素轻链结合结构域氨基酸序列的轻链(lc)cdr1、lc cdr2 和lc cdr3,其分别与本文提供的抗间皮素轻链结合结构域的轻链 (lc)cdr1、lc cdr2和lc cdr3具有70-100%的序列同一性, 和/或(ii)抗间皮素重链结合结构域氨基酸序列的重链(hc)cdr1、 hc cdr2和hc cdr3,其分别与本文提供的抗间皮素重链结合结构 域的重链(hc)cdr1、hc cdr2和hc cdr3具有70-100%的序列 同一性。22.如实施方案1-21中任一项所述的分离的核酸分子,其编码轻 链可变区,其中所述轻链可变区包含相对于本文提供的轻链可变区的 轻链可变区氨基酸序列具有至少一个但不多于30个修饰的氨基酸序 列,或与本文提供的轻链可变区的轻链可变区氨基酸序列具有95-99% 同一性的序列。23.如实施方案1-22中任一项所述的分离的核酸分子,其编码重 链可变区,其中所述重链可变区包含相对于本文提供的重链可变区的 重链可变区氨基酸序列具有至少一个但不多于30个修饰的氨基酸序 列,或与本文提供的重链可变区的重链可变区氨基酸序列具有95-99% 同一性的序列。24.如实施方案1-23中任一项所述的分离的核酸分子,其编码重 链可变区,其中所述重链可变区包含相对于本文提供的重链可变区的 重链可变区氨基酸序列具有至少一个但不多于30个修饰的氨基酸序 列,或与本文提供的v
hh
区的单结构域抗体氨基酸序列具有95-99% 同一性的序列。
25.如实施方案24所述的分离的核酸分子,其中所述v
hh
区的 序列在本文提供的序列中列出。26.如实施方案1-23中任一项所述的分离的核酸分子,其中所述 tfp包含tcr亚单位的细胞外结构域,所述细胞外结构域包括选自 下组的蛋白质的细胞外结构域或其部分:tcrα链、tcrβ链、cd3ε tcr亚单位、cd3γtcr亚单位、cd3δtcr亚单位、其功能片段及 其具有至少一个但不多于20个修饰的氨基酸序列。27.如实施方案1-26中任一项所述的分离的核酸分子,其中所编 码的tfp包含跨膜结构域,所述跨膜结构域包括选自下组的蛋白质的 跨膜结构域:tcrα链、tcrβ链、cd3εtcr亚单位、cd3γtcr亚 单位、cd3δtcr亚单位、其功能片段及其具有至少一个但不多于20 个修饰的氨基酸序列。28.如实施方案1-27中任一项所述的分离的核酸分子,其中所编 码的tfp包含跨膜结构域,所述跨膜结构域包括选自下组的蛋白质的 跨膜结构域:tcrα链、tcrβ链、tcrζ链、cd3εtcr亚单位、cd3γ tcr亚单位、cd3δtcr亚单位、cd45、cd2、cd4、cd5、cd8、 cd9、cd16、cd22、cd33、cd28、cd37、cd64、cd80、cd86、 cd134、cd137、cd154、其功能片段及其具有至少一个但不多于20 个修饰的氨基酸序列。29.如实施方案1-28中任一项所述的分离的核酸分子,其进一步 包含编码共刺激结构域的序列。30.如实施方案29所述的分离的核酸分子,其中所述共刺激结构 域是从选自下组的蛋白质获得的功能性信号传导结构域:dap10、 dap12、cd30、light、ox40、cd2、cd27、cd28、cds、icam
-ꢀ
1、lfa-1(cd11a/cd18)、icos(cd278)和4-1bb(cd137)及其 具有至少一个但不多于20个修饰的氨基酸序列。31.如实施方案1-30中任一项所述的分离的核酸分子,其中所述 至少一个但不多于20个修饰包括介导细胞信号传导的氨基酸修饰或 响应于配体与所述tfp结合而磷酸化的氨基酸修饰。32.如实施方案1-31中任一项所述的分离的核酸分子,其中所述 分离的核酸分子为mrna。33.如实施方案1-32中任一项所述的分离的核酸分子,其中所述 tfp包含tcr亚单位的基于免疫受体酪氨酸的活化基序(itam), 所述itam包括选自下组的蛋白质的itam或其部分:cd3ζtcr亚 单位、cd3εtcr亚单位、cd3γtcr亚单位、cd3δtcr亚单位、 tcrζ链、fcε受体1链、fcε受体2链、fcγ受体1链、fcγ受体2a 链、fcγ受体2b1链、fcγ受体2b2链、fcγ受体3a链、fcγ受体3b 链、fcβ受体1链、tyrobp(dap12)、cd5、cd16a、cd16b、cd22、 cd23、cd32、cd64、cd79a、cd79b、cd89、cd278、cd66d、其 功能片段及其具有至少一个但不多于20个修饰的氨基酸序列。34.如实施方案33所述的分离的核酸分子,其中所述itam取 代cd3γ、cd3δ或cd3ε的itam。35.如实施方案33所述的分离的核酸分子,其中所述itam选 自cd3ζtcr亚单位、cd3εtcr亚单位、cd3γtcr亚单位和cd3δ tcr亚单位,并取代选自cd3ζtcr亚单位、cd3εtcr亚单位、 cd3γtcr亚单位和cd3δtcr亚单位的不同itam。36.如实施方案1-35中任一项所述的分离的核酸分子,其中所述 核酸包含核苷酸类似
物。37.如实施方案36所述的分离的核酸分子,其中所述核苷酸类似 物选自2
’-
o-甲基、2
’-
o-甲氧基乙基(2
’-
o-moe)、2
’-
o-氨基丙基、 2
’-
脱氧、t-脱氧-2
’-
氟代、2
’-
o-氨基丙基(2
’-
o-ap)、2
’-
o-二甲基氨 基乙基(2
’-
o-dmaoe)、2
’-
o-二甲基氨基丙基(2
’-
o-dmap)、t
-ꢀ
o-二甲基氨基乙氧基乙基(2
’-
o-dmaeoe)、2
’-
o-n-甲基乙酰氨基 (2
’-
o-nma)修饰的、锁定核酸(lna)、乙烯核酸(ena)、肽核 酸(pna)、1
’
,5
’-
脱水己糖醇核酸(hna)、吗啉代、甲基膦酸酯核 苷酸、硫醇膦酸酯核苷酸和2
’-
氟代n3-p5
’-
亚磷酰胺。38.如实施方案1-37中任一项所述的分离的核酸分子,其进一步 包含前导序列。39.如实施方案1-38中任一项所述的分离的核酸分子,其中包含 作为由所述核酸编码的抗间皮素结合结构域的抗原结合结构域的所 述人或人源化抗体结构域,或包含所述抗间皮素结合结构域的抗体, 或表达由所述核酸编码的抗间皮素结合结构域的细胞,具有以下亲和 力值:至多约200nm、100nm、75nm、50nm、25nm、20nm、15 nm、14nm、13nm、12nm、11nm、10nm、9nm、8nm、7nm、 6nm、5nm、4nm、3nm、2nm、1nm、0.9nm、0.8nm、0.7nm、 0.6nm、0.5nm、0.4nm、0.3nm、0.2nm、0.1nm、0.09nm、0.08 nm、0.07nm、0.06nm、0.05nm、0.04nm、0.03nm、0.02nm或0.01 nm;和/或至少约100nm、75nm、50nm、25nm、20nm、15nm、 14nm、13nm、12nm、11nm、10nm、9nm、8nm、7nm、6nm、 5nm、4nm、3nm、2nm、1nm、0.9nm、0.8nm、0.7nm、0.6nm、 0.5nm、0.4nm、0.3nm、0.2nm、0.1nm、0.09nm、0.08nm、0.07 nm、0.06nm、0.05nm、0.04nm、0.03nm、0.02nm或0.01nm;和 /或约200nm、100nm、75nm、50nm、25nm、20nm、15nm、14 nm、13nm、12nm、11nm、10nm、9nm、8nm、7nm、6nm、5 nm、4nm、3nm、2nm、1nm、0.9nm、0.8nm、0.7nm、0.6nm、 0.5nm、0.4nm、0.3nm、0.2nm、0.1nm、0.09nm、0.08nm、0.07 nm、0.06nm、0.05nm、0.04nm、0.03nm、0.02nm或0.01nm。40.一种由实施方案1-39中任一项所述的核酸分子编码的分离的 多肽分子。41.一种分离的重组tfp分子,其包含人或人源化抗间皮素结合 结构域、tcr细胞外结构域、跨膜结构域和细胞内结构域。42.一种分离的重组tfp分子,其包含人或人源化抗间皮素结合 结构域、tcr细胞外结构域、跨膜结构域和细胞内信号传导结构域, 其中所述tfp分子能够与内源tcr复合物和/或至少一种内源tcr 多肽在功能上相互作用。43.一种分离的重组tfp分子,其包含人或人源化抗间皮素结合 结构域、tcr细胞外结构域、跨膜结构域和细胞内信号传导结构域, 其中所述tfp分子能够在功能上整合到内源tcr复合物中。44.如实施方案41所述的分离的tfp分子,其包含抗体或抗体 片段,所述抗体或抗体片段包含人或人源化抗间皮素结合结构域、 tcr细胞外结构域、跨膜结构域和细胞内结构域。45.如实施方案41-44中任一项所述的分离的tfp分子,其中所 述抗间皮素结合结构域是scfv或v
h
结构域。46.如实施方案41-45中任一项所述的分离的tfp分子,其中所 述抗间皮素结合结构域包含与本文提供的抗间皮素轻链的氨基酸序 列、其功能片段或其具有至少一个但不多于30个修饰的氨基酸序列 具有95-100%同一性的重链。47.如实施方案41-46中任一项所述的分离的tfp分子,其中所 述抗间皮素结合结构域包含与本文提供的抗间皮素重链的氨基酸序 列、其功能片段或其具有至少一个但不多于
30个修饰的氨基酸序列 具有95-100%同一性的轻链。48.如实施方案41-47中任一项所述的分离的tfp分子,其包含 tcr细胞外结构域,所述tcr细胞外结构域包括选自下组的蛋白质 的细胞外结构域或其部分:tcrα链、tcrβ链、cd3εtcr亚单位、 cd3γtcr亚单位、cd3δtcr亚单位、其功能片段及其具有至少一 个但不多于20个修饰的氨基酸序列。49.如实施方案41-48中任一项所述的分离的tfp分子,其中所 述tcr包含选自下组的蛋白质的细胞外结构域或其部分:t细胞受 体的α或β链、cd3δ、cd3ε或cd3γ。50.如实施方案41-49中任一项所述的分离的tfp分子,其中所 述抗间皮素结合结构域通过连接体序列与所述tcr细胞外结构域连 接。51.如实施方案50所述的分离的tfp分子,其中所述连接体区 包括(g4s)
n
,其中n=1至4。52.如实施方案41-51中任一项所述的分离的tfp分子,其进一 步包含编码共刺激结构域的序列。53.如实施方案41-52中任一项所述的分离的tfp分子,其进一 步包含编码细胞内信号传导结构域的序列。54.如实施方案41-53中任一项所述的分离的tfp分子,其进一 步包含前导序列。55.一种包含编码实施方案41-54中任一项所述的tfp的序列的 核酸。56.如实施方案55所述的核酸,其中所述核酸选自dna和rna。57.如实施方案55或56所述的核酸,其中所述核酸为mrna。58.如实施方案55-57中任一项所述的核酸,其中所述核酸包含 核苷酸类似物。59.如实施方案58所述的核酸,其中所述核苷酸类似物选自2
’-ꢀ
o-甲基、2
’-
o-甲氧基乙基(2
’-
o-moe)、2
’-
o-氨基丙基、2
’-
脱氧、 t-脱氧-2
’-
氟代、2
’-
o-氨基丙基(2
’-
o-ap)、2
’-
o-二甲基氨基乙基(2
’-ꢀ
o-dmaoe)、2
’-
o-二甲基氨基丙基(2
’-
o-dmap)、t-o-二甲基氨 基乙氧基乙基(2
’-
o-dmaeoe)、2
’-
o-n-甲基乙酰氨基(2
’-
o-nma) 修饰的、锁定核酸(lna)、乙烯核酸(ena)、肽核酸(pna)、 1
’
,5
’-
脱水己糖醇核酸(hna)、吗啉代、甲基膦酸酯核苷酸、硫醇膦 酸酯核苷酸和2
’-
氟代n3-p5
’-
亚磷酰胺。60.如实施方案55-59中任一项所述的核酸,其进一步包含启动 子。61.如实施方案55-60中任一项所述的核酸,其中所述核酸是体 外转录的核酸。62.如实施方案55-61中任一项所述的核酸,其中所述核酸进一 步包含编码聚(a)尾的序列。63.如实施方案55-62中任一项所述的核酸,其中所述核酸进一 步包含3
’
utr序列。64.一种包含编码实施方案41-54中任一项所述的tfp的核酸分 子的载体。65.如实施方案64所述的载体,其中所述载体选自dna、rna、 质粒、慢病毒载体、腺病毒载体、劳斯肉瘤病毒(rsv)载体或逆转 录病毒载体。66.如实施方案64或65所述的载体,其进一步包含启动子。67.如实施方案64-66中任一项所述的载体,其中所述载体是体 外转录的载体。68.如实施方案64-67中任一项所述的载体,其中所述载体中的 核酸序列进一步包含聚(a)尾。69.如实施方案64-68中任一项所述的载体,其中所述载体中的 核酸序列进一步包含
3
’
utr。70.一种细胞,其包含实施方案1-39中任一项所述的分离的核酸 分子、实施方案40所述的多肽分子、实施方案41-54中任一项所述 的tfp分子、实施方案55-63中任一项所述的核酸、实施方案64-69 中任一项所述的载体。71.如实施方案70所述的细胞,其中所述细胞是人t细胞。72.如实施方案71所述的细胞,其中所述t细胞是cd8+或cd4+ t细胞。73.如实施方案71所述的细胞,其中所述t细胞是γ-δt细胞。74.如实施方案71所述的细胞,其中所述t细胞是nkt细胞。75.如实施方案70-73中任一项所述的细胞,其进一步包含编码 抑制性分子的核酸,所述抑制性分子包含第一多肽,所述第一多肽包 含抑制性分子的至少一部分,与包含来自细胞内信号传导结构域的正 信号的第二多肽缔合。76.如实施方案75所述的细胞,其中所述抑制性分子包含第一多 肽和第二多肽,所述第一多肽包含pd1的至少一部分,所述第二多肽 包含共刺激结构域和初级信号传导结构域。77.一种包含至少两个tfp分子的人cd8+或cd4+ t细胞,所 述tfp分子包含人或人源化抗间皮素结合结构域、tcr细胞外结构 域、跨膜结构域和细胞内结构域,其中所述tfp分子能够与所述人 cd8+或cd4+ t细胞的表面之中、之处和/或之上的内源tcr复合物 和/或至少一种内源tcr多肽在功能上相互作用。78.一种蛋白质复合物,其包含:i)tfp分子,其包含人或人源化抗间皮素结合结构域、tcr细胞 外结构域、跨膜结构域和细胞内结构域;以及ii)至少一种内源tcr亚单位或内源tcr复合物。79.如实施方案78所述的蛋白质复合物,其中所述tcr包含选 自下组的蛋白质的细胞外结构域或其部分:tcrα链、tcrβ链、cd3ε tcr亚单位、cd3γtcr亚单位和cd3δtcr亚单位。80.如实施方案78或79所述的蛋白质复合物,其中所述抗间皮 素结合结构域通过连接体序列与所述tcr细胞外结构域连接。81.如实施方案80所述的蛋白质复合物,其中所述连接体区包括 (g4s)
n
,其中n=1至4。82.一种蛋白质复合物,其包含:(a)由实施方案1-39中任一项所述的分离的核酸分子编码的tfp, 以及(b)至少一种内源tcr亚单位或内源tcr复合物。83.一种人cd8+或cd4+ t细胞,其中每个实施方案78-82中任 一项所述的蛋白质复合物包含至少两个不同的tfp蛋白。84.一种人cd8+或cd4+ t细胞,其包含至少两个不同的由实施 方案1-39中任一项所述的分离的核酸分子编码的tfp分子。85.一种人cd8+或cd4+ t细胞的群体,其中所述群体的t细 胞单独或共同地包含至少两个tfp分子,所述tfp分子包含人或人 源化抗间皮素结合结构域、tcr细胞外结构域、跨膜结构域和细胞内 结构域,其中所述tfp分子能够与所述人cd8+或cd4+ t细胞的表 面之中、之处和/或之上的内源tcr复合物和/或至少一种内源tcr 多肽在功能上相互作用。
86.一种人cd8+或cd4+ t细胞的群体,其中所述群体的t细 胞单独或共同地包含至少两个由实施方案1-39中任一项所述的分离 的核酸分子编码的tfp分子。87.一种制备细胞的方法,该方法包括用实施方案1-39中任一项 所述的分离的核酸分子、实施方案55-63中任一项所述的核酸或实施 方案64-69中任一项所述的载体转导t细胞。88.一种产生rna工程化细胞群体的方法,该方法包括将体外 转录的rna或合成rna引入细胞中,其中所述rna包含编码实施 方案41-54中任一项所述的tfp分子的核酸。89.一种在哺乳动物中提供抗肿瘤免疫的方法,该方法包括向所 述哺乳动物施用有效量的实施方案1-39中任一项所述的分离的核酸 分子、实施方案40所述的多肽分子、表达实施方案40所述的多肽分 子的细胞、实施方案41-54中任一项所述的tfp分子、实施方案55
-ꢀ
63中任一项所述的核酸、实施方案64-69中任一项所述的载体或者实 施方案70-77和83-87中任一项所述的细胞。90.如实施方案89所述的方法,其中所述细胞是自体t细胞。91.如实施方案89所述的方法,其中所述细胞是同种异体t细 胞。92.如实施方案89-91中任一项所述的方法,其中所述哺乳动物 是人。93.一种治疗患有与间皮素表达相关的疾病的哺乳动物的方法, 该方法包括向所述哺乳动物施用有效量的实施方案1-39中任一项所 述的分离的核酸分子、实施方案40所述的多肽分子、表达实施方案 40所述的多肽分子的细胞、实施方案41-54中任一项所述的tfp分 子、实施方案55-63中任一项所述的核酸、实施方案64-69中任一项 所述的载体或者实施方案70-77和83-86中任一项所述的细胞。94.如实施方案93所述的方法,其中所述与间皮素表达相关的疾 病选自增生性疾病、癌症、恶性肿瘤和与间皮素表达相关的非癌症相 关适应症。95.如实施方案93所述的方法,其中所述疾病是选自间皮瘤、肾 细胞癌、胃癌、乳腺癌、肺癌、卵巢癌、前列腺癌、结肠癌、宫颈癌、 脑癌、肝癌、胰腺癌、甲状腺癌、膀胱癌、输尿管癌、肾癌、子宫内 膜癌、食管癌、胃癌、胸腺癌、胆管癌和胃癌的癌症。96.如实施方案93所述的方法,其中所述疾病是选自下组的癌 症:间皮瘤、乳头状浆液性卵巢腺癌、透明细胞卵巢癌、混合型米勒 氏卵巢癌、子宫内膜粘液性卵巢癌、胰腺腺癌、胰腺导管腺癌、子宫 浆液性癌、肺腺癌、肝外胆管癌、胃腺癌、食管腺癌、结直肠腺癌、 乳腺腺癌、与间皮素表达相关的疾病及其组合。97.如实施方案93所述的方法,其中所述表达tfp分子的细胞 与提高表达tfp分子的细胞的功效的药剂联合施用。98.如实施方案93-97中任一项所述的方法,其中与施用有效量 的表达抗间皮素嵌合抗原受体(car)的t细胞的哺乳动物相比,所 述哺乳动物中释放的细胞因子较少。99.如实施方案93-98中任一项所述的方法,其中所述表达tfp 分子的细胞与改善与施用表达tfp分子的细胞相关的一种或多种副 作用的药剂联合施用。100.如实施方案93-99中任一项所述的方法,其中所述表达tfp 分子的细胞与治疗与间皮素相关的疾病的药剂联合施用。101.用作药物的实施方案1-39中任一项所述的分离的核酸分子、 实施方案40所述的分离的多肽分子、表达实施方案40所述的多肽分 子的细胞、实施方案41-54中任一项所述
的分离的tfp、实施方案55
-ꢀ
63中任一项所述的核酸、实施方案64-69中任一项所述的载体、实施 方案78-82中任一项所述的复合物或者实施方案70-77和83-86中任 一项所述的细胞。102.一种治疗患有与间皮素表达相关的疾病的哺乳动物的方法, 该方法包括向所述哺乳动物施用有效量的实施方案1-39中任一项所 述的分离的核酸分子、实施方案40所述的多肽分子、表达实施方案 40所述的多肽分子的细胞、实施方案41-54中任一项所述的tfp分 子、实施方案55-63中任一项所述的核酸、实施方案64-69中任一项 所述的载体或者实施方案70-77和83-86中任一项所述的细胞,其中 与施用有效量的表达抗间皮素嵌合抗原受体(car)的t细胞的哺乳 动物相比,所述哺乳动物中释放的细胞因子较少。实施例
[0312]
通过参考以下实验实施例进一步详细描述本发明。提供这些实施 例仅仅是为了说明的目的,除非另有说明,否则并非意在限制。因此, 本发明绝不应被解释为限于以下实施例,而应被解释为涵盖由于本文所 提供的教导而变得显而易见的任何和所有变化。在没有进一步描述的情 况下,相信本领域普通技术人员可使用前面的描述和以下说明性实施 例制备和利用本发明的化合物并实践所要求保护的方法。以下工作实 施例具体指出了本发明的各个方面,并且不应被解释为以任何方式限 制本公开内容的其余部分。实施例1:tfp构建体
[0313]
通过将经由编码短连接体(sl): aaaggggsggggsggggsle(seq id no:2)或长连接体(ll): aaaievmypppylggggsggggsggggsle(seq id no:3)的dna 序列与cd3或tcr dna片段连接的抗间皮素scfv dna片段克隆到 p510载体(system biosciences(sbi))中的xbai和ecor1位点, 来工程化抗间皮素tfp构建体。
[0314]
产生的抗间皮素tfp构建体是p510_抗间皮素_ll_tcrα(抗 间皮素scfv-长连接体-人全长t细胞受体α链)、p510_抗间皮素 _ll_tcrαc(抗间皮素scfv-长连接体-人t细胞受体α恒定域链)、 p510_抗间皮素_ll_tcrβ(抗间皮素scfv-长连接体-人全长t细胞受 体β链)、p510_抗间皮素_ll_tcrβc(抗间皮素scfv-长连接体-人 t细胞受体β恒定域链)、p510_抗间皮素_ll_cd3γ(抗间皮素scfv
-ꢀ
长连接体-人cd3γ链)、p510_抗间皮素_ll_cd3δ(抗间皮素scfv
-ꢀ
长连接体-人cd3δ链)、p510_抗间皮素_ll_cd3ε(抗间皮素scfv
-ꢀ
长连接体-人cd3ε链)、p510_抗间皮素_sl_tcrβ(抗间皮素scfv
-ꢀ
短连接体-人全长t细胞受体β链)、p510_抗间皮素_sl_cd3γ(抗 间皮素scfv-短连接体-人cd3γ链)、p510_抗间皮素_sl_cd3δ(抗 间皮素scfv-短连接体-人cd3δ链)、p510_抗间皮素_sl_cd3ε(抗 间皮素scfv-短连接体-人cd3ε链)。
[0315]
通过将编码抗间皮素、部分cd28细胞外结构域、cd28跨膜结 构域、cd28细胞内结构域和cd3ζ的合成dna克隆到p510载体中 的xbai和ecor1位点来产生抗间皮素car构建体p510_抗间皮素 _28ζ。实施例2:抗体序列抗体序列的产生
[0316]
人间皮素多肽规范序列是uniprot登录号q13421(或q13421
-ꢀ
1)。提供了能够特异性结合人间皮素多肽及其片段或结构域的抗体 多肽。可使用多种技术产生抗间皮素抗体(参见例如,nicholson等人, 1997)。在使用鼠抗间皮素抗体作为起始材料的情况下,对于
临床环 境需要鼠抗间皮素抗体的人源化,在临床环境中,小鼠特异性残基可 在接受t细胞受体(tcr)融合蛋白(tfp)治疗(即采用tfp间皮 素构建体转导的t细胞的治疗)的受试者中诱导人抗小鼠抗原 (hama)应答。通过将来自鼠抗间皮素抗体的cdr区移植到适当 的人种系受体构架上来完成人源化,任选地包括对cdr和/或构架区 的其他修饰。如本文提供的,抗体和抗体片段残基编号遵循kabat编 号(kabat e.a.等人,1991;chothia等人,1987)。scfv的产生
[0317]
使用人或人源化抗间皮素igg产生tfp构建体的scfv序列。 获得编码人或人源化v
l
和v
h
结构域的dna序列,并且任选地针对 在来自智人(homo sapiens)的细胞中表达而优化构建体的密码子。 v
l
和v
h
结构域在scfv中出现的顺序是变化的(即,v
l-v
h
或v
h-v
l
方向),并且“g4s”或“g4s”亚单位的三个拷贝(g4s)3连接可变域以产 生scfv域。抗间皮素scfv质粒构建体可具有可选的flag、his或其 他亲和标签,并且被电穿孔到hek293或其他合适的人或哺乳动物细 胞系中,并进行纯化。验证试验包括通过facs的结合分析、使用 proteon的动力学分析和表达间皮素的细胞的染色。
[0318]
示例性的抗间皮素v
l
和v
h
结构域、cdr和编码它们的核苷酸 序列可以是美国专利号9,272,002、8,206,710、9,023,351、7,081,518、8,911,732、9,115,197和9,416,190以及美国专利公开号20090047211 中描述的那些。其他示例性抗间皮素v
l
和v
h
结构域、cdr和编码 它们的核苷酸序列分别可属于以下单克隆抗体:大鼠抗间皮素抗体 420411、大鼠抗间皮素抗体420404、小鼠抗间皮素抗体mn-1、小鼠 抗间皮素抗体mb-g10、小鼠抗间皮素抗体abin233753、兔抗间皮 素抗体fqs3796(3)、兔抗间皮素抗体tq85、小鼠抗间皮素抗体 ta307799、大鼠抗间皮素抗体295d、大鼠抗间皮素抗体b35、小鼠 抗间皮素抗体5g157、小鼠抗间皮素抗体129588、兔抗间皮素抗体 11c187、小鼠抗间皮素抗体5b2、兔抗间皮素抗体sp74、兔抗间皮素 抗体d4x7m、小鼠抗间皮素抗体c-2、小鼠抗间皮素抗体c-3、小鼠 抗间皮素抗体g-1、小鼠抗间皮素抗体g-4、小鼠抗间皮素抗体k1、 小鼠抗间皮素抗体b-3、小鼠抗间皮素抗体200-301-a87、小鼠抗间皮 素抗体200-301-a88、兔抗间皮素抗体epr2685(2)、兔抗间皮素抗体 epr4509或兔抗间皮素抗体ppi-2e(ihc)。
[0319]
在一些实施方案中,使用单结构域(v
hh
)结合物,例如seq idno:53-55中所示的结合物(分别为sd1、sd4和sd6)。tcr亚单位的来源
[0320]
人t细胞受体(tcr)复合物的亚单位均含有细胞外结构域、 跨膜结构域和细胞内结构域。人tcr复合物含有cd3-ε多肽、cd3
-ꢀ
γ多肽、cd3-δ多肽、cd3-ζ多肽、tcrα链多肽和tcrβ链多肽。人 cd3-ε多肽规范序列为uniprot登录号p07766。人cd3-γ多肽规范序 列为uniprot登录号p09693。人cd3-δ多肽规范序列为uniprot登录 号p043234。人cd3-ζ多肽规范序列为uniprot登录号p20963。人 tcrα链规范序列为uniprot登录号q6isu1。人tcrβ链c区规范序 列为uniprot登录号p01850,人tcrβ链v区序列为p04435。
[0321]
人cd3-ε多肽规范序列为: mqsgthwrvlglcllsvgvwgqdgneemggitqtpykvsisgtt viltcpqypgseilwqhndkniggdeddknigsdedhlslkefsele qsgyyvcyprgskpedanfylylrarvcencmemdvmsvativi vdicitggllllvyywsknrkakakpvtrgagaggrqrgqnker pppvpnpdyepirkgqrdlysglnqrri(seq id no:4)。
[0322]
人cd3-γ多肽规范序列为: meqgkglavlilaiillqgtlaqsikgnhlvkvydyqedgsvll
tc daeaknitwfkdgkmigfltedkkkwnlgsnakdprgmyqckg sqnkskplqvyyrmcqncielnaatisgflfaeivsifvlavgvyfi agqdgvrqsrasdkqtllpndqlyqplkdreddqyshlqgnqlr rn(seq id no:5)。
[0323]
人cd3-δ多肽规范序列为: mehstflsglvlatllsqvspfkipieeledrvfvncntsitwvegt vgtllsditrldlgkrildprgiyrcngtdiykdkestvqvhyrmc qscveldpatvagiivtdviatlllalgvfcfaghetgrlsgaadt qallrndqvyqplrdrddaqyshlggnwarnks(seq id no:6)。
[0324]
人cd3-ζ多肽规范序列为: mkwkalftaailqaqlpiteaqsfglldpklcylldgilfiygvilt alflrvkfsrsadapayqqgqnqlynelnlgrreeydvldkrrg rdpemggkpqrrknpqeglynelqkdkmaeayseigmkgerrrg kghdglyqglstatkdtydalhmqalppr(seq id no:7)。
[0325]
人tcrα链规范序列为: magtwlllllalgcpalptgvggtpfpslappimllvdgkqqmvv vclvldvappgldspiwfsagngsaldaftygpspatdgtwtnla hlslpseelasweplvchtgpgaeghsrstqpmhlsgeastartcp qeplrgtpggalwlgvlrlllfklllfdllltcsclcdpagplpsp atttrlralgshrlhpatetggreatssprpqprdrrwgdtppgr kpgspvwgegsylssyptcpaqawcsrsalrapssslgaffagdlp pplqagaa(seq id no:57)。
[0326]
人tcrα链c区规范序列为: pniqnpdpavyqlrdskssdksvclftdfdsqtnvsqskdsdvyit dktvldmrsmdfksnsavawsnksdfacanafnnsiipedtffpsp esscdvklveksfetdtnlnfqnlsvigfrilllkvagfnllmtlrl wss(seq id no:9)。
[0327]
人tcrα链v区ctl-l17规范序列为: mamllgasvlilwlqpdwvnsqqknddqqvkqnspslsvqegri silncdytnsmfdyflwykkypaegptflisissikdknedgrftvf lnksakhlslhivpsqpgdsavyfcaakgagtaskltfgtgtrlq vtl(seq id no:10)。
[0328]
人tcrβ链c区规范序列为: edlnkvfppevavfepseaeishtqkatlvclatgffpdhvelsww vngkevhsgvstdpqplkeqpalndsryclssrlrvsatfwqnpr nhfrcqvqfyglsendewtqdrakpvtqivsaeawgradcgfts vsyqqgvlsatilyeillgkatlyavlvsalvlmamvkrkdf (seq id no:11)。
[0329]
人tcrβ链v区ctl-l17规范序列为: mgtsllcwmalcllgadhadtgvsqnprhnitkrgqnvtfrcdp isehnrlywyrqtlgqgpefltyfqneaqleksrllsdrfsaerpk gsfstleiqrteqgdsamylcasslaglnqpqhfgdgtrlsil(seq id no:12)。
[0330]
人tcrβ链v区yt35规范序列为: mdswtfccvslcilvakhtdagviqsprhevtemgqevtlrckpis ghnslfwyrqtmmrglelliyfnnnvpiddsgmpedrfsakmpn asfstlkiqpseprdsavyfcassfstcsanygytfgsgtrltvv (seq id no:13)。从tcr结构域和scfv产生tfp
[0331]
使用连接体序列如g4s、(g4s)2、(g4s)3或(g4s)4将间皮素scfv 以重组方式连接至cd3-ε或其他tcr亚单位(参见图2)。使用各种 连接体和scfv配置。作为全长多肽或仅其恒定域,使用tcrα和tcrβ 链产生tfp。允许用tcrα和tcrβ链的任何可变序列制备tfp。tfp表达载体
[0332]
提供了表达载体,其包含:启动子(巨细胞病毒(cmv)增强 子-启动子)、实现分泌的信号传导序列、聚腺苷酸化信号和转录终止 子(牛生长激素(bgh)基因)、允许附加型复制和在原核生物中复 制的元件(例如,sv40起点和cole1或本领域已知的其他元件)以 及允许选择的元件(氨苄青霉素抗性基因和零霉素(zeocin)标记)。
[0333]
优选地,将编码tfp的核酸构建体克隆到慢病毒表达载体中, 并基于tfp.间皮素
转导的t细胞(“间皮素.tfp”或“间皮素.tfp t细 胞”或“tfp.间皮素”或“tfp.间皮素t细胞”)响应于间皮素+靶细胞的 效应t细胞应答的数量和质量来验证表达。效应t细胞应答包括但不 限于细胞扩充、增殖、倍增、细胞因子产生和靶细胞溶解或细胞溶解 活性(即脱粒)。
[0334]
使用tfp.间皮素慢病毒转移载体产生包装在vsv-g假型化慢 病毒颗粒中的基因组物质。将慢病毒转移载体dna与vsv-g、gag/pol 和rev这三种包装组分以及试剂混合,以将它们一起 转染到hek-293(胚肾,crl-1573
tm
)细胞中。在24和48 小时后,收集培养基,过滤,并通过超速离心浓缩。所得病毒制剂储 存在-80℃下。通过在sup-t1(t细胞淋巴母细胞淋巴瘤, crl-1942
tm
)细胞上滴定来确定转导单元的数目。通过用例如抗cd3 抗cd28珠子激活新鲜的幼稚t细胞达24小时,然后添加适当数目 的转导单元以获得所需百分比的转导t细胞,来产生重定向的tfp. 间皮素t细胞。允许这些经修饰的t细胞扩充,直至它们静止并且大 小减小,此时将它们冷冻保存以供后续分析。使用coulter multisizer
tm iii测量细胞数目和大小。在冷冻保存之前,通过流式细胞术分析来确 定转导的细胞(在细胞表面上表达tfp.间皮素)的百分比和该表达的 相对荧光强度。从直方图中,通过比较转导的百分比与它们的相对荧 光强度来检查tfp的相对表达水平。
[0335]
在一些实施方案中,通过用多种病毒载体转导t细胞来引入多 种tfp。评估人源化tfp重定向t细胞的细胞溶解活性、增殖能力和细胞因 子分泌
[0336]
使用本领域已知的试验来确定tfp.间皮素t细胞产生细胞表面 表达的tfp以及杀伤靶肿瘤细胞、增殖和分泌细胞因子的功能能力。
[0337]
用人白细胞介素2(il-2)处理人外周血单个核细胞(pbmc, 例如来自进行单采血液成分术的正常供体的血液,通过对t细胞、 cd4+和cd8+淋巴细胞进行负选择获得其幼稚t细胞),然后用抗 cd3x抗cd28珠激活,例如在37℃、5%co2下在10%rpmi中,然 后用编码tfp的慢病毒载体转导。使用流式细胞术试验确认细胞表面 存在tfp,诸如通过抗flag抗体或抗鼠可变域抗体。使用elisa或 其他测定来测量细胞因子(例如,ifn-γ)产生。实施例3:人all小鼠模型中的人tfp t细胞功效
[0338]
原代人all细胞可在免疫受损的小鼠(例如,nsg或nod) 中生长,而不必在体外培养。同样,培养的人all细胞系可在这样的 小鼠中诱导白血病。携带all的小鼠可用于测试人tfp.间皮素t细 胞的功效,例如,在模型hallx5447中。该模型中的读出是在不存 在和存在静脉内(i.v.)施用人tfp.间皮素t细胞的情况下静脉内输 注all细胞后小鼠的存活。实施例4:多重tfp多肽的证明,以及多重人源化tfp重定向t细胞的用途
[0339]
本文提供的tfp多肽能够在功能上与内源tcr亚单位多肽缔 合以形成功能性tcr复合物。这里,使用慢病毒载体中的多个tfp 转导t细胞,以便产生功能性多重重组tcr复合物。例如,提供了 t细胞,其含有i)第一tfp,其具有来自cd3-δ多肽的细胞外结构 域、跨膜结构域和细胞内结构域,以及间皮素特异性scfv抗体片段, 和ii)第二tfp,其具有来自cd3-γ多肽的细胞外结构域、跨膜结构 域和细胞内结构域,以及间皮素特异性scfv抗体片段。第一tfp和 第二tfp能够彼此相互作用并与内源tcr亚单位多肽相互作用,从 而形成功能性tcr复合物。
[0340]
如以上实施例2和实施例3中所提供的,可在液体瘤和实体瘤 中证明这些多重人
源化tfp.间皮素t细胞的用途。实施例5:用tfp转导的t细胞的制备慢病毒产生
[0341]
编码合适的构建体的慢病毒如下制备。将5x106个hek-293ft 细胞接种到100mm培养皿中,并使其达到70-90%汇合度过夜。将 2.5μg指定的dna质粒和20μl慢病毒包装混合物(alstem,目录 号vp100)在0.5ml不含血清的dmem或i培养基中 稀释并轻柔混合。在单独的管中,将30μl转染试剂 (alstem,目录号nf100)在0.5ml不含血清的dmem或不含血清的dmem或i培养基中稀释并轻柔混合。然后将nanofect/dmem和 dna/dmem溶液混合在一起并涡旋10-15秒,然后在室温下温育 dmem-质粒-nanofect混合物15分钟。将来自前一步骤的完全转染 复合物逐滴添加至细胞板并摇动以将转染复合物均匀分散在板中。然 后将板在潮湿的5%co2孵箱中在37℃下温育过夜。第二天,用10 ml新鲜培养基替代上清液并补充20μl viralboost(500x,alstem, 目录号vb100)。然后将板在37℃下再温育24小时。随后将含有慢 病毒的上清液收集到50ml无菌加盖的锥形离心管中并置于冰上。在 4℃下以3000rpm离心15分钟后,用低蛋白质结合的0.45μm无菌 过滤器过滤澄清的上清液,随后通过在4℃下以25,000rpm超速离心 (beckmann,l8-70m)1.5小时分离病毒。取出沉淀物并重新悬浮于 dmem培养基中,使用lenti-x qrt-pcr滴定试剂盒(clontech;目 录号631235)通过定量rt-pcr建立慢病毒浓度/滴度。通过用dna 酶i处理除去任何残留的质粒dna。将病毒原液制备物立即用于感染 或等分并储存在-80℃下以供将来使用。pbmc分离
[0342]
由全血或血沉棕黄层制备外周血单个核细胞(pbmc)。将全血 收集在10ml肝素真空采血管中并立即处理或在4℃下储存过夜。将 约10ml的抗凝全血与无菌磷酸盐缓冲盐水(pbs)缓冲液在50ml 锥形离心管(pbs,ph 7.4,不含ca
2+
/mg
2+
)中混合,总体积为20ml。 然后将20ml该血液/pbs混合物轻轻覆盖在15ml plus(ge healthcare,17-1440-03)的表面上,然后在室温下以400 g离心30-40分钟,不施加减速。
[0343]
血沉棕黄层购自research blood components(boston,ma)。 通过添加15ml(ge health care),并以1000g离心 1分钟来准备管(greiner bio-one)。将血沉棕黄层于pbs (ph 7.4,不含ca
2+
或mg
2+
)中1:3稀释。将稀释的血沉棕黄层转移 至leucosep管并以1000g离心15分钟,不施加减速。小心地去除在 稀释的血浆/ficoll界面处所见到的含有pbmc的细胞层以使ficoll的 污染最小化。然后通过在室温下以200g离心10分钟用40ml pbs 洗涤pbmc三次,来去除残留的ficoll、血小板和血浆蛋白质。随后 用血细胞计数器对细胞进行计数。洗涤的pbmc用car-t培养基 (aim(bsa)(life technologies),其含有5%ab 血清和1.25μg/ml两性霉素b(gemini bioproducts,woodland,ca)、 100u/ml青霉素和100μg/ml链霉素)洗涤一次。或者,将洗涤的 pbmc转移到隔离小瓶中并在-80℃下冷冻24小时,然后储存在液氮 中以供后续使用。t细胞活化
[0344]
在病毒转导之前,用抗人cd28和cd3抗体偶联的磁珠刺激由 全血或血沉棕黄层制备的pbmc达24小时。将新鲜分离的pbmc在 不含huil-2的car-t培养基(aim v-albumax(bsa)(lifetechnologies),其含有5%ab血清和1.25μg/ml两性霉素b(geminibioproducts)、100u/ml青霉素和100μg/ml链霉素)中洗涤一次, 然后以1x106个细胞/ml的终浓度重新悬浮于含有300iu/ml人il-2 (来自1000x原液;invitrogen)的car-t培养基中。如果pbmc先 前已被冷冻,则将它们解冻并以1x107个细胞/ml重新悬浮于存在10% fbs、100u/ml青霉素和100μg/ml链霉素的9ml预热(37℃) cdmem培养基(life technologies)中,浓度为1x106个细胞/ml,然 后在car-t培养基中洗涤一次,以1x106个细胞/ml重新悬浮于car
-ꢀ
t培养基中,并如上所述添加il-2。
[0345]
在活化之前,用1ml无菌1x pbs(ph 7.4)将抗人cd28和 cd3抗体偶联的磁珠(可从例如invitrogen,life technologies获得) 洗涤三次,使用磁架从溶液中分离珠子,然后重新悬浮于含有300 iu/ml人il-2的car-t培养基中,至终浓度为4x107个珠子/ml。随 后通过将25μl(1x106个珠子)珠子转移至1ml pbmc,从而以1:1 的珠子与细胞之比混合pbmc和珠子。然后将所需数目的等分试样分 配到12孔低附着或未处理的细胞培养板的单个孔中,并在37℃、5% co2下温育24小时,然后进行病毒转导。t细胞转导/转染和扩充
[0346]
在pbmc活化后,将细胞在37℃、5%co2下温育48小时。在 冰上解冻慢病毒,并将5x106个慢病毒与2μl transplus
tm
(alstem) /ml培养基(最终稀释度为1:500)一起添加到1x106个细胞的每个孔 中。将细胞再温育24小时,然后重复添加病毒。或者,在冰上解冻慢 病毒,并在5μg/ml聚凝胺(polybrene)(sigma)的存在下以5或50 moi添加相应的病毒。将细胞在室温下以100g旋转接种100分钟。 然后使细胞在300iu/ml人il-2的持续存在下生长6-14天(总温育 时间取决于所需的car-t细胞的最终数目)。每2-3天分析细胞浓 度,此时添加培养基以使细胞悬浮液维持在1x106个细胞/ml。
[0347]
在一些情况下,用体外转录的(ivt)mrna对活化的pbmc 进行电穿孔。在一个实施方案中,在300iu/ml重组人il-2(r&dsystems)的存在下,用dynabeads(thermofisher)以1:1的比例将人 pbmc刺激3天(可使用其他刺激试剂,如来自milyeni pharmaceuticals 的transact t细胞试剂)。在电穿孔之前去除珠子。洗涤细胞并以 2.5x107个细胞/ml的浓度重新悬浮于opti-mem培养基 (thermofisher)中。将200μl细胞悬浮液(5x106个细胞)转移至2 mm间隙electroporation cuvettes plus
tm
(harvard apparatus btx)并 在冰上预冷却。向细胞悬浮液中添加10μg ivt tfp mrna。然后使 用ecm830 electro square wave porator(harvard apparatus btx)在 200v下将mrna/细胞混合物电穿孔20毫秒。在电穿孔后立即将细 胞转移至新鲜细胞培养基(aim v albumax(bsa)无血清培养基+ 5%人ab血清+300iu/ml il-2)中并在37℃下温育。通过细胞染色验证tfp表达
[0348]
在慢病毒转导或mrna电穿孔后,通过流式细胞术确认抗间皮 素tfp的表达,其中使用抗小鼠fab抗体检测鼠抗间皮素scfv。将t 细胞在3ml染色缓冲液(pbs,4%bsa)中洗涤三次,并以每孔1x106个细胞重新悬浮于pbs中。对于死细胞排除,将细胞与 fixable aqua dead cell stain(invitrogen)在冰上一起温育30分钟。 用pbs洗涤细胞两次,并重新悬浮于50μl染色缓冲液中。为了阻断 fc受体,将1μl 1:
100稀释的正常山羊igg(bd bioscience)添加至 每个管中,并在冰中温育10分钟。向每个管添加1.0ml facs缓冲 液,充分混合,通过以300g离心5分钟使细胞沉淀。通过 r-藻红蛋白标记的人msln igg1 fc或人igg1同种型对照检测scfvtfp的表面表达。将1μg抗体添加至相应的样品中并在冰上温育30 分钟。然后将细胞洗涤两次,并使用来自bd bioscience的抗cd3 apc (克隆ucht1)、抗cd4-pacific blue(克隆rpa-t4)、抗-cd8 apccy7 (克隆sk1)对表面标志物进行染色。使用lsrfortessatm x20(bdbiosciences)进行流式细胞术,并使用facs diva软件获取数据,并 用flowjo(treestar,inc.ashland,or)进行分析。
[0349]
示例性结果显示在图5a中,其示出了对cd8(抗cd8 apccy7, y轴)和间皮素(“msln”)(r-藻红蛋白标记的hmsln igg, x轴)染色的活化pbmc细胞的表面表达分析。从左到右显示了未转 导的或用抗msln-cd3ε、抗msln-cd28ζ和抗msln-41bbζ构建 体转导的细胞。cd8+、msln+细胞的比例示于每幅图的右上角。
[0350]
图5b示出了对msln和gfp染色的活化pbmc细胞的类似 结果,该细胞用包含内部单结构域(“sd”)间皮素结合物的tfp构建 体转导。第一行显示(从左到右)未转导的细胞和用阳性对照抗msln
-ꢀ
cd3εtfp(“ss1”)转导的细胞。第2-4行分别显示用gfp标记的(从 左到右)cd3εtfp、cd3γtfp、tcrβtfp和cd28ζcar构建体转 导的细胞中的抗msln结合物sd1、sd4和sd6。gfp+、msln+细 胞的比例示于每幅图的右上角。实施例6:通过流式细胞术进行的细胞毒性测定
[0351]
用荧光染料羧基荧光素二乙酸琥珀酰亚胺酯(cfse)标记对间 皮素呈阳性或阴性的靶细胞。将这些靶细胞与效应t细胞混合,这些 效应t细胞是未转导的、用对照car-t构建体转导的或用tfp转导 的。在指定的温育期后,通过流式细胞术对每种效应细胞/靶细胞培养 物确定死靶细胞与cfse标记的活靶细胞和阴性对照靶细胞的百分比。 相对于仅含有靶细胞的孔计算每种t细胞阳性靶细胞培养物中靶细 胞的存活百分比。
[0352]
通过使用流式细胞术比较在效应细胞和靶细胞共温育后,在没 有或具有效应t细胞的情况下靶细胞中存活的靶细胞的数目,来测量 效应t细胞的细胞毒活性。在使用间皮素tfp或car-t细胞的实验 中,靶细胞是间皮素阳性细胞,而用作阴性对照的细胞是间皮素阴性 细胞。
[0353]
将靶细胞洗涤一次,并以1x106个细胞/ml重新悬浮于pbs中。 将荧光染料羧基荧光素二乙酸琥珀酰亚胺酯(cfse)(thermofisher) 以0.03μm的浓度添加至细胞悬浮液中,并将细胞在室温下温育20 分钟。通过以5倍于反应体积的体积向细胞悬浮液中添加完全细胞培 养基(rpmi-1640+10%hi-fbs)来停止标记反应,并将细胞在室温 下再温育2分钟。通过离心沉淀细胞,并将其以2x105个细胞/ml重 新悬浮于细胞毒性培养基(不含酚红的rpmi1640(invitrogen)加5% ab血清(gemini bioproducts))中。将50微升cfse标记的靶细胞 悬浮液(相当于10,000个细胞)添加至96孔u形底板(corning)的 每个孔中。
[0354]
将用抗间皮素tfp构建体转导的效应t细胞与作为阴性对照的 未转导的t细胞一起洗涤,并以2x106个细胞/ml或1x106个细胞/ml 悬浮在细胞毒性培养基中。将50μl效应t细胞悬浮液(相当于 100,000或50,000个细胞)添加至平板接种的靶细胞,以在100μl的 总体积内分别达到10比1或5比1的效应物与靶标之比。然后将培 养物混合,离心,并在37℃和5%co2下温育4小时。在该温育后, 立即按照制造商的建议将7aad(7-氨基放线菌素d)
(biolegend) 添加至培养的细胞,并用bd lsrfortessa
tm
x-20(bd biosciences) 进行流式细胞术。使用软件(treestar,inc.)进行流式细胞术 数据的分析。
[0355]
通过用具有效应t细胞和靶细胞的样品中的活靶细胞 (cfse+7-aad-)数目除以仅具有靶细胞的样品中的活(cfse+7
-ꢀ
aad-)细胞的数目来计算靶细胞的存活百分比。效应细胞的细胞毒性 计算为靶细胞杀伤百分比=100%-细胞存活百分比。
[0356]
当与未转导或用非间皮素特异性car对照转导的t细胞比较 时,用抗msln 28ζcar构建体转导的t细胞可证明对表达间皮素 的细胞具有细胞毒性。然而,用抗间皮素-cd3ε转导的t细胞可比抗 间皮素car对照诱导更有效的针对靶标的细胞毒性。抗间皮素-cd3γ tfp也可介导强效细胞毒性,其大于抗间皮素car在效应物:靶标比 为5:1-10:1时观察到的细胞毒性。用抗间皮素tcrα和抗间皮素tcrβ tfp可观察到一定的细胞毒性。用替代的铰链区构建的抗间皮素tfp 可获得类似的结果。再者,相比于抗间皮素car转导的t细胞,抗 间皮素-cd3ε或抗间皮素-cd3γtfp转导的t细胞对表达间皮素的靶 细胞的细胞毒性可能较大。
[0357]
用编码间皮素特异性tfp的mrna电穿孔的t细胞也可以表 现出对表达间皮素的细胞强效的细胞毒性。虽然对照或抗间皮素tfp 构建体未观察到间皮素阴性细胞的显著杀伤,但用抗间皮素-cd3εsl 或抗间皮素-cd3γsl tfp转导的t细胞可观察到表达间皮素的细胞 的间皮素特异性杀伤。实施例8:通过实时细胞毒性测定确定细胞毒性
[0358]
在实时细胞毒性测定(rtca)形式中,抗间皮素tfp还可表 现出优于抗间皮素car的细胞毒性。rtca测定实时测量专用96孔 板的每个孔中贴壁的靶细胞单层的电阻抗,并将最终读出呈现为被称 为细胞指数的值。细胞指数的变化表明由于共培养的t细胞效应物杀 伤靶细胞而破坏靶细胞单层。因此,效应t细胞的细胞毒性可被评价 为具有靶细胞和效应t细胞的孔的细胞指数与仅具有靶细胞的孔的 细胞指数相比的变化。
[0359]
在dmem、10%fbs、1%抗生素-抗霉菌剂(life technologies) 中培养贴壁靶细胞。为了制备rtca,将50μl例如dmem培养基 添加至e板(acea biosciences,inc,目录号:jl-10-156010-1a)的 适当孔中。然后将板置于rtca mp仪器(acea biosciences,inc.) 中,并如制造商手册中所述将适当的板布局和测定计划输入rtca 2.0 软件。每15分钟进行基线测量,共100次测量。然后将100μl体积 的1x104个靶细胞添加至每个测定孔,使细胞沉降15分钟。将板返回 读取器并继续读取。
[0360]
第二天,将效应t细胞洗涤并重新悬浮于细胞毒性培养基(不 含酚红的rpmi1640(invitrogen)加5%ab血清(gemini bioproducts; 100-318))中。然后从仪器中取出板,将悬浮在细胞毒性培养基(不 含酚红的rpmi1640+5%ab血清)中的效应t细胞以100,000个细 胞或50,000个细胞添加至每个孔中,以分别达到10比1或5比1的 效应物与靶标之比。然后将板放回仪器。每2分钟进行测量,共100 次测量,然后每15分钟进行测量,共1,000次测量。
[0361]
在rtca测定中,用抗间皮素28ζcar转导的t细胞可观察到 间皮素转导的细胞的杀伤,如根据相对于单独的细胞或与用对照car 构建体转导的t细胞共温育的细胞,添加效应细胞后细胞指数的时间 依赖性降低所证明的。然而,表达抗间皮素-cd3εtfp的t细胞的靶 细胞杀伤可比抗间皮素car所观察到的更深和更快。例如,在添加 用抗间皮素-cd3εtfp转导的t细胞的4小时内,可基本上完成对表 达间皮素的靶细胞的杀伤。用许多包含其他cd3
的tfp构建体和tcr 构建体转导的t细胞可观察到很少杀伤或无杀伤。用替代的铰链区构 建的抗间皮素tfp可获得类似的结果。与抗间皮素car转导的t细 胞相比,抗间皮素-cd3ε或抗间皮素-cd3γtfp转导的t细胞对间皮 素转导的靶细胞的细胞毒性可能更大。
[0362]
相对于用于转导的病毒量(moi),tfp转导的t细胞的细胞 毒活性可以是剂量依赖性的。随着抗间皮素-cd3εtfp慢病毒的moi 增加,可观察到间皮素阳性细胞的杀伤增加,进一步加强了tfp转导 与细胞毒活性之间的关系。
[0363]
rtca测定的示例性结果显示在图6中。通过将经由编码连接 体:ggggsggggsggggsle(seq id no:1)的dna序列与cd3ε dna片段连接的抗msln scfv dna片段克隆到p510载体(来自 sbi)中的xbai和ecori位点来工程化抗msln tfp构建体。产生 的抗msln tfp构建体是p510_抗msln_ss1_cd3ε(抗msln ss1 scfv-连接体-人cd3ε链)。
[0364]
从pcmv6_xl4_间皮素(origene)经pcr扩增全长间皮素 (nm_005823),并通过gibson重组反应将其克隆到xbai和ecori 限制酶消化的p527a(pcdh-ef1-mcs-t2a-puro)(sbi)中。
[0365]
用于rtca的靶细胞是间皮素阳性hela细胞(宫颈腺癌, ccl-2
tm
),并且间皮素阴性pc-3细胞(前列腺腺癌, crl-1435
tm
)用作阴性对照。将贴壁靶细胞在含有10%fbs和1%抗 生素-抗霉菌剂(life technologies)的dmem中培养。
[0366]
确定指示细胞毒性的归一化细胞指数。活化的pbmc未经处理 (迹线#1)、未转导(迹线#2),或者用空载体(迹线#3)、抗mslntfp(“抗msln-cd3εtruc”,迹线#4)、具有cd28ζ(迹线#5)或 41bbζ(迹线#6)信号传导结构域的抗msln car(分别为“抗msln
-ꢀ
28ζcar”和“抗msln-41bbζcar”)转导。
[0367]
如图6a所示,与阴性对照相比,抗msln tfp转导的t细胞 有效杀伤靶msln阳性hela细胞。相反,msln阴性pc-3细胞未 被任何构建体有效杀伤(图6b)。
[0368]
使用具有单结构域抗间皮素结合物的内部tfp构建体进行了类 似的实验。图6c示出了使用来自两个不同人供体(顶部和底部)的 t细胞在高靶标密度细胞系(hela-(msln
high
))中对msln阳性 细胞的杀伤。示出了具有抗msln结合物sd1(左)、sd4(中)和 sd6(右)的tfp t细胞的细胞杀伤迹线。活化的pbmc未经转导 (迹线#1),或用cd3εtfp(迹线#2)、cd3γtfp(迹线#3)、tcrβ tfp(迹线#4)或cd28ζcar(迹线#5)转导。在实时细胞分析仪(rtca) 测定中确定指示细胞毒性的归一化细胞指数。如图所示,除了未转导 的t细胞外,所有t细胞都能够杀伤癌细胞。实施例7:具有高或低靶标密度的细胞中基于萤光素酶的细胞毒性测定
[0369]
基于萤光素酶的细胞毒性测定(“luc-cyto”测定)通过间接测量 共培养后残留的活靶细胞中的萤光素酶的酶活性来评估tfp t和 car t细胞的细胞毒性。luc-cyto测定中使用的高靶标密度细胞是 hela-msln
high
细胞,使用的低靶标密度细胞是表达低水平间皮素的 pc3细胞(pc3-msln
low
),每种细胞均稳定转导以表达萤火虫萤光 素酶。编码萤火虫萤光素酶的dna由合成,并插入单启动子慢病毒载体pcdh527a-1(system bioscience) 的多克隆位点。
[0370]
如上所述包装携带萤火虫萤光素酶的慢病毒。然后用携带慢病 毒的萤火虫萤光素酶构建体转导hela-msln
high
或pc3-msln
low
细胞 24小时,然后用嘌呤霉素(5μg/ml)选择。
通过用bright-glo
tm
萤光 素酶测定系统(promega)测量细胞中的萤光素酶的酶活性来确认 hela-luc-msln
high-和pc3-luc-msln
low-萤光素酶细胞的产生。
[0371]
用空表达载体(“nt”)或以下tfp或car转导来自两个不同 人供体的单独群体:抗msln(阳性对照,“ss1”,亲和力11nm)、 抗msln-sd1(亲和力25nm)、抗msln-sd4(亲和力6nm)或抗 msln sd6(亲和力0.59nm),其各自为cd3εtfp、cd3γtfp、 tcrβtfp和cd28ζcar的形式。将两个转导的t细胞群体与hela
-ꢀ
msln
high
(图7)或pc3-msln
low
(图8)一起温育。
[0372]
将靶细胞以每孔5000个细胞接种在96孔板中。将tfp t、cart或对照细胞以1:1(黑色条)或1:5(灰色条)的效应物-靶标比添加 至靶细胞。然后将细胞混合物在37℃、5%co2下培养24小时,之后 通过萤光素酶测定系统测量活靶细胞中的萤光素酶的酶 活性。将细胞离心成沉淀物并重新悬浮于含有萤光素酶底物的培养基 中。萤光素酶通过细胞溶解释放,因此,更高的萤光素酶活性对应于 更大百分比的细胞死亡。
[0373]
使用表达高水平msln的细胞的结果显示在图7中。显示出在 无t细胞(“仅靶标”)、空载体转导的(“nt”)、抗msln(阳性对 照,“ss1”)或者具有内部抗间皮素结合物sd1(图7a)、sd4(图 7b)和sd6(图7c)的抗间皮素tfp t细胞的样品中的%细胞杀伤, 其各自为cd3εtfp、cd3γtfp、tcrβtfp和cd28ζcar的形式。 在每个图中,黑色条代表t细胞与靶细胞的比率为1:1,灰色条代表 t细胞与靶细胞的比率为1:5。从图中可以看出,所有tfp-t细胞、 car-t细胞和阳性对照ss1均有效杀伤msln。
[0374]
图8是显示出抗msln car t细胞和tfp t细胞针对表达低 水平间皮素的靶细胞系(pc3-mslnlow)的活性的一系列图示。显示 出在无t细胞(“仅靶标”)、空载体转导的(“nt”)、抗msln(阳 性对照,“ss1”)或者tfp形式为cd3ε(图8a)、cd3γ(图8b)、 tcrβ(图8c)和cd28ζcar(图8d)的抗间皮素结合物sd1、sd4 和sd6的样品中的%细胞杀伤。在每个图中,黑色条代表t细胞与靶 细胞的比率为1:1,灰色条代表t细胞与靶细胞的比率为1:5。对于第 二t细胞供体观察到类似的结果。
[0375]
如图所示,如预期的,1:1比率的t细胞与靶细胞导致靶细胞的 最高水平的杀伤。此外,所有tfp t和car t细胞在表达高水平 msln的细胞中显示出相似的活性。实施例8:通过facs测量t细胞活化
[0376]
使用msln+和msln-k562细胞进行表达抗msln car和 tfp构建体的t细胞的活化,并显示在图9中。如上所述,用50moi lv连续两天转导活化的pbmc并扩充。转导后第8天,在细胞毒性 培养基(不含酚红的rpmi1640(invitrogen)加5%ab血清(geminibioproducts;100-318))中,用靶细胞(过表达msln的k562细胞) 以1:1的e:t比(每种细胞类型0.2x106个)建立pbmc的共培养物。 使用过表达bcma的k562细胞作为阴性对照。在共培养开始后24 小时收获细胞,用pbs洗涤三次,并用live/dead aqua在冰上染色 30分钟。为了阻断fc受体,添加人fc阻断剂(bd)并在室温下温 育10分钟。随后用来自bd biosciences的抗cd3 apc(克隆ucht1)、 抗cd8 apccy7(克隆sk1)、抗cd69-alexa700(克隆fn50) 以及抗cd25-pe(克隆bc96,ebioscience)对细胞进行染色。将细胞 洗涤两次并通过bd lsrii-fortessa进行分析。使用分析软 件(tree star,inc.)如上所述分析数据。
[0377]
如图9a所示,从左到右,t细胞为未转导的、用空载体转导 的、用抗msln-cd3εtfp、抗msln-28ζcar或抗msln-41bbζ car转导的。与msln-细胞共培养的细胞显示在上面一行,
与msln+ 靶细胞共培养的细胞显示在下面一行。然后用对表面活化标志物 cd69和cd25具有特异性的抗体对细胞进行染色。用抗cd69染色 的细胞数对应于x轴,用抗cd25染色的细胞数对应于y轴。如图所 示,相对于与msln-细胞共培养,通过与msln+细胞培养激活了表 达抗间皮素car和tfp构建体的t细胞,如通过升高的cd69和 cd25表达水平所证明的(图9b)。显示了msln-(白色条)和msln+ (黑色条)细胞中每种构建体的cd25+细胞的百分比。
[0378]
使用k562 msln-细胞(图9c,圆形)和k562-msln+细胞(图 9c,正方形)在未转导的t细胞或用抗msln阳性对照结合物(“510
-ꢀ
ss1-cd3ε”)转导的t细胞中进行类似的实验。数据表示cd25+、 cd69+和cd25+/cd69+细胞的总和。在图9d中,显示了与具有tfp 形式cd3ε、cd3γ、tcrβ和cd28ζcar的供体t细胞组合的k562 msln-靶细胞(左图)和k562 msln+细胞(右图)中内部抗msln 结合物sd1(正方形)、sd4(圆形)和sd6(三角形)的数据。使 用来自第二t细胞供体的细胞观察到类似的结果。
[0379]
可通过分析粒酶b产生来类似地评估t细胞的活化。如上所述 培养并扩充t细胞,并根据制造商的试剂盒说明(gemini bioproducts; 100-318)进行粒酶b的细胞内染色。收获细胞,用pbs洗涤三次, 并用人fc阻断剂阻断10min。用抗cd3 apc(克隆ucht1)和抗 cd8 apccy7(克隆sk1)在4℃下针对表面抗原对细胞染色30min。 然后用固定/透化溶液(bd cytofix/cytoperm固定/透化试剂盒,目录 号554714)在4℃下将细胞固定20min,通过用bd perm/wash缓冲 液洗涤使之流动。随后用抗粒酶b alexafluor700(克隆gb11)对细 胞进行染色,用bd perm/wash缓冲液洗涤两次并重新悬浮于facs 缓冲液中。在bd lsrii-fortessa上获得数据并使用(treestar inc.)进行分析。
[0380]
如图10a所示,从左到右,t细胞为未转导的、用空载体转导 的、用抗msln-cd3εtfp、抗msln-28ζcar或抗msln-41bbζ car转导的。与msln-细胞共培养的细胞显示在上面一行,与msln+ 靶细胞共培养的细胞显示在下面一行。用抗grb染色的细胞数对应于 x轴,用抗cd8染色的细胞数对应于y轴。如图所示,表达抗间皮素 car和tfp构建体的t细胞通过与msln+细胞一起培养而活化,而 与msln-细胞一起培养未活化。这些结果再次显示在图10b中,其 中显示了间皮素阴性(“msln
-”
,白色条)和间皮素阳性(“msln+, 黑色条”)细胞中每种构建体的grb+细胞百分比。这些数据证明了表 达msln的细胞特异性激活t细胞的能力。实施例9:通过elisa的il-2和ifn-γ分泌
[0381]
与识别携带同源抗原的细胞相关的效应t细胞活化和增殖的另 一种量度是效应细胞因子如白细胞介素-2(il-2)和干扰素-γ(ifn-γ) 的产生。
[0382]
如产品说明书中所述对人il-2(目录号#eh2il2,thermoscientific)和ifn-γ(目录号#khc4012,invitrogen)进行elisa测 定。在一个实例中,将50μl重建的标准品或样品一式两份添加至96 孔板的每个孔,然后添加50μl生物素化抗体试剂。通过轻轻敲击板 几次来混合样品。然后将50μl标准稀释剂添加至不含标准品或样品 的所有孔,并用粘性板盖小心地密封板,之后在室温(20-25℃)下温 育3小时。然后移除板盖,清空板内容物,并用洗涤缓冲液填充每个 孔。该洗涤程序重复共3次,并将板吸印到纸巾或其他吸收材料上。 向每个孔添加100μl制备的链霉抗生物素-hrp溶液,并添加新的板 盖,之后在室温下温育30分钟。再次移除板盖,丢弃板内容物,并向 每个孔中添加100μl tmb底物溶液。使反应在室温下在黑暗中进行 30分钟,之后向每个孔中添加100μl停止溶液。评估该板。在停止 反应30分钟内,在设定为450nm和550nm的elisa读板仪上测量 吸光度。用450nm值减去550nm
值,并且相对于从il-2标准曲线 获得的值来计算未知样品中的il-2量。
[0383]
或者,使用人细胞因子磁性缓冲试剂盒(invitrogen,lhb0001m) 与人il-2磁珠试剂盒(invitrogen,lhc0021m)和人ifn-γ磁珠试剂 盒(invitrogen,lhc4031m)进行2-plex测定。简言之,将25μl人 il-2和ifn-γ抗体珠添加至96孔板的每个孔中,并按照以下指南洗 涤:用200μl 1x洗涤溶液洗涤两次,使板与磁性96孔板分离器 (invitrogen,a14179)接触,使珠子沉降1分钟并倾析液体。然后, 将50μl温育缓冲液添加至板的每个孔中,并一式两份添加100μl重 建的标准品,或一式三份添加50μl样品(来自细胞毒性测定的上清 液)和50μl测定稀释剂,总体积为150μl。样品在黑暗中以600rpm 用3mm轨道半径的定轨摇床在室温下混合2小时。按照相同的洗涤 指南洗涤板,并向每个孔中添加100μl人il-2和ifn-γ生物素化检 测抗体。样品在黑暗中以600rpm用3mm轨道半径的定轨摇床在室 温下混合1小时。按照相同的洗涤指南洗涤板,并向每个孔中添加100 μl链霉抗生物素-r-藻红蛋白。样品在黑暗中以600rpm用3mm轨 道半径的定轨摇床在室温下混合30分钟。按照相同的洗涤指南将板 洗涤3次,并在倾析液体后,将样品重新悬浮于150μl的1x洗涤溶 液中。样品以600rpm用3mm轨道半径的定轨摇床混合3分钟,并 在4℃下储存过夜。然后,按照相同的洗涤指南洗涤板,并将样品重 新悬浮于150μl的1x洗涤溶液中。
[0384]
使用magpix系统(luminex)和xponent软件读取板。使 用milliplex analyst软件进行数据分析,该软件提供标准曲线和细 胞因子浓度。
[0385]
相对于未转导的或对照car转导的t细胞,用抗间皮素tfp 转导的t细胞在与内源性表达间皮素的细胞或间皮素转导的细胞共 培养时,可产生更高水平的il-2和ifn-γ。相反,与间皮素阴性细胞 或未转导的细胞共培养可导致从tfp转导的t细胞释放很少的或没 有细胞因子。与先前的细胞毒性数据一致,用替代铰链区构建的抗间 皮素tfp在与携带间皮素的靶细胞共培养时可产生类似的结果。
[0386]
与先前的细胞毒性数据一致,抗间皮素-cd3ε和抗间皮素-cd3γ 可产生tfp构建体的最高il-2和ifn-γ水平。然而,用抗间皮素
-ꢀ
cd3ε和抗间皮素-cd3γtfp转导的t细胞产生的细胞因子可与表达 抗间皮素-28ζcar的t细胞产生的细胞因子相当,但tfp表现出更 高水平的靶细胞杀伤。tfp可比car更有效地杀伤靶细胞,但释放 相当的或更低水平的促炎细胞因子,这种可能性代表tfp相对于car 的潜在优势,因为这些细胞因子的升高水平与过继car-t疗法的剂 量限制毒性相关。
[0387]
示例性结果显示在图11中。如上所述,将活化的pbmc用50 moi慢病毒转导连续两天并进行扩充。转导后第8天,在细胞毒性培 养基(不含酚红的rpmi1640(invitrogen)加5%ab血清(geminibioproducts;100-318))中,用靶细胞(过表达msln的k562细胞) 以1:1的e:t比(每种细胞类型0.2x106个)建立pbmc的共培养物。 使用过表达bcma的k562细胞作为阴性对照。24小时后,如上所 述通过elisa分析细胞的ifn-γ(图11a)和il-2(图11b)表达。 在每个图中,从左到右,t细胞为未转导的、用空载体转导的、用抗 msln-cd3εtfp、抗msln-28ζcar或抗msln-41bbζcar转导 的。与msln-细胞共培养的细胞用白色条表示,与msln+靶细胞共 培养的细胞用黑色条表示。如图所示,表达抗间皮素car和tfp构 建体的t细胞通过与msln+细胞共培养而活化,而与msln-细胞共 培养未活化,如通过ifn-γ和il-2产生所证明的,进一步证明了表达 msln的细胞特异性激活t细胞的能力。实施例10:通过流式细胞术的cd107a暴露
[0388]
对于t细胞活化的另一种测定是cd107a的表面表达,cd107a 是溶酶体相关膜蛋白(也称为lamp-1),其位于静息细胞中的细胞 质溶细胞颗粒的膜中。效应t细胞的脱粒(细胞溶解活性的先决条件) 导致在激活诱导的颗粒胞吐后cd107a向细胞表面的动员。因此,除 细胞因子产生外,cd107a暴露还提供了与细胞毒性密切相关的t细 胞活化的额外量度。
[0389]
分别洗涤靶细胞和效应细胞并重新悬浮于细胞毒性培养基 (rpmi+5%人ab血清+1%抗生素抗霉菌剂)中。在0.5μl/孔的 pe/cy7标记的抗人cd107a(lamp-1)抗体(克隆h4a3,bdbiosciences)的存在下,通过将100μl最终体积的2x105个效应细胞 与2x105个靶细胞组合在u形底96孔板(corning)中进行测定。然 后将培养物在37℃、5%co2下温育1小时。在该温育后,立即将10 μl 1:10稀释的分泌抑制剂莫能菌素(1000x溶液,bd golgistop
tm
) 小心地添加至每个孔中,而不干扰细胞。然后将板在37℃、5%co2下再温育2.5小时。在该温育后,用apc抗人cd3抗体(克隆ucht1, bd biosciences)、percp/cy5.5抗人cd8抗体(克隆sk1,bdbiosciences)和pacific blue抗人cd4抗体(克隆rpa-t4,bdbiosciences)对细胞进行染色,然后在37℃、5%co2下温育30分钟。 随后用facs缓冲液洗涤细胞2次,并在分析前重新悬浮于100μlfacs缓冲液和100ul ic固定缓冲液中。
[0390]
通过流式细胞术检测t细胞表面上cd107a的暴露。使用 lsrfortessa
tm
x20(bd biosciences)进行流式细胞术,并使用flowjo 软件(treestar,inc.ashland,or)进行流式细胞术数据的分析。对每 种效应物/靶细胞培养物确定cd3门内为cd107+ve的cd8+效应细 胞的百分比。
[0391]
与先前的细胞毒性和细胞因子数据一致,表达间皮素的靶细胞 与用抗间皮素-28ζcar转导的效应t细胞的共培养可诱导相对于与 间皮素阴性靶细胞一起温育的效应物的表面cd107a表达增加。相比 之下,在相同条件下,表达抗间皮素-cd3εll或抗间皮素-cd3γlltfp的效应物可表现出cd107a表达的5-7倍诱导。用替代的铰链区 构建的抗间皮素tfp在与携带间皮素的靶细胞共培养时可产生类似 的结果。实施例11:体内小鼠功效研究
[0392]
为了评估用抗间皮素tfp转导的效应t细胞在体内实现抗肿瘤 应答的能力,将用抗间皮素-28ζcar、抗间皮素-cd3εll tfp或抗间 皮素-cd3γll tfp转导的效应t细胞过继转移到先前已用间皮素+人 癌细胞系接种的nod/scid/il-2rγ-/-(nsg-jax)小鼠中。
[0393]
在研究开始之前,至少6周龄的雌性nod/scid/il-2rγ-/-(nsg
-ꢀ
jax)小鼠获自jackson laboratory(库存号005557)并在实验使用前 适应3天。用于接种的人间皮素表达细胞系维持在对数期培养中,然 后收获并用台盼蓝计数以确定活细胞计数。在肿瘤攻击当天,将细胞 以300g离心5分钟,并以0.5-1x106个细胞/100μl重新悬浮于预热 的无菌pbs中。制备用于过继转移的t细胞,其未转导或用抗间皮 素-28ζcar、抗间皮素-cd3εll tfp或抗cd3γll tfp构建体转导。 在研究的第0天,用0.5-1x106个间皮素表达细胞静脉内攻击每个实 验组10只动物。3天后,将100μl无菌pbs中的5x106个效应t细 胞群体静脉内转移至每只动物。每天记录动物的详细临床观察直至安 乐死。每周对所有动物进行体重测量直至死亡或安乐死。在过继转移 测试品和对照品后35天对所有动物实施安乐死。研究期间显示垂死 的任何动物由研究主任在与兽医协商后判断是否实施安乐死。
[0394]
相对于未转导的t细胞,用抗间皮素-28ζcar、抗间皮素-cd3ε ll tfp或抗间皮素-cd3γll tfp转导的t细胞的过继转移可延长携 带间皮素表达细胞系肿瘤的小鼠的存活,
并且可表明抗间皮素car 和tfp转导的t细胞都能够介导靶细胞杀伤,并且这些小鼠模型中 的存活相应增加。总体而言,这些数据可表明tfp代表工程化嵌合受 体的替代平台,其在体外和体内均表现出对第一代car的优异抗原 特异性杀伤。实施例12:体内实体瘤异种移植小鼠模型中的人tfp t细胞治疗
[0395]
还可在携带来源于表达人间皮素的all、cll、nhl或msto 人细胞系的皮下实体瘤的免疫受损小鼠模型中测试用人tfp.间皮素 t细胞治疗的功效。响应于用人tfp.间皮素t细胞治疗的肿瘤收缩可 通过卡尺测量肿瘤大小或通过按照由表达gfp的肿瘤细胞发射的绿 色荧光蛋白(gfp)信号的强度来评估。
[0396]
原代人实体瘤细胞可在免疫受损小鼠中生长,而不必在体外培 养。示例性的实体癌细胞包括实体瘤细胞系,诸如在癌症基因组图谱 (tcga)和/或广泛癌症细胞系百科全书(ccle,参见barretina等 人,nature 483:603(2012))中提供的。示例性的实体癌细胞包括从间 皮瘤、肾细胞癌、胃癌、乳腺癌、肺癌、卵巢癌、前列腺癌、结肠癌、 宫颈癌、脑癌、肝癌、胰腺癌、肾癌、子宫内膜癌或胃癌分离的原代 肿瘤细胞。在一些实施方案中,待治疗的癌症选自间皮瘤、乳头状浆 液性卵巢腺癌、透明细胞卵巢癌、混合型米勒氏卵巢癌、子宫内膜粘 液性卵巢癌、胰腺腺癌、胰腺导管腺癌、子宫浆液性癌、肺腺癌、肝 外胆管癌、胃腺癌、食管腺癌、结直肠腺癌和乳腺腺癌。这些小鼠可 用于测试tfp.间皮素t细胞在人肿瘤异种移植模型中的功效(参见 例如,morton等人,nat.procol.2:247(2007))。在皮下植入或注射 1x10
6-1x107个原代细胞(ec基质物质中胶原酶处理的肿瘤悬浮液) 或肿瘤片段(ec基质物质中的原代肿瘤片段)后,让肿瘤生长至200
-ꢀ
500mm3,然后开始治疗。
[0397]
进行一个这样的实验以测试如上所述的间皮瘤异种移植小鼠模 型中msln特异性单结构域抗体(sdab)活性的体内功效。将萤光素 酶标记的mst0-211h-fl-msln-luc以每只小鼠1x106个细胞与 以1:1的比例皮下接种。通过每周两次卡尺测量监测肿瘤体 积。肿瘤注射后14天,当肿瘤体积约为300mm3时,将1x107个t细 胞静脉内注射到每只动物中。所用的t细胞包括用cd3ε-sd1 tfp、 cd3γ-sd1 tfp、cd3ε-sd4 tfp、cd3γ-sd4 tfp、cd28ζsd1 car和 cd28ζsd1 car转导的t细胞。使用没有t细胞注射的一组小鼠作 为阴性对照。
[0398]
结果显示在图12a中。注射cd3ε-sd1 tfp和cd3γ-sd1 tfpt细胞的小鼠显示出最大和最快的肿瘤体积减小,然而注射任何物质 但不注射t细胞的对照小鼠在注射t细胞后显示肿瘤体积减小。
[0399]
通过在间皮瘤异种移植小鼠模型中对存活小鼠进行再攻击,在 体内测试sd1ε-tfp和γ-tfp t细胞的持久性功效。
[0400]
每只小鼠皮下接种1x106个肿瘤细胞(msto 211h fl mslnluc)与(1比1)。一组小鼠注射raji细胞作为阴性对照, 一组小鼠单独注射msto细胞,也作为阴性对照。每周两次通过卡尺 测量监测肿瘤体积。肿瘤注射后14天(当肿瘤体积达到约300mm3时),将1x107个msto(msln+)或raji(msln-,作为阴性对照) 静脉内注射到每只动物中。结果显示在图12b中。图中的每条线代表 一只动物。如图所示,先前已用抗msln tfp t细胞治疗的小鼠能够 再次减小肿瘤体积或根除肿瘤,表明最初注射的t细胞持续存在于小 鼠中,或者小鼠已经产生抗msln记忆应答。相反,用raji(msln-) 细胞再攻击的小鼠无法控制
raji肿瘤的生长,因此说明了tfp t细 胞应答的特异性。实施例13.患者来源的mslnε-tfp t细胞在msln肿瘤异种移植 小鼠模型中的体内功效
[0401]
使用来自卵巢癌患者的sd1ε-tfp t细胞测试sd1ε-tfp t细胞 对表达间皮素的肿瘤细胞(msto-msln-luc)的体外和体内抗肿瘤 功效。
[0402]
如上所述制备慢病毒。从卵巢癌患者的全血制备cd4
+
和cd8
+ t细胞
[0403]
如下从卵巢癌患者的全血纯化cd4
+
和cd8
+ t细胞(示意性概 述显示在图13a中)。收集40-50ml卵巢癌患者的肝素化全血并由 conversant bio(huntsville,alabama)运送过夜。用等体积的pbs稀 释血液,并将35ml稀释的全血小心地铺在50ml锥形管中的15ml (ge healthcare,目录号:17-5442-02)上方。然后在不 减速的情况下在吊桶式转头中在室温下以800
×
g离心20min。抽吸 上层,使界面处的单个核细胞层(淋巴细胞、单核细胞和血小板)不 受干扰。将单个核细胞层转移至新的50ml锥形管中,添加30mlpbs并在室温下以300
×
g离心10min。向沉淀物添加1-2ml ack 溶解缓冲液(thermofisher,目录号:a1049201),充分混合,在室 温下温育2min,并添加20ml pbs,在室温下以300
×
g离心10min。 将细胞沉淀物重新悬浮于10ml冰冷的macs缓冲液中,并通过 cellometer auto 2000对细胞进行计数。根据制造商的说明,使用 miltenyi人cd4/8微珠(目录号:130-045-101;130-045-201)进行 cd4+和cd8+ t细胞分离。
[0404]
如上所述产生tfp t细胞,并通过facs测定转导。在靶细胞 (msln
high
细胞系mst0-211h-fl msln(在内部从亲本msto-211h, atcc,crl-2081产生)上证实间皮素表达,并在与萤光素酶测定同 一天通过流式细胞术证实sd1ε-tfp t细胞的msln-fc表达。在r10 培养基中制备萤光素酶标记的靶细胞(mst0-211h-fl msln-luc或 msln-细胞系c30-luc(a2780,sigma))的单一悬浮液。将100μl 中的1x104个靶细胞添加至96孔平底板。如图所示,以不同的效应物
ꢀ-
靶标比(e:t)以100μl添加tfp t细胞。基于facs的转导效率和t细胞活化测定
[0405]
将tfp t细胞解冻,去珠(如果在dynabeads+il-2条件下离体 扩充),洗涤,然后重新悬浮于不含细胞因子的t细胞培养基中。添 加所需数目的t细胞(100μl)以分别达到5:1、1:1和1:5的效应物: 靶标比。以测试的比率对每种类型的t细胞制备三个重复。然后将细 胞在37℃、5%co2下培养24小时。共培养24小时后,将板以300
ꢀ×
g离心2分钟以沉淀细胞。从每个孔中小心移取100μl的培养上清 液以供luminex测定。将来自bright-glo
tm
萤光素酶测定系统 (promega,#e2650)的100μl测定缓冲液添加至每个孔中。通过轻 轻地上下吸移将每个孔中的内容物混合。将细胞-试剂混合物在室温 下在黑暗中保持3分钟以完全溶解细胞。将来自每个孔的200μl细 胞溶解物转移至greiner-one白色壁96孔板。通过spectramax m5读 板仪(molecular devices)测量发光的相对发光单位(rlu)。通过以下列出的公式计算肿瘤溶解的百分比(%):
测定
[0406]
如前所述,收获来自肿瘤-t细胞共培养物的上清液并储存在
-ꢀ
80℃下。根据制造商的说明,使用millipore luminex试剂盒 (hcd8mag-15k)检测细胞因子谱。在不进行任何稀释的情况下接 种上清液,并使用magpix技术测量读取。皮下间皮瘤异种移植小鼠模型和体内评估
[0407]
在本研究中使用雌性6周龄nsg小鼠(nod.cg-prkdc
scid il2rg
tm1wj1
/szj,目录号:005557,jackson laboratories)。在与研究相 同的条件下使动物适应最少3天。将mst0-211h-flmsln-luc细胞 以1x106个细胞/100μl的浓度悬浮于无菌pbs中。然后将pbs细胞 悬浮液与冰冷的以1:1混合,每只小鼠的最终注射体积为 200μl。将所得细胞悬浮液保持在冰上,直至在小鼠的 背侧后胁皮下施用。用caliper测量以肿瘤体积监测肿瘤生长。肿瘤 体积计算如下:肿瘤体积=1/2(长x宽2)
[0408]
在肿瘤细胞注射后10天,根据肿瘤体积(200~300mm3)将动 物随机化并分成10组以接受来自不同患者的sd1ε-tfp t细胞的注 射(每组小鼠的数目根据在注射当天回收的sd1ε-tfp t细胞的数目 而变化)。t细胞注射日被认为是研究的第0天。在无菌pbs中以5x106个细胞/100μl的浓度制备t细胞。然后通过尾静脉将细胞悬浮 液静脉内注射到小鼠内。来自卵巢癌患者的sd1ε-tfp t细胞的离体扩充
[0409]
用编码具有靶向msln的sd1结合物的tfp的cd3ε形式的 慢病毒制备msln特异性sdab tfp t细胞。在第10天通过活细胞计 数确定的扩充倍数在用制备的细胞中与第0天相 比为8.58至28.2倍(17.8+/-3.3),并且在用制 备的细胞中与第0天相比为10至33.6倍(22.9+/-5.0)。在扩充的 第10天,通过对cd4
+
和cd8
+
群体上gfp和msln-fc的存在的表 面染色确定sd1ε-tfp t细胞的转导效率。用dynabeads+il-2制备 的细胞中的转导效率范围为28.6%至52.1%(40.9+/-4.0%),用 transact+il-7/15制备的细胞中的转导效率为5.7%至46.9%(26.8 +/-6.3%);dynabeads+il-2与transact+il-7/15条件之间的扩充 倍数和转导效率没有显著差异。每个细胞的载体拷贝数与转导效率一 致,在dynabeads+il-2或transact+il7/15条件下为每个细胞约 1~2个拷贝数,但患者1的每个细胞具有0.38个载体拷贝数。来自卵巢癌患者的sd1ε-tfp t细胞的体外抗肿瘤活性
[0410]
使用萤光素酶报告基因肿瘤细胞溶解测定来检测来自卵巢癌患 者的sd1ε-tfp t细胞的体外功效。在测定当天,在msto-211h
-ꢀ
flmsln-luc细胞系上确认间皮素表达(图13b);所有sd1ε-tfp t 细胞显示不同水平的肿瘤杀伤。对于患者1、2、4和5观察到强效的 肿瘤细胞溶解(75%~97%)。在mslnε-tfp t细胞与mst0-211h
-ꢀ
flmsln-luc(msln高表达物)以5:1的效应物:靶标比共培养时, 患者3在5:1的效应物:靶标比下显示~35%的肿瘤溶解,5名患者中的 4名(患者1、2、4和5)在1:1的效应物:靶标比下显示平均50%的 肿瘤溶解,5名患者中的2名(患者4和5)即使在1:5的效应物:靶 标比下也显示~50%的肿瘤溶解。所有t细胞均显示肿瘤细胞的快速 杀伤。当与间皮素阴性细胞系c30-luc共培养时,所有
mslnε-tfp
tm t细胞未观察到肿瘤溶解(图13c)。使用人cd8组分析 来自5名患者的mslnε-tfp的细胞因子谱,与未转导的t细胞相比, mslnε-tfp
tm
t细胞的细胞溶解细胞因子如ifn-γ、gm-csf、粒酶
-ꢀ
a/b、il-2、mip-1α/β、tnf-α和穿孔蛋白显著增加(图13d-图13l)。mslnε-tfp t细胞在表达msln的肿瘤小鼠异种移植模型中的体内功效
[0411]
使用mst0-211h-flmsln-luc建立皮下异种移植的间皮素表 达肿瘤小鼠模型。每周两次测量肿瘤体积。在肿瘤注射后第10天, 平均肿瘤体积达到200-300mm3,并且来自一个正常供体(nd12,图 14a)和患者1-4(图14b-图14e)的第10天扩充的mslnε-tfp t细 胞进行解冻并确认转导效率。静脉内注射每只小鼠5x106个mslnε
-ꢀ
tfp t细胞或匹配的未转导的t细胞,之后监测肿瘤体积。来自4名 患者中的3名(患者1、2和4)的mslnε-tfp t细胞在t细胞注射 后第20天显示完全肿瘤清除。肿瘤清除维持至第40天。6只小鼠中 的5只接受来自患者3的mslnε-tfp t细胞,其显示部分保护。在 接受来自nd12的mslnε-tfp t细胞的所有四名患者中,一名显示 完全肿瘤清除,两名显示部分肿瘤清除。尾注虽然本文已经显示和描述了本发明的优选实施方案,但对于本领 域技术人员而言显而易见的是,这些实施方案仅以示例的方式提供。 在不脱离本发明的情况下,本领域技术人员现将想到许多变化、改变 和替换。应当理解,在实施本发明的过程中可采用本文描述的本发明 实施方案的各种替代方案。以下权利要求旨在限定本发明的范围,并 且由此涵盖这些权利要求范围内的方法和结构及其等同物。附录a:序列概述
起点商标作为专业知识产权交易平台,可以帮助大家解决很多问题,如果大家想要了解更多知产交易信息请点击 【在线咨询】或添加微信 【19522093243】 与客服一对一沟通,为大家解决相关问题。
与客服一对一沟通,为大家解决相关问题。
此文章来源于网络,如有侵权,请联系删除
相关标签:

 热门咨询
热门咨询




tips